
Spirulina platensis Biofertilization for Enhancing Growth, Photosynthetic Capacity and Yield of Lupinus luteus

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Method
2.2.1. Spirulina platensis Biomass Preparation
2.2.2. Spirulina Extract Preparation
2.2.3. Experimental Design
2.2.4. Measurement of Chlorophyll Content Index and Gas Exchange
2.3. Chemical Analysis
2.4. Statistical Analysis
3. Results
3.1. Growth Parameters and Phytohormonal Levels
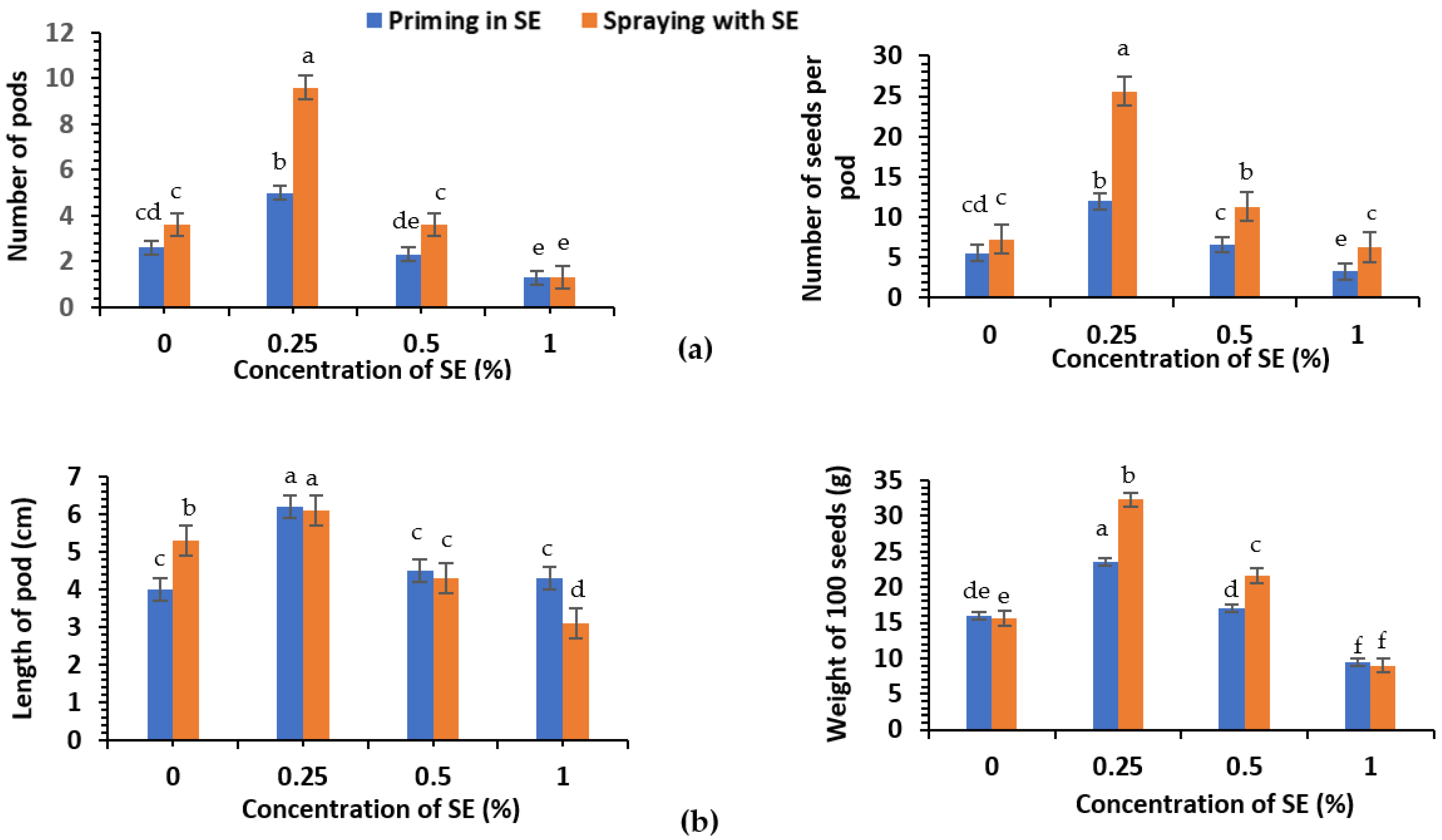
3.2. Yield Parameters
3.3. Photosynthetic Pigments and Activity
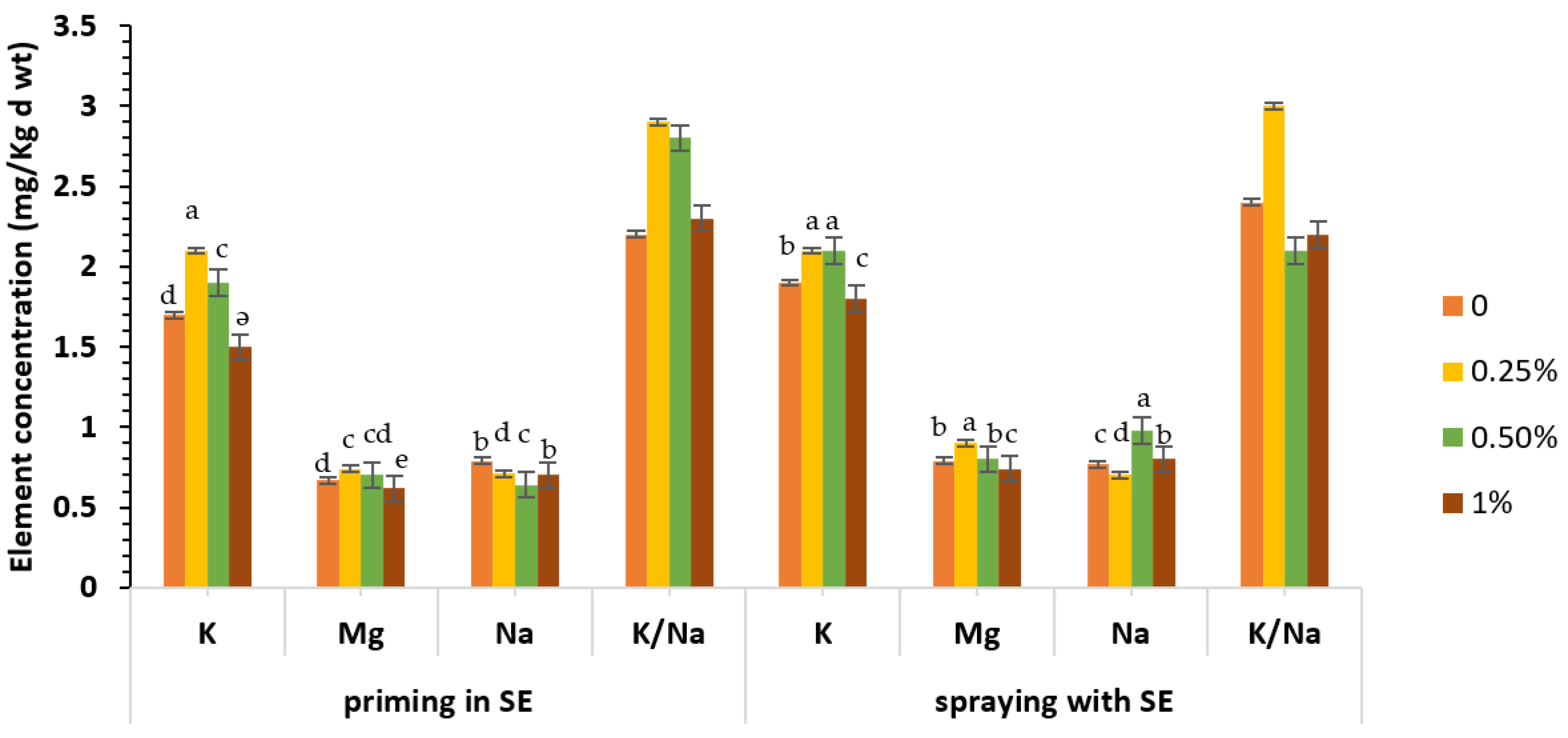
3.4. Total Carbohydrate and Nutrient Content
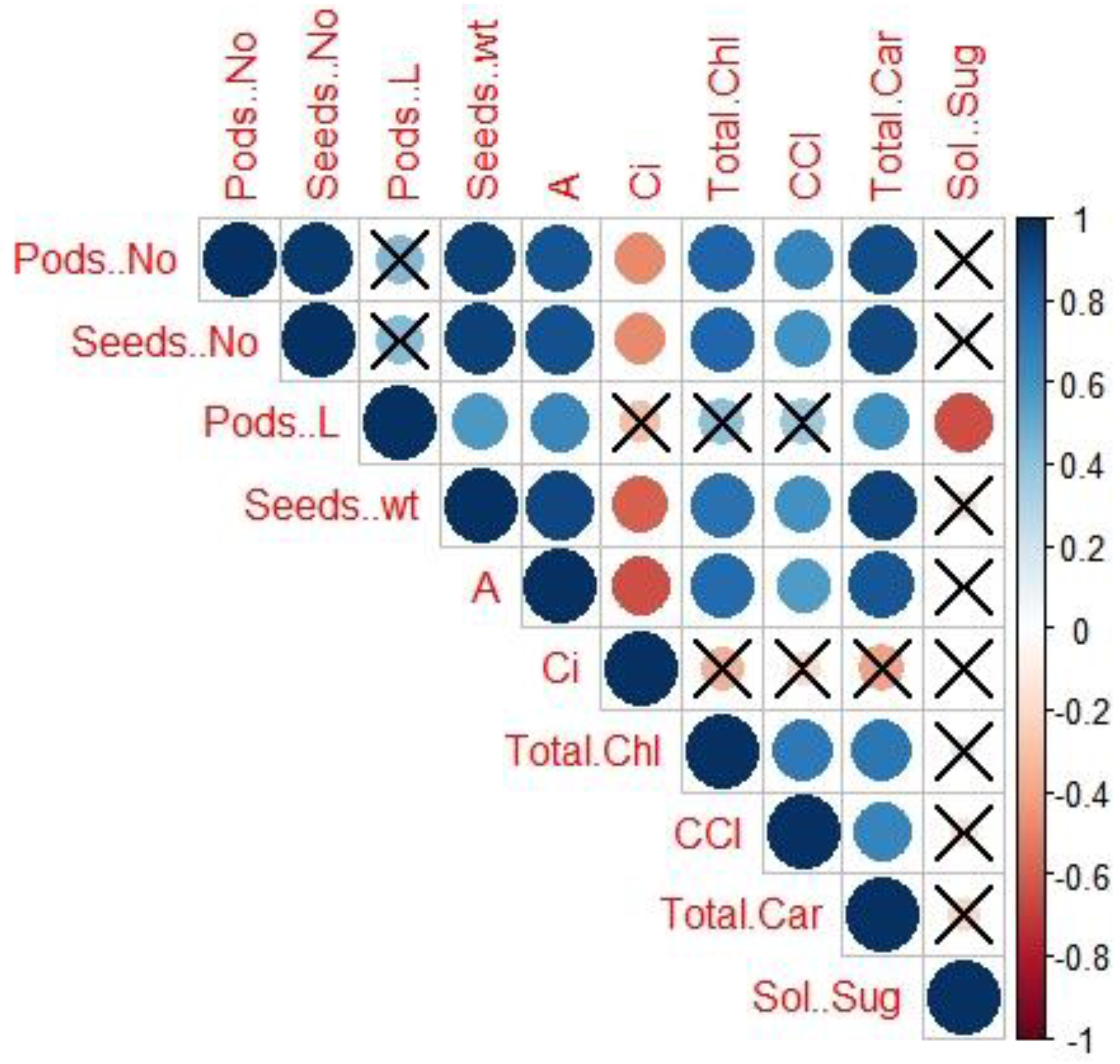
3.5. The Correlation between the Yield Parameters and Photosynthetic Measurements
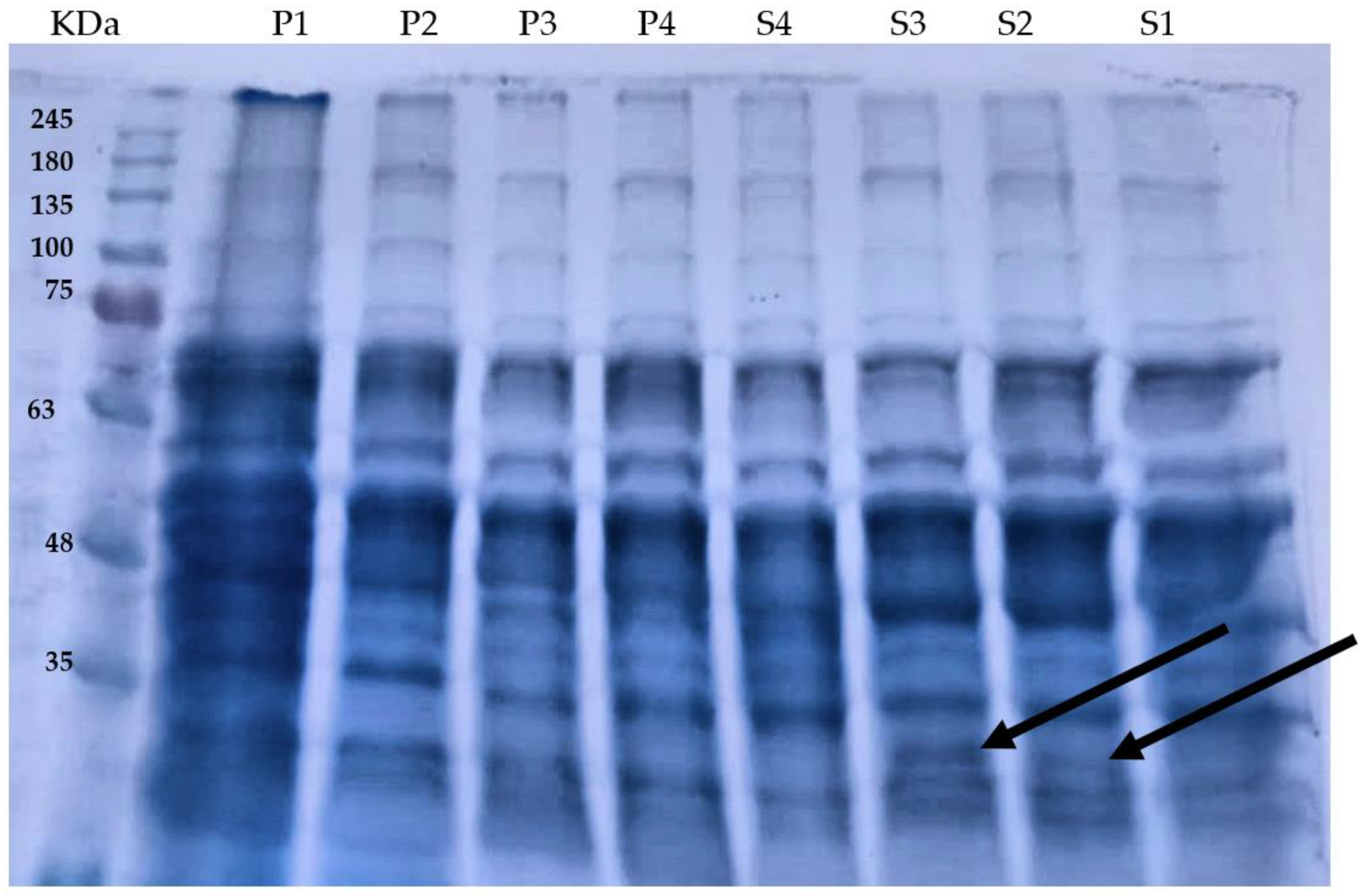
3.6. Protein Profile
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gonçalves, A.L. The use of microalgae and cyanobacteria in the Improvement of agricultural practices: A Review on their biofertilising, biostimulating and biopesticide roles. A review on their biofertilising, biostimulating and biopesticide roles. Appl. Sci. 2021, 11, 871. [Google Scholar] [CrossRef]
- Win, T.T.; Barone, G.D.; Secundo, F.; Fu, P. Algal biofertilizers and plant growth stimulants for sustainable agriculture. Indust. Biotechnol. 2018, 14, 4. [Google Scholar] [CrossRef]
- Kumar, G.; Shekh, A.; Jakhu, S.; Sharma, Y.; Kapoor, R.; Sharma, T.R. Bioengineering of microalgae: Recent advances, perspectives, and regulatory challenges for industrial application. Front. Bioeng. Biotechnol. 2020, 8, 914. [Google Scholar] [CrossRef] [PubMed]
- Singh, S. A review on possible elicitor molecules of cyanobacteria: Their role in improving plant growth and providing tolerance against biotic or abiotic stress. J. Appl. Microbiol. 2014, 117, 1221–1244. [Google Scholar] [CrossRef] [PubMed]
- Santini, G.; Biondi, N.; Rodolfi, L.; Mario, R. Tredici Cyanobacteria: An emerging strategy to improve yields and sustainability in agriculture. Plants 2021, 10, 643. [Google Scholar] [CrossRef]
- Ranjan, K.; PriyaRamakrishnan, P.; Shivay, S. Cyanobacterial inoculation modifies the rhizosphere microbiome of rice planted to a tropical alluvial soil. Appl. Soil Ecol. 2016, 108, 195–203. [Google Scholar] [CrossRef]
- Tuhy, L.; Smoraj, M.; Witkowska, Z.; Chojnacka, K. Biofortification of maize with micronutrients by Spirulina. Open Chem. 2015, 13, 1119–1126. [Google Scholar] [CrossRef]
- Górka, B.; Korzeniowska, K.; Lipok, J.; Wieczorek, P.P. The Biomass of Algae and Algal Extracts in Agricultural Production. In Algae Biomass: Characteristics and Applications; Springer Science and Business Media LLC: Berlin/Heildeberg, Germany, 2018; pp. 103–114. [Google Scholar]
