
Recovery of Orange Peel Essential Oil from ‘Sai-Namphaung’ Tangerine Fruit Drop Biomass and Its Potential Use as Citrus Fruit Postharvest Diseases Control

 ,
,  , , ,
, , , 

Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site and Survey of Preharvest Losses
2.2. Essential Oil and Chemical Analysis
2.3. Isolation of Fungal Causing Post-Harvest Disease
2.4. Pathogenicity Test
2.5. In Vitro Antifungal Assay of the Essential Oil
2.6. In Vivo Antifungal Assay of the Essential Oil
2.7. Statistical Analysis
3. Results and Discussion
3.1. Pre-Harvest Losses
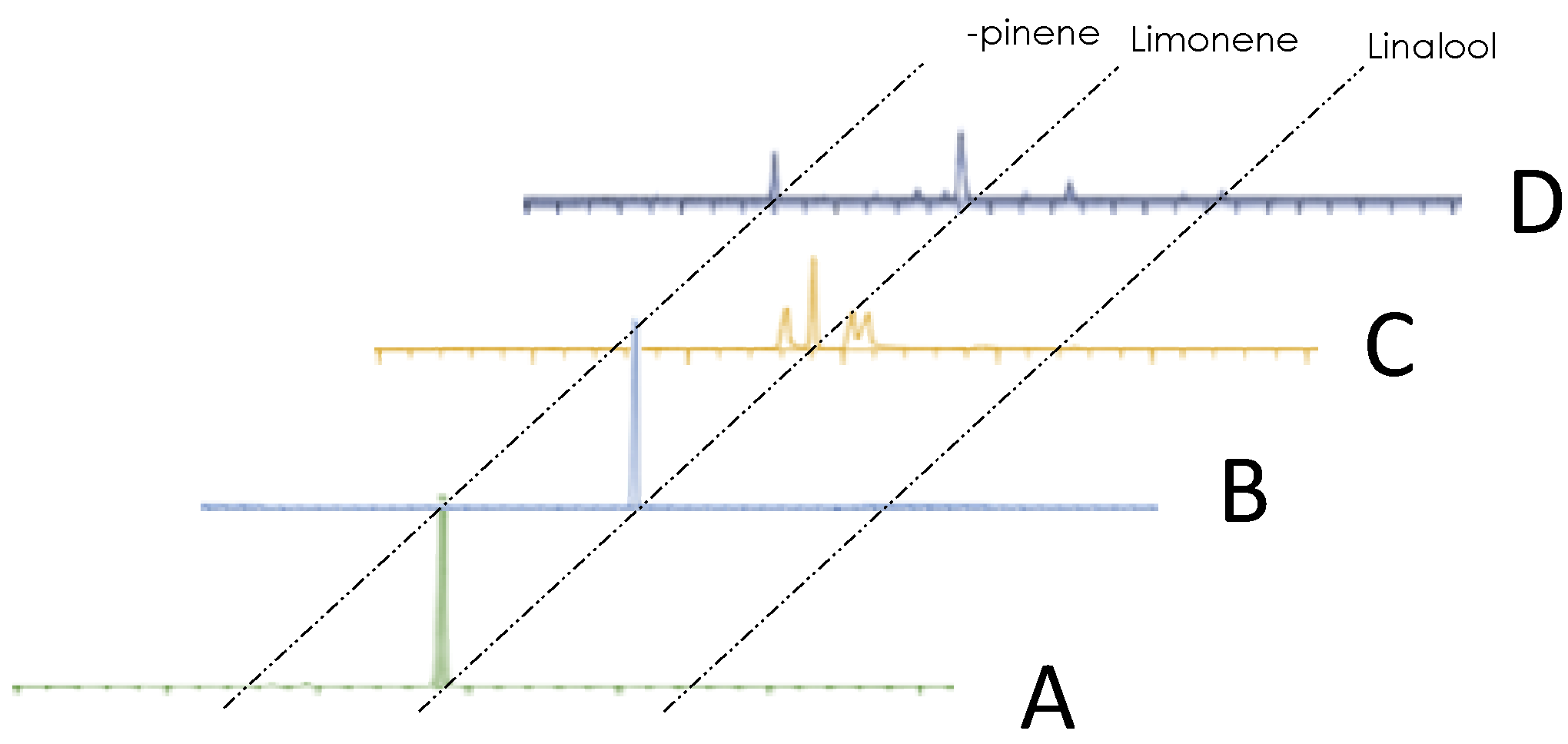
3.2. Volatile Components
3.3. Pathogenic Identification and Phylogenetic Species of Fungi
3.4. In Vitro Antifungal Assays of Essential Oils and In Vivo Study
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Conflicts of Interest
References
- Goldenberg, L.; Yaniv, Y.; Porat, R.; Carmi, N. Mandarin fruit quality: A review. J. Sci. Food Agric. 2018, 98, 18–26. [Google Scholar] [CrossRef]
- Gustafsson, J.; Cederberg, C.; Sonesson, U.; Emanuelsson, A. The Methodology of the FAO Study: Global Food Losses and Food Waste-Extent, Causes and Prevention”-FAO, 2011; Food and Agriculture Organization of the United Nations: Roma, Italy, 2013. [Google Scholar]
- Perondi, D.; Fraisse, C.W.; Dewdney, M.M.; Cerbaro, V.A.; Andreis, J.H.D.; Gama, A.B.; Junior, G.J.S.; Amorim, L.; Pavan, W.; Peres, N.A. Citrus advisory system: A web-based postbloom fruit drop disease alert system. Comput. Electron. Agric. 2020, 178, 105781. [Google Scholar] [CrossRef]
- Boluda-Aguilar, M.; García-Vidal, L.; del Pilar González-Castañeda, F.; López-Gómez, A. Mandarin peel wastes pretreatment with steam explosion for bioethanol production. Bioresour. Technol. 2010, 101, 3506–3513. [Google Scholar] [CrossRef] [PubMed]
- De Barros, C.H.N.; Cruz, G.C.F.; Mayrink, W.; Tasic, L. Bio-based synthesis of silver nanoparticles from orange waste: Effects of distinct biomolecule coatings on size, morphology, and antimicrobial activity. Nanotechnol. Sci. Appl. 2018, 11, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Yi, F.; Jin, R.; Sun, J.; Ma, B.; Bao, X. Evaluation of mechanical-pressed essential oil from Nanfeng mandarin (Citrus reticulata Blanco cv. Kinokuni) as a food preservative based on antimicrobial and antioxidant activities. LWT 2018, 95, 346–353. [Google Scholar] [CrossRef]
- Barbieri, C.; Borsotto, P. Essential oils: Market and legislation. In Potential of Essential Oils; IntechOpen: London, UK, 2018; pp. 107–127. [Google Scholar]
- Grandviewresearch. Citrus Oil Market Size, Share & Trends Analysis Report by Product (Orange Oil, Lemon Oil), by Application (Personal Care, Food & Beverages, Aromatherapy), by Region, and Segment Forecasts, 2019–2025. Available online: https://www.grandviewresearch.com/industry-analysis/citrus-oil-market# (accessed on 26 April 2022).
- Javed, S.; Javaid, A.; Nawaz, S.; Saeed, M.; Mahmood, Z.; Siddiqui, S.; Ahmad, R. Phytochemistry, GC-MS analysis, antioxidant and antimicrobial potential of essential oil from five citrus species. J. Agric. Sci. 2014, 6, 201. [Google Scholar] [CrossRef][Green Version]
- Chandharakool, S.; Koomhin, P.; Sinlapasorn, J.; Suanjan, S.; Phungsai, J.; Suttipromma, N.; Songsamoe, S.; Matan, N.; Sattayakhom, A. Effects of Tangerine Essential Oil on Brain Waves, Moods, and Sleep Onset Latency. Molecules 2020, 25, 4865. [Google Scholar] [CrossRef]
- Tao, N.; Jia, L.; Zhou, H. Anti-fungal activity of Citrus reticulata Blanco essential oil against Penicillium italicum and Penicillium digitatum. Food Chem. 2014, 153, 265–271. [Google Scholar] [CrossRef]
- Hamad, Y.K.; Fahmi, M.M.; Zaitoun, F.M.; Ziyada, S.M. Role of essential oils in controlling fungi that cause decline disease of guava. Int. J. Pure Appl. Biosci. 2015, 3, 143–151. [Google Scholar] [CrossRef]
- Hosseini, S.S.; Khodaiyan, F.; Yarmand, M.S. Optimization of microwave assisted extraction of pectin from sour orange peel and its physicochemical properties. Carbohydr. Polym. 2016, 140, 59–65. [Google Scholar] [CrossRef]
- Meneguzzo, F.; Ciriminna, R.; Zabini, F.; Pagliaro, M. Review of evidence available on hesperidin-rich products as potential tools against COVID-19 and hydrodynamic cavitation-based extraction as a method of increasing their production. Processes 2020, 8, 549. [Google Scholar] [CrossRef]
- Perazzini, H.; Perazzini, M.T.; Freire, F.B.; Freire, F.B.; Freire, J.T. Modeling and cost analysis of drying of citrus residues as biomass in rotary dryer for bioenergy. Renew. Energy 2021, 175, 167–178. [Google Scholar] [CrossRef]
- Bhatti, H.N.; Bajwa, I.I.; Hanif, M.A.; Bukhari, I.H. Removal of lead and cobalt using lignocellulosic fiber derived from Citrus reticulata waste biomass. Korean J. Chem. Eng. 2010, 27, 218–227. [Google Scholar] [CrossRef]
- Porto, D.S.; Forim, M.R.; Costa, E.S.; Fernandes, J.B.; da Silva, M.F. Evaluation of lignins of trunk and roots from Citrus sinensis L. Osbeck: A large available Brazilian biomass. J. Braz. Chem. Soc. 2021, 32, 29–39. [Google Scholar] [CrossRef]
- Bruno, M.R.; Russo, D.; Cetera, P.; Faraone, I.; Lo Giudice, V.; Milella, L.; Todaro, L.; Sinisgalli, C.; Fritsch, C.; Dumarçay, S. Chemical analysis and antioxidant properties of orange-tree (Citrus sinensis L.) biomass extracts obtained via different extraction techniques. Biofuels Bioprod. Biorefining 2020, 14, 509–520. [Google Scholar] [CrossRef]
- Moraes Bazioli, J.; Belinato, J.R.; Costa, J.H.; Akiyama, D.Y.; Pontes, J.G.d.M.; Kupper, K.C.; Augusto, F.; de Carvalho, J.E.; Fill, T.P. Biological control of citrus postharvest phytopathogens. Toxins 2019, 11, 460. [Google Scholar] [CrossRef]
- Talibi, I.; Boubaker, H.; Boudyach, E.; Ait Ben Aoumar, A. Alternative methods for the control of postharvest citrus diseases. J. Appl. Microbiol. 2014, 117, 1–17. [Google Scholar] [CrossRef]