- Renuka, N.; Guldhe, A.; Prasanna, R.; Singh, P.; Bux, F. Microalgae as multi-functional options in modern agriculture: Current trends, prospects and challenges. Biotechnol. Adv. 2018, 36, 1255–1273. [Google Scholar] [CrossRef]
- Anitha, L.; Bramari, G.S.; Kalpana, P. Effect of supplementation of Spirulina platensis to enhance the zinc status in plants of Amaranthus gangeticus, Phaseolus aureus and Tomato. Adv. Biosc. Biotechnol. 2016, 7, 289–299. [Google Scholar] [CrossRef] [Green Version]
- Battacharyya, D.; Babgohari, M.Z.; Rathor, P.; Prithiviraj, B. Seaweed extracts as biostimulants in horticulture. Sci. Hortic. 2015, 196, 39–48. [Google Scholar] [CrossRef]
- Niewiadomska, A.; Sulewska, H.; Wolna-Maruwka, A.; Ratajczak, K.; Waraczewska, Z.; Budka, A. The Influence of Bio-Stimulants and Foliar Fertilizers on Yield, Plant Features, and the Level of Soil Biochemical Activity in White Lupine (Lupinus albus L.) cultivation. Agronomy 2020, 10, 150. [Google Scholar] [CrossRef] [Green Version]
- Panda, D.; Pramanik, K.; Nayak, B.R. Use of seaweed extracts as plant growth regulators for sustainable agriculture. Int. J. Bio-Resour. Stress Manag. 2012, 3, 404–411. [Google Scholar]
- Godlewska, K.; Michalak, I.; Pacyga, P.; Baśladyńska, S.; Chojnacka, K. Potential applications of cyanobacteria: Spirulina platensis filtrates and homogenates in agriculture. World J. Microbiol. Biotechnol. 2019, 35, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koru, E. Earth Food Spirulina (Arthrospira): Production and quality standards. In Food Additive; IntechOpen: London, UK, 2012. [Google Scholar]
- Ahsan, M.; Habib, B.; Parvin, M.; Hasan, R. A Review on Culture, Production and Use of Spirulina as Food for Humans and Feeds for Domestic Animals and Fish; FAO: Rome, Italy, 2008. [Google Scholar]
- Fenner, K.; Canonica, S.; Wackett, L.P.; Elsner, M. Evaluating pesticide degradation in the environment: Blindspots and emerging opportunities. Science 2013, 341, 752–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mala, R.; Ruby, A.S.; Mahalakshmi, R.; Rajeswari, S. Agronomic biofortification of Amaranthus dubius with macro nutrients and vitamin A. In Proceedings of the IOP Conference Series: Materials Science and Engineering, Narsimha Reddy Engineering College, Secunderabad, India, 3–4 July 2017. [Google Scholar]
- Bhowmik, D.; Dubey, J.; Mehra, S. Evaluating potential of Spirulina as inoculant for pulses. Acad. J. Plant Sci. 2010, 3, 161–164. [Google Scholar]
- Geries, L.S.M.; Elsadany, A.Y. Maximizing growth and productivity of onion (Allium cepa L.) by Spirulina platensis extract and nitrogen-fixing endophyte Pseudomonas stutzeri. Arch. Microbiol. 2021, 203, 169–181. [Google Scholar] [CrossRef]