- Jhalegar, M.J.; Sharma, R.; Singh, D. In vitro and in vivo activity of essential oils against major postharvest pathogens of Kinnow (Citrus nobilis × C. deliciosa) mandarin. J. Food Sci. Technol. 2015, 52, 2229–2237. [Google Scholar] [CrossRef]
- Sahoo, U.K.; Nath, A.J.; Lalnunpuii, K. Biomass estimation models, biomass storage and ecosystem carbon stock in sweet orange orchards: Implications for land use management. Acta Ecol. Sin. 2021, 41, 57–63. [Google Scholar] [CrossRef]
- Sangta, J.; Wongkaew, M.; Tangpao, T.; Withee, P.; Haituk, S.; Arjin, C.; Sringarm, K.; Hongsibsong, S.; Sutan, K.; Pusadee, T. Recovery of Polyphenolic Fraction from Arabica Coffee Pulp and Its Antifungal Applications. Plants 2021, 10, 1422. [Google Scholar] [CrossRef]
- Sharma, N.; Tripathi, A. Fungitoxicity of the essential oil of Citrus sinensis on post-harvest pathogens. World J. Microbiol. Biotechnol. 2006, 22, 587–593. [Google Scholar] [CrossRef]
- Tangpao, T.; Krutmuang, P.; Kumpoun, W.; Jantrawut, P.; Pusadee, T.; Cheewangkoon, R.; Sommano, S.R.; Chuttong, B. Encapsulation of Basil Essential Oil by Paste Method and Combined Application with Mechanical Trap for Oriental Fruit Fly Control. Insects 2021, 12, 633. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Yuan-Ying, S.; Cai, L. An optimized protocol of single spore isolation for fungi. Cryptogam. Mycol. 2013, 34, 349–356. [Google Scholar] [CrossRef]
- Haituk, S.; Withee, P.; Sangta, J.; Senwanna, C.; Khamsaw, P.; Karunarathna, A.; Hongsibsong, S.; Sringarm, K.; Prasad, S.K.; Sommano, S.R. Production of Non-Volatile Metabolites from Sooty Molds and Their Bio-Functionalities. Processes 2022, 10, 329. [Google Scholar] [CrossRef]
- Haituk, S.; Suwannarach, N.; Hongsanan, S.; Senwanna, C.; Cheewangkoon, R. New genus of epiphytic sooty mold: Alloscorias syngonii (Readerielliopsidaceae) from Thailand. Phytotaxa 2021, 507, 271–282. [Google Scholar] [CrossRef]
- Billones-Baaijens, R.; Jaspers, M.; Allard, A.; Hong, Y.; Ridgway, H.; Jones, E. Management of Botryosphaeriaceae species infection in grapevine propagation materials. Phytopathol. Mediterr. 2015, 54, 355–367. [Google Scholar]
- Plaza, P.; Torres, R.; Usall, J.; Lamarca, N.; Vinas, I. Evaluation of the potential of commercial post-harvest application of essential oils to control citrus decay. J. Hortic. Sci. Biotechnol. 2004, 79, 935–940. [Google Scholar] [CrossRef]
- Perumal, S.; Pillai, S.; Cai, L.W.; Mahmud, R.; Ramanathan, S. Determination of minimum inhibitory concentration of Euphorbia hirta (L.) extracts by tetrazolium microplate assay. J. Nat. Prod. 2012, 5, 68–76. [Google Scholar]
- Gottwald, J.R.; Krysan, P.J.; Young, J.C.; Evert, R.F.; Sussman, M.R. Genetic evidence for the in planta role of phloem-specific plasma membrane sucrose transporters. Proc. Natl. Acad. Sci. USA 2000, 97, 13979–13984. [Google Scholar] [CrossRef]
- Li, Y.; Zhao, R.; Li, Y.; Zhou, Z. Limonin Enhances the Antifungal Activity of Eugenol Nanoemulsion against Penicillium Italicum In Vitro and In Vivo Tests. Microorganisms 2021, 9, 969. [Google Scholar] [CrossRef]
- Naqvi, S.A.M.H. Diagnosis and Management of Pre and Post-harvest Diseases of Citrus fruit. In Diseases of Fruits and Vegetables Volume I: Diagnosis and Management; Naqvi, S.A.M.H., Ed.; Springer: Dordrecht, The Netherlands, 2004; pp. 339–359. [Google Scholar]
- Timmer, L.; Zitko, S. Evaluation of a model for prediction of postbloom fruit drop of citrus. Plant Dis. 1996, 80, 380–383. [Google Scholar] [CrossRef]
- Lima, W.G.; Spósito, M.B.; Amorim, L.; Gonçalves, F.P.; de Filho, P.A.M. Colletotrichum gloeosporioides, a new causal agent of citrus post-bloom fruit drop. Eur. J. Plant Pathol. 2011, 131, 157. [Google Scholar] [CrossRef]
- Nartvaranant, P. Effects of fruit thinning on fruit drop, leaf carbohydrates concentration, fruit carbohydrates concentration, leaf nutrient concentration and fruit quality in Pummelo cultivar Thong Dee. Songklanakarin J. Sci. Technol. 2016, 38, 249–255. [Google Scholar]
- Garnier, M.; Bové, J.M. Citrus greening disease and the greening bacterium. In Proceedings of the International Organization of Citrus Virologists Conference Proceedings (1957–2010), New Dehli, India, 23–27 November 1992. [Google Scholar]
- Tipu, M.M.H.; Masud, M.M.; Jahan, R.; Baroi, A.; Hoque, A.K.M.A. Identification of citrus greening based on visual symptoms: A grower’s diagnostic toolkit. Heliyon 2021, 7, e08387. [Google Scholar] [CrossRef]
- Katz, E.; Boo, K.H.; Kim, H.Y.; Eigenheer, R.A.; Phinney, B.S.; Shulaev, V.; Negre-Zakharov, F.; Sadka, A.; Blumwald, E. Label-free shotgun proteomics and metabolite analysis reveal a significant metabolic shift during citrus fruit development. J. Exp. Bot. 2011, 62, 5367–5384. [Google Scholar] [CrossRef]
- Spiegel-Roy, P.; Goldschmidt, E.E. The Biology of Citrus; Cambridge University Press: Cambridge, UK, 1996. [Google Scholar]
- Ahmad, M.M.; Rehman, S.U.; Anjum, F.M.; Bajwa, E.E. Comparative physical examination of various citrus peel essential oils. Int. J. Agric. Biol. 2006, 8, 186–190. [Google Scholar]
- Teigiserova, D.A.; Tiruta-Barna, L.; Ahmadi, A.; Hamelin, L.; Thomsen, M. A step closer to circular bioeconomy for citrus peel waste: A review of yields and technologies for sustainable management of essential oils. J. Environ. Manag. 2021, 280, 111832. [Google Scholar] [CrossRef]
- Kamal, G.; Anwar, F.; Hussain, A.; Sarri, N.; Ashraf, M. Yield and chemical composition of Citrus essential oils as affected by drying pretreatment of peels. Int. Food Res. J. 2011, 18, 1275. [Google Scholar]
- Kwangjai, J.; Cheaha, D.; Manor, R.; Sa-ih, N.; Samerphob, N.; Issuriya, A.; Wattanapiromsakul, C.; Kumarnsit, E. Modification of brain waves and sleep parameters by Citrus reticulata Blanco. cv. Sai-Nam-Phueng essential oil. Biomed. J. 2021, 44, 727–738. [Google Scholar] [CrossRef]