- Hama, J.R.; Bjarne, W.S. Natural alkaloids from narrow-leaf and yellow lupins transfer to soil and soil solution in agricultural fields. Environ. Sci. Eur. 2020, 32, 126. [Google Scholar] [CrossRef]
- Knecht, K.T.; Sanchez, P.; Kinder, D.H. Lupine Seeds (Lupinus spp.): History of Use, Use as An Antihyperglycemic Medicinal, and Use as a Food Plant Nuts and Seeds in Health and Disease Prevention. In Nuts and Seeds in Health and Disease Prevention, 2nd ed.; Academic Press: Cambridge, MA, USA, 2020; pp. 393–402. [Google Scholar]
- Lucas, M.M.; Stoddard, F.L.; Annicchiarico, P.; Frías, J.; Martínez-Villaluenga, C.; Sussmann, D.; Duranti, M.; Seger, A.; Zander, P.M.; Pueyo, J.J. The future of lupin as a protein crop in Europe. Front. Plant Sci. 2015, 6, 705. [Google Scholar] [CrossRef]
- Zarrouk, C. Contribution a L’etude D’une Cyanobacterie: Influence de Divers Facteurs Physiques et Chimiques Sur la Croissance et la Photosynthese de Spirulina Maxima (Setchell et Gardner) Geitler. Ph.D Thesis, University of Paris, Paris, France, 1966. [Google Scholar]
- Pant, G.; Kumar, G.; Karthik, L.; Prasuna, R.G.; Bhaskara, K.V. Antioxidant activity of methanolic extract of blue green algae Anabaena sp. (Nostocaceae). Eur. J. Exp. Biol. 2011, 1, 156–162. [Google Scholar]
- Metzner, H.; Rau, H.; Senger, H. Studies on synchronization of some pigment-deficient Chlorella mutants. Planta 1965, 65, 186. [Google Scholar] [CrossRef]
- Umbriet, W.W.; Burris, R.H.; Stauffer, J.F.; Cohen, P.P.; Johanse, W.J.; Page, L.; Potter, V.R.; Schneided, W.C. Monometric Technique: A Manual Description Method Applicable to Study of Describing Metabolism; Burgess Publishing Company: Minneapolis, MN, USA, 1959; p. 239. [Google Scholar]
- Essa, H.L.; Abdelfattah, M.S.; Marzouk, A.S.; Shedeed, Z.A.; Guirguis, H.A.; El-Sayed, M.M.H. Biogenic copper nanoparticles from Avicennia marina leaves: Impact on seed germination, detoxification enzymes, chlorophyll content and uptake by wheat seedlings. PLoS ONE 2021, 16, e0249764. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680e5. [Google Scholar] [CrossRef] [PubMed]
- Wasfy, W.S.; Orrin, E.S. Identification of plant hormones from Cotton ovules. Plant Physiol. 1975, 55, 550–554. [Google Scholar]
- Ronga, D.; Biazzi, E.; Parati, K.; Carminati, D.; Carminati, E.; Tava, A. Microalgal Biostimulants and Biofertilisers in Crop Productions. Agronomy 2019, 9, 192. [Google Scholar] [CrossRef] [Green Version]
- Supraja, K.V.; Bunushree, B.; Balasubramanian, P. Efficacy of microalgal extracts as biostimulants through seed treatment and foliar spray for tomato cultivation. Ind. Crop. Prod. 2020, 151, 112453. [Google Scholar]