- Chutia, M.; Bhuyan, P.D.; Pathak, M.G.; Sarma, T.C.; Boruah, P. Antifungal activity and chemical composition of Citrus reticulata Blanco essential oil against phytopathogens from North East India. LWT-Food Sci. Technol. 2009, 42, 777–780. [Google Scholar] [CrossRef]
- Lota, M.-L.; de Rocca Serra, D.; Tomi, F.; Joseph, C. Chemical variability of peel and leaf essential oils of mandarins from Citrus reticulata Blanco. Biochem. Syst. Ecol. 2000, 28, 61–78. [Google Scholar] [CrossRef]
- Varano, A.; Shirahigue, L.D.; Azevedo, F.A.; Altenhofen da Silva, M.; Ceccato-Antonini, S.R. Mandarin essential oil as an antimicrobial in ethanolic fermentation: Effects on Limosilactobacillus fermentum and Saccharomyces cerevisiae. Lett. Appl. Microbiol. 2022. [Google Scholar] [CrossRef] [PubMed]
- Sriwichai, T.; Suksathan, R.; Charoenlertthanakit, N.; Sommano, S.R. Zanthoxylum spp.: A new potential sources of essential oil for the perfumery and pharmaceutical industries in Thailand. Med. Plants-Int. J. Phytomed. Relat. Ind. 2019, 11, 26–45. [Google Scholar] [CrossRef]
- Sriwichai, T.; Wisetkomolmat, J.; Pusadee, T.; Sringarm, K.; Duangmal, K.; Prasad, S.K.; Chuttong, B.; Sommano, S.R. Aromatic Profile Variation of Essential Oil from Dried Makwhaen Fruit and Related Species. Plants 2021, 10, 803. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.M.; Iqbal, Z.; Anjum, F.M.; Sultan, J.I. Genetic variability to essential oil composition in four citrus fruit species. Pak. J. Bot. 2006, 38, 319. [Google Scholar]
- Li, R.; Yang, J.-J.; Shi, Y.-X.; Zhao, M.; Ji, K.-L.; Zhang, P.; Xu, Y.-K.; Hu, H.-B. Chemical composition, antimicrobial and anti-inflammatory activities of the essential oil from Maqian (Zanthoxylum myriacanthum var. pubescens) in Xishuangbanna, SW China. J. Ethnopharmacol. 2014, 158, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Houbraken, J.; de Vries, R.P.; Samson, R.A. Modern taxonomy of biotechnologically important Aspergillus and Penicillium species. Adv. Appl. Microbiol. 2014, 86, 199–249. [Google Scholar]
- Frisvad, J.C.; Samson, R.A. Polyphasic taxonomy of Penicillium subgenus Penicillium. A guide to identification of food and air-borne terverticillate Penicillia and their mycotoxins. Stud. Mycol. 2004, 49, 1–174. [Google Scholar]
- Torres-Calzada, C.; Tapia-Tussell, R.; Higuera-Ciapara, I.; Perez-Brito, D. Morphological, pathological and genetic diversity of Colletotrichum species responsible for anthracnose in papaya (Carica papaya L). Eur. J. Plant Pathol. 2013, 135, 67–79. [Google Scholar] [CrossRef]
- Ajay Kumar, G. Colletotrichum gloeosporioides: Biology, pathogenicity and management in India. J. Plant Physiol. Pathol. 2014, 2, 2–11. [Google Scholar]
- Nelson, P.E.; Dignani, M.C.; Anaissie, E.J. Taxonomy, biology, and clinical aspects of Fusarium species. Clin. Microbiol. Rev. 1994, 7, 479–504. [Google Scholar] [CrossRef] [PubMed]