- Wally, O.S.D.; Critchley, A.T.; Hiltz, D.; Craigie, J.S.; Han, X.; Zaharia, L.I.; Abrams, S.R.; Prithiviraj, B. Regulation of phytohormone bio-synthesis and accumulation in Arabidopsis following treatment with commercial extract from the marine macroalga Ascophyllum nodosum. J. Plant Growth Regul. 2013, 32, 324–339. [Google Scholar] [CrossRef]
- Haroun, S.A.; Hussein, M.H. The promotive effect of algal biofertilizers on growth, protein pattern and some metabolic activities of Lupinus termis plants grown in siliceous soil. Asian J. Plant Sci. 2000, 2, 944–951. [Google Scholar] [CrossRef] [Green Version]
- Singh, D.P.; Prabha, R.; Yandigeri, M.S.; Arora, D.K. Cyanobacteria-mediated phenylpropanoids and phytohormones in rice (Oryza sativa) enhance plant growth and stress tolerance. Antonie Van Leeuwenhoek 2011, 100, 557–568. [Google Scholar] [CrossRef]
- Sergeeva, E.; Liaimer, A.; Bergman, B. Evidence for production of the phytohormone indole-3-acetic acid by cyanobacteria. Planta 2002, 215, 229–238. [Google Scholar] [CrossRef]
- Ghaderiardakani, F.; Collas, E.; Damiano, D.K.; Tagg, K.; Graham, N.S.; Coates, J.C. Effects of green seaweed extract on Arabidopsis early development suggest roles for hormone signalling in plant responses to algal fertilizers. Sci. Rep. 2019, 9, 1983. [Google Scholar] [CrossRef]
- Likhitkar, V.S.; Tarar, J.L. Effect of presoaking seed treatment with Nostoc muscorum extracts on cotton. Ann. Plant Physiol. 1995, 9, 113–116. [Google Scholar]
- Adam, M.S. The promotive effect of the cyanobacterium Nostoc muscorum on the growth of some crop plants. Acta Microbiol. Pol. 1999, 48, 163–171. [Google Scholar]
- Dreakeiwicz, M. Chlorophyllase: Occurrence, functions, mechanism of action, effect of external and internal factors. Photosynthetica 1994, 30, 321–332. [Google Scholar]
- Rana, A.; Joshi, M.; Prasanna, R.; Shivay, Y.S.; Nain, L. Biofortification of wheat through inoculation of plant growth promoting rhizobacteria and cyanobacteria. Eur. J. Soil Biol. 2012, 50, 118–126. [Google Scholar] [CrossRef]
- Osman, M.E.H.; Abo-Shady, A.M.; Maysa, M.F.; El-Nagar, M.M.F. The treatment of broad bean seeds with algal suspensions to study their effects on certain growth and yield parameters. J. Environ. Sci. 2020, 49, 1–7. [Google Scholar] [CrossRef]
- Salvi, L.; Brunetti, C.; Cataldo, E.; Storchi, P.; Mattii, G.B. Eco-physiological traits and phenylpropanoid profiling on potted Vitis vinifera L. cv Pinot Noir Subjected to Ascophyllum nodosum treatments under post-version low water availability. Appl. Sci. 2020, 10, 4473. [Google Scholar] [CrossRef]
- Abul Faiz, M.D.; Rakibuzzaman, M.D.; Wasin, E.S.; Asmaul, M.S.T.; Mahato, A.K. Foliar application of Spirulina and Oscillatoria on growth and yield of okra as bio-fertilizer. J. Biosci. Agri. Res. 2019, 22, 1840–1844. [Google Scholar]
- Agwa, O.K.; Ogugbue, C.J.; Williams, E.E. Field evidence of Chlorella vulgaris potentials as a biofertilizer for Hibiscus esculentus. Int. J. Agric. Res. 2017, 12, 181–189. [Google Scholar] [CrossRef] [Green Version]
- Puglisi, I.; Bella, E.; Rovetto, E.I.; Lo Piero, A.R.; Baglieri, A. Biostimulant effect and biochemical response in lettuce seedlings treated with a Scenedesmus quadricauda extract. Plants 2020, 9, 123. [Google Scholar] [CrossRef] [Green Version]
- Spinelli, F.; Fiori, G.; Noferini, M.; Sprocatti, M.; Costa, G. A novel type of seaweed extract as a natural alter-native to the use of iron chelates in strawberry production. Sci. Hortic. 2010, 125, 263–269. [Google Scholar] [CrossRef]
- Fan, D.; Hodges, D.M.; Critchley, A.T.; Prithiviraj, B.A. Commercial extract of brown macroalga (Ascophyllum nodosum) affects yield and the nutritional quality of spinach in vitro. Commun. Soil Sci. Plant Anal. 2013, 44, 1873–1884. [Google Scholar] [CrossRef]
- Villarruel-López, A.; Ascencio, F.; Nuño, K. Microalgae, a potential natural functional food Source—A Review. Pol. J. Food Nutr. Sci. 2017, 67, 251–263. [Google Scholar] [CrossRef] [Green Version]
- Fredeen, A.L.; Raab, T.K.; Rao, I.M.; Terry, N. Effects of phosphorus nutrition on photosynthesis in Glycine max L. Merr. Planta 1990, 181, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Priyadarshani, I.; Rath, B. Commercial and industrial applications of micro algae—A review. J. Algal. Biomass Utln. 2012, 89–100. [Google Scholar]
- Sun, T.; Rao, S.; Zhou, X.; Li, L. Plant carotenoids: Recent advances and future perspective Molecul. Horticulture 2022, 2, 3. [Google Scholar] [CrossRef]
- Li, Y.; He, N.; Hou, J.; Xu, L.; Liu, C.; Zhang, J.; Wang, Q.; Zhang, X.; Wu, X. Factors influencing leaf chlorophyll content in natural forests at the biome scale. Front. Ecol. Evol. 2018, 6, 64. [Google Scholar] [CrossRef] [Green Version]
- Mikiciuk, M.; Dobrolmilska, R. Assessment of yield and physiological indices of small-sized tomato cv. ‘Bianka F1’ under the influence of biostimulators of marine algae origin. Acta Sci. Pol. Hort. Cultus. 2014, 13, 31–41. [Google Scholar]
- Xu, C.; Leskovar, D.I. Effect of A. nodosum extracts on spinach growth, physiology and nutrition value under drought stress. Sci. Hort. 2015, 183, 39–47. [Google Scholar] [CrossRef]
- Wong, S.C.; Cowan, I.R.; Farquhar, G.D. Stomatal conductance correlates with photosynthetic capacity. Nature 1979, 282, 424–426. [Google Scholar] [CrossRef]
- Qu, M.; Hamdani, S.; Li, W.; Wang, S.; Tang, J.; Chen, Z.; Song, Q.; Li, M.; Zhao, H.; Chang, T.; et al. Rapid stomatal response to fluctuating light: An under-explored mechanism to improve drought tolerance in rice. Funct. Plant Biol. 2016, 43, 727–773. [Google Scholar] [CrossRef] [Green Version]