- Sandoval-Denis, M.; Guarnaccia, V.; Polizzi, G.; Crous, P. Symptomatic Citrus trees reveal a new pathogenic lineage in Fusarium and two new Neocosmospora species. Pers.-Mol. Phylogeny Evol. Fungi 2018, 40, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Suprapta, D.N.; Arai, K.; Iwai, H. Distribution of Geotrichum candidum citrus race in citrus groves and non-citrus fields in Japan. Mycoscience 1995, 36, 277–282. [Google Scholar] [CrossRef]
- Kara, M.; Soylu, E.M. Assessment of glucosinolate-derived isothiocyanates as potential natural antifungal compounds against citrus sour rot disease agent Geotrichum citri-aurantii. J. Phytopathol. 2020, 168, 279–289. [Google Scholar] [CrossRef]
- Jing, L.; Lei, Z.; Li, L.; Xie, R.; Xi, W.; Guan, Y.; Sumner, L.W.; Zhou, Z. Antifungal activity of citrus essential oils. J. Agric. Food Chem. 2014, 62, 3011–3033. [Google Scholar] [CrossRef]
- Zhang, H.-L.; Gan, X.-Q.; Fan, Q.-F.; Yang, J.-J.; Zhang, P.; Hu, H.-B.; Song, Q.-S. Chemical constituents and anti-inflammatory activities of Maqian (Zanthoxylum myriacanthum var. pubescens) bark extracts. Sci. Rep. 2017, 7, 45805. [Google Scholar]
- Wang, W.; Liu, S.; Deng, L.; Ming, J.; Yao, S.; Zeng, K. Control of Citrus Post-harvest Green Molds, Blue Molds, and Sour Rot by the Cecropin A-Melittin Hybrid Peptide BP21. Front. Microbiol. 2018, 9, 2455. [Google Scholar] [CrossRef]
- Boubaker, H.; Karim, H.; El Hamdaoui, A.; Msanda, F.; Leach, D.; Bombarda, I.; Vanloot, P.; Abbad, A.; Boudyach, E.; Aoumar, A.A.B. Chemical characterization and antifungal activities of four Thymus species essential oils against postharvest fungal pathogens of citrus. Ind. Crops Prod. 2016, 86, 95–101. [Google Scholar] [CrossRef]
- Yang, R.; Miao, J.; Shen, Y.; Cai, N.; Wan, C.; Zou, L.; Chen, C.; Chen, J. Antifungal effect of cinnamaldehyde, eugenol and carvacrol nanoemulsion against Penicillium digitatum and application in postharvest preservation of citrus fruit. LWT 2021, 141, 110924. [Google Scholar] [CrossRef]
- Dou, S.; Ouyang, Q.; You, K.; Qian, J.; Tao, N. An inclusion complex of thymol into β-cyclodextrin and its antifungal activity against Geotrichum citri-aurantii. Postharvest Biol. Technol. 2018, 138, 31–36. [Google Scholar] [CrossRef]
- Wang, W.; de Silva, D.D.; Moslemi, A.; Edwards, J.; Ades, P.K.; Crous, P.W.; Taylor, P.W.J. Colletotrichum Species Causing Anthracnose of Citrus in Australia. J. Fungi 2021, 7, 47. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.K.; Yang, H.J.; Jung, H.; Yoon, D.J.; Sang, M.K.; Jeun, Y.-C. Application of Volatile Antifungal Plant Essential Oils for Controlling Pepper Fruit Anthracnose by Colletotrichum gloeosporioides. Plant Pathol. J. 2015, 31, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Moosa, A.; Farzand, A.; Sahi, S.T.; Khan, S.A.; Abbas, M.F.; Abbas, A.; Binyamin, R. First report of post-harvest Fusarium rot caused by Fusarium oxysporum on Citrus reticulata Blanco cv. ‘Kinnow’in Pakistan. J. Plant Pathol. 2020, 102, 945–946. [Google Scholar] [CrossRef]
- Moosa, A.; Farzand, A.; Khan, S.A.; Ahmad, T.; Aslam, H.M.U.; Saeed, S.; Shafique, M.; Masroor, A.; Akhtar, S. First Report of Postharvest Fusarium Rot of Mandarin (Citrus reticulata ‘Kinnow’) Caused by Fusarium equiseti in Pakistan. Plant Dis. 2021, 105, 4170. [Google Scholar] [CrossRef]
- Hasan, M.F.; Islam, M.A.; Sikdar, B. First report on molecular identification of Fusarium species causing fruit rot of mandarin (Citrus reticulata) in Bangladesh. F1000Research 2020, 9, 1212. [Google Scholar] [CrossRef]
- Singh, H.; Al-samarai, G.; Syarhabil, M. Exploitation of natural products as an alternative strategy to control postharvest fungal rotting of citrus. Int. J. Sci. Res. Publ. 2012, 2, 108–111. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Compound Name | Formular | Retention Index | ‘Sai-Namphaung’ Tangerine | ‘Fremont’ | Zanthoxylum myriacanthum | Commercial | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 162 DAF | 178 DAF | 193 DAF | 207 DAF | 221 DAF | |||||||
| % Yield | 5.27 ± 1.87 d | 10.7± 0.10 a | 5.30 ± 0.30 d | 9.73 ± 0.41 b | 7.70 ± 0.21 c | 3.30 ± 0.93 e | |||||
| 1 | Sabinene | C10H16 | 936 | - | - | - | - | - | - | 0.051 | - |
| 2 | β-pinene | C10H16 | 975 | 0.021 | 0.011 | 0.011 | 0.011 | 0.011 | - | 0.004 | - |
| 3 | β-myrcene | C10H16 | 980 | - | 0.012 | 0.012 | 0.012 | 0.012 | 0.012 | - | - |
| 4 | 1R-α-pinene | C10H16 | 993 | - | - | - | - | - | - | 0.010 | - |
| 5 | α-terpinene | C10H16 | 1009 | - | - | - | - | - | - | 0.012 | - |
| 6 | 2-propanol, 1,1′-oxybis | C6H14O3 | - | - | - | - | - | - | - | - | 0.198 |
| 7 | o-cymene | C10H14 | 1027 | - | - | - | - | - | - | 0.008 | - |
| 8 | limonene | C10H16 | 1032.5 | 1.210 | 0.853 | 0.962 | 0.900 | 0.821 | 1.219 | 0.103 | 0.248 |
| 9 | 1-propanol, 2-(2-hydroxypropoxy) | C6H14O3 | - | - | - | - | - | - | - | - | 0.145 |
| 10 | dipropylene glycol | C6H14O3 | - | - | - | - | - | - | - | - | 0.171 |
| 11 | γ-terpinene | C10H16 | 1046.2 | - | - | - | - | - | - | 0.022 | - |
| 12 | Ocimene | C10H16 | 1089 | - | - | - | - | - | - | 0.005 | - |
| 13 | Linalool | C10H18O | 1104 | - | - | - | - | - | 0.024 | 0.009 | - |
| Isolate Number | Colony | Conidial Morphology | Morphological Characteristics | Pathogenic Score |
|---|---|---|---|---|
| BACI01 | Light green fluffy colony initiated after 14 days | Ovoid, unicellular (~5 × 10 µm) | Penicillium sp. Branching pattern: Regular Phialide: Flask-shaped or cylindrical Conidia shape: Globose smooth-walled conidia, sometimes ellipsoidal, sometimes typical cylindrical and green [53,54] | ++++ |
| BACI02 | White fluffy colony initiated after 7 days | Oblong, unicellular (~3 × 20 µm) | Colletotrichum sp. Branching pattern: Irregular Phialide: Simple, short and erect Conidia shape: Hyaline one celled, cylindrical, with both ends rounded. Sometimes ovoid to oblong, slightly curved or dumbbell shaped depending upon the host from which the pathogen is isolated and its area of origin [55,56] | ++ |