- Mutale-joan, C.; Redouane, B.; Najib, E.; Yassine, K.; Lyamlouli, K.; Laila, S.; Zeroual, Y.; Hicham, E. Screening of microalgae liquid extracts for their bio stimulant properties on plant growth, nutrient uptake and metabolite profile of Solanum lycopersicum L. Sci. Rep. 2020, 10, 2820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakoda, K.; Yamori, W.; Shimada, T.; Sugano, S.S.; Hara-Nishimura, I.; Tanaka, Y. Stomatal density affects gas diffusion and CO2 assimilation dynamics in Arabidopsis under fluctuating light. bioRxiv 2020. [Google Scholar] [CrossRef]
- Pego, J.V.; Kortstee, A.J.; Huijser, C.; Smeekens, S. Photosynthesis, sugars and the regulation of gene expression. J. Exp. Bot. 2000, 51, 407–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araya, T.; Noguchi, K.; Terashima, I. Effects of carbohydrate accumulation on photosynthesis differ between sink and source leaves of Phaseolus vulgaris L. Plant Cell Physiol. 2006, 47, 644–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osman, M.E.H.; El-Sheekh, M.; El-Naggar, A.; Gheda, S. Effect of two species of cyanobacteria as biofertilizers on some metabolic activities, growth, and yield of pea plant. Biol. Fert. Soils 2010, 6, 861–875. [Google Scholar] [CrossRef]
- Popova, L.P.; Stoinova, Z.G.; Maslenkova, L.T. Involvement of abscisic acid in photosynthetic process in Hordeum vulgare L. during salinity stress. J. Plant Growth Regul. 1995, 4, 211–218. [Google Scholar] [CrossRef]
- Duranti, M.; Consonni, A.; Magni, C.; Sessa, F.; Scarafoni, A. The major proteins of lupin seed: Characterization and molecular properties for use as functional and nutraceutical ingredients. Trends Food Sci. Technol. 2008, 19, 624–633. [Google Scholar] [CrossRef]
- Melo, T.S.; Ferreira, R.B.; Teixeira, A.N. The seed storage proteins from Lupinus albus. Phytochemistry 1994, 37, 641–648. [Google Scholar] [CrossRef]
- Dobiesz, W.; Piotrowicz-Cieślak, A.I. Proteins in relation to vigor and viability of white Lupin (Lupinus albus L.) Seed Stored for 26 Years. Front. Plant Sci. 2017, 8, 1392. [Google Scholar] [CrossRef]
- Borowitzka, M.A. Techno-Economic Modeling for Biofuels from Microalgae. In Algae for Biofuels and Energy; Metzler, J.B., Ed.; Springer Netherlands: Dordrecht, The Netherlands, 2013; pp. 255–264. [Google Scholar]
- Tredici, M.R.; Rodolfi, L.; Biondi, N.; Bassi, N.; Sampietro, G. Techno-economic analysis of microalgal biomass production in a 1-ha Green Wall Panel (GWP®) plant. Algal Res. 2016, 19, 253–263. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Root | Shoot | Leaves | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Length (cm) | Fresh wt (g) | Dry wt (mg) | Length (cm) | Fresh wt (g) | Dry wt (mg) | Number | Dry wt (g) | ||