| BACI03 | Light orange fluffy colony initiated after 7 days | Macroconidia: falcate, curved, multicellular, 6 septate (~3 × 70 µm) | Fusarium sp. Branching pattern: Sporodochium Phialide: A monophialide is a condio-phore with only one opening or pore through which endoconidia are extruded, while a polyphialide has two or more such openings orpores. Conidia shape: Three types ofspores-called macroconidia, microconidia, and chlamydospores which are intermediate in size and shape [57,58] | - |
| BACI04 | White fluffy colony initiated after 3 days | Cylindrical unicellular (~1 × 2 µm) | Geotrichum sp. Branching pattern: Either simple (non-branched or mono-verticillate), one-stage branched (biverticillate symmetrical), two-stage branched (biverticillate asymmetrical) or three- to more-staged branched, phialide is flask-shaped, consisting of a cylindrical basal part and a distinct neck, or lanceolate (with a narrow basal part tapering to a somewhat pointed apex). Conidia shape: Oval or cylindrical sometime globose, ellipsoidal, cylindrical or fusiform, hyaline or greenish, smooth or rough-walled [59,60] | - |
| Essential Oil Types | Essential Oil Concentrations | |||||
|---|---|---|---|---|---|---|
| Pathogen Isolates | 256 µL/mL | 128 µL/mL | 64 µL/mL | 32 µL/mL | 16 µL/mL | |
| ‘Sai-Namphaung’ | G | + | + | + | + | + |
| F | - | + | + | + | + | |
| P | - | + | + | + | + | |
| C | - | - | - | + | + | |
| ‘Fremont’ | G | - | - | + | + | + |
| F | - | - | + | + | + | |
| P | - | - | + | + | + | |
| C | - | - | - | - | - | |
| Commercial citrus oil | G | - | - | + | + | + |
| F | - | - | - | - | + | |
| P | - | - | - | - | + | |
| C | - | - | - | - | - | |
| Zanthoxylum myriacanthum oil | G | - | - | - | - | - |
| F | - | - | - | - | - | |
| P | - | - | - | - | - | |
| C | - | - | - | - | - | |
| Citrus Causing Pathogens | Treatments | |||
|---|---|---|---|---|
| Penicillium digitatum | 2 days | 3 days | 4 days | |
| Control | 471.16 ± 211.40 ab | 909.23 ± 70.13 a | 1142.99 ± 12.60 a | |
 |  |  | ||
| ‘Fremont’ | 729.64 ± 153.04 a | 973.17 ± 60.39 a | 1184.73 ± 0.41 a | |
 |  |  | ||
| Commercial citrus oil | 341.40 ± 103.59 c | 617.82 ± 145.13 b | 930.47 ± 98.25 b | |
 |  |  | ||
| Zanthoxylum myriacanthum oil | 296.62 ± 187.68 c | 514.93 ± 214.17 c | 652.59 ± 278.87 c | |
 |  |  | ||
| Colletotrichum gloeosporioides | 3 days | 5 days | 7 days | |
| control | 137.58 ± 27.16 a | 122.19 ± 22.80 a | 128.84 ± 44.05 a | |
 |  |  | ||
| ‘Sai-Namphaung’ | 109.21 ± 9.11 b | 113.66 ± 9.07 b | 97.06 ± 5.25 b | |
 |  |  | ||
| ‘Fremont’ | 72.41 ± 27.26 c | 79.73 ± 23.13 b | 43.39 ± 28.37 d | |
 |  |  | ||
| Commercial citrus oil | 99.28 ± 20.59 b | 80.54 ± 16.75 b | 80.27 ± 16.47 b | |
 |  |  | ||
| Zanthoxylum myriacanthum oil | 74.71 ± 4.46 b | 55.93 ± 2.91 c | 54.47 ± 4.40 d | |
 |  |  | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khamsaw, P.; Lumsangkul, C.; Karunarathna, A.; Onsa, N.E.; Kawichai, S.; Chuttong, B.; Sommano, S.R. Recovery of Orange Peel Essential Oil from ‘Sai-Namphaung’ Tangerine Fruit Drop Biomass and Its Potential