| Priming in 1 SE (%) | 0 | 6.63 ± 0.28 bc | 0.31 ± 0.03 d | 50 ±1 cd | 32.1 ± 2.20 c | 3.50 ± 0.25 b | 446 ± 75 b | 13 ± 0.570 bc | 0.52 ± 0.01 b |
| 0.25 | 8.06 ± 0.72 a | 0.58 ± 0.09 b | 70 ± 5 b | 39.1 ± 1.70 ab | 4.81 ± 0.76 a | 593 ± 85a | 15 ± 1.150 a | 0.70 ± 0.04 a | |
| 0.5 | 7.50 ± 0.25 ab | 0.45 ± 0.02 c | 60 ± 10 bc | 32.2 ± 2.00 c | 3.07 ± 0.25 b | 330 ± 34 bc | 12 ± 1.150 c | 0.47 ± 0.05 bc | |
| 1.0 | 5.30 ± 0.87 d | 0.32 ± 0.01 d | 20 ± 5 e | 24.7± 2.70 d | 2.26 ± 0.21 c | 226 ± 37 c | 11 ± 1.000 c | 0.35 ± 0.04 c | |
| Spraying with 1 SE (%) | 0 | 6.63 ± 0.02 bc | 0.59 ± 0.01 b | 60 ± 5 bc | 37.5 ± 0.76 b | 3.53 ± 0.48 b | 450 ± 34 b | 12 ± 1.000 c | 0.38 ± 0.08 c |
| 0.25 | 8.00 ± 0.23 a | 0.73 ± 0.07 a | 90 ± 5 a | 40.5 ± 1.50 a | 5.19 ± 0.54 a | 620 ± 115 a | 15 ± 1.700 ab | 0.78 ± 0.13 a | |
| 0.5 | 7.23 ± 0.60 ab | 0.56 ± 0.01 b | 50 ± 5 c | 33.3 ± 1.00 c | 3.34 ± 0.22 b | 373 ± 47 b | 13 ± 1.000 abc | 0.43 ± 0.06 bc | |
| 1.0 | 6.20 ± 0.98 c | 0.25 ± 0.08 d | 40 ± 9 d | 31.6 ± 1.00 c | 3.30 ± 0.47 b | 440 ± 50 b | 13 ± 0.057 abc | 0.35 ± 0.04 c | |
| Treatment | Auxin | Gibberellins | Zeatin | Benzyl-Adenine | |
|---|---|---|---|---|---|
| Priming in 1 SE (%) | 0 | 41.61 ± 0.8 e | 119.89 ± 1.70 e | 1.19 ± 0.11 c | 1.05 ± 0.10 e |
| 0.25 | 65.53 ± 0.4 b | 215.55 ± 1.83 b | 2.41 ± 0.16 a | 1.63 ± 0.08 c | |
| 0.5 | 24.52 ± 0.1 f | 149.45 ± 1.09 c | 2.16 ± 0.04 b | 1.44 ± 0.03 d | |
| Spraying with 1 SE (%) | 0 | 48.61 ± 0.9 d | 164.53 ± 2.02 e | 1.21 ± 0.02 c | 1.33 ± 0.04 d |
| 0.25 | 68.38 ± 2.1 a | 941.25 ± 2.26 a | 1.32 ± 0.02 c | 6.33 ± 0.15 a | |
| 0.5 | 52.26 ± 2.8 c | 213.55 ± 2.02 e | 1.30 ± 0.07 c | 2.19 ± 0.10 b | |
| Treatment | Chl a | Chl b | Carotenoids | Total Chl | Chl a/Chl b | CCI | |
|---|---|---|---|---|---|---|---|
| Priming in 1 SE (%) | 0 | 0.682 ± 0.001 cd | 0.339 ± 0.002 e | 0.066 ± 0.001 c | 1.022 ± 0.003 c | 2.0 ± 0.01 b | 19.96 ± 2.9 c |
| 0.25 | 0.720 ± 0.014 b | 0.443 ± 0.024 b | 0.092 ± 0.005 b | 1.163 ± 0.030 b | 1.6 ± 0.08 d | 42.06 ± 4.9 a | |
| 0.5 | 0.620 ± 0.004 e | 0.406 ± 0.010 c | 0.072 ± 0.002 bc | 1.027 ± 0.013 c | 1.4 ± 0.04 de | 32.12 ± 0.49 b | |
| 1.0 | 0.540 ± 0.008 f | 0.371 ± 0.021 d | 0.070 ± 0.003 bc | 0.911 ± 0.025 d | 1.4 ± 0.08 e | 13.65 ± 1.70 d | |
| Spraying with 1 SE (%) | 0 | 0.684 ± 0.001 cd | 0.329 ± 0.010 e | 0.079 ± 0.003 bc | 1.014 ± 0.011 c | 2.0 ± 0.08 b | 22.18 ± 2.2 c |
| 0.25 | 0.867 ± 0.009 a | 0.469 ± 0.017 a | 0.137 ± 0.028 a | 1.334 ± 0.021 a | 1.8 ± 0.07 c | 45.30 ± 2.2 a | |
| 0.5 | 0.695 ± 0.001 c | 0.324 ± 0.004 e | 0.078 ± 0.023 b | 1.019 ± 0.005 c | 2.1 ± 0.03 a | 33.02 ± 1.3 b | |
| 1.0 | 0.679 ± 0.001 d | 0.325 ± 0.003 e | 0.059 ± 0.004 c | 1.005 ± 0.004 c | 2.0 ± 0.02 ab | 20.70 ± 2.2 c | |
| Treatment | Photosynthetic Rate (A) | Stomatal Conductance (Gs) | Transpiration Rate (E) | Internal CO2 (Ci) | |
|---|---|---|---|---|---|
| Priming in 1 SE (%) | 0 | 2.29 ± 0.730 d | 0.020 ± 0.008 d | 0.532 ± 0.01 cd | 243.0 ± 71.8 b |
| 0.25 | 3.39 ± 0.250 b | 0.040 ± 0.009 bc | 0.990 ± 0.01 b | 268.2 ± 35.9 ab | |
| 0.5 | 2.79 ± 0.150 c | 0.027 ± 0.009 cd | 0.820 ± 0.03 bc | 240.2 ± 16.3 b | |
| 1.0 | 1.07 ± 0.380 e | 0.020 ± 0.011 d | 0.480 ± 0.01 d | 312.5 ± 59.0 a | |
| Spraying with 1 SE (%) | 0 | 2.19 ± 0.090 d | 0.030 ± 0.008 cd | 0.960 ± 0.02 b | 293.2 ± 23.4 ab |
| 0.25 | 4.52 ± 0.104 a | 0.050 ± 0.001 a | 1.420 ± 0.03 a | 242.2 ± 4.1 b | |
| 0.5 | 3.52 ± 0.137 b | 0.047 ± 0.005 ab | 1.480 ± 0.01 a | 280.2 ± 9.7 ab | |
| 1.0 | 1.28 ± 0.061 e | 0.020 ± 0.001 d | 0.710 ± 0.08 bcd | 295.5 ± 3.6 ab | |
| Treatment | Soluble Sugar | Insoluble Carbohydrates | Total Carbohydrates | |
|---|---|---|---|---|
| Priming in 1 SE (%) | 0 | 18.74 ± 0.09 c | 34.66 ± 0.53 c | 53.41 ± 0.44 c |
| 0.25 | 16.37 ± 0.02 f | 40.78 ±1.91 b | 57.15 ± 1.89 b | |
| 0.5 | 19.39 ± 0.25 b | 25.70 ± 1.86 de | 45.09 ± 2.05 de | |
| 1.0 | 19.95 ± 0.30 a | 23.07 ± 0.67 e | 43.03 ± 0.36 e | |
| Spraying with 1 SE (%) | 0 | 19.90 ± 0.33 a | 33.32 ± 0.13 c | 53.22 ± 0.43 c |
| 0.25 | 17.38 ± 0.16 e | 46.51 ± 2.14 a | 63.89 ± 2.17 a | |
| 0.5 | 17.77 ± 0.18 d | 36.42 ± 2.88 c | 54.20 ± 3.00 bc | |
| 1.0 | 18.64 ± 0.08 c | 28.69 ± 3.11 d | 47.33 ± 3.03 d | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shedeed, Z.A.; Gheda, S.; Elsanadily, S.; Alharbi, K.; Osman, M.E.H. Spirulina platensis Biofertilization for Enhancing Growth, Photosynthetic Capacity and Yield of Lupinus luteus. Agriculture 2022, 12, 781. https://doi.org/10.3390/agriculture12060781
Shedeed ZA, Gheda S, Elsanadily S, Alharbi K, Osman MEH. Spirulina platensis Biofertilization for Enhancing Growth, Photosynthetic Capacity and Yield of Lupinus luteus. Agriculture. 2022; 12(6):781. https://doi.org/10.3390/agriculture12060781
Chicago/Turabian StyleShedeed, Zeinab A., Saly Gheda, Sara Elsanadily, Khadiga Alharbi, and Mohamed E. H. Osman. 2022. "Spirulina platensis Biofertilization for Enhancing Growth, Photosynthetic Capacity and Yield of Lupinus luteus" Agriculture 12, no. 6: 781. https://doi.org/10.3390/agriculture12060781
APA StyleShedeed, Z. A., Gheda, S., Elsanadily, S., Alharbi, K., & Osman, M. E. H. (2022). Spirulina platensis Biofertilization for Enhancing Growth, Photosynthetic Capacity and Yield of Lupinus luteus. Agriculture, 12(6), 781. https://doi.org/10.3390/agriculture12060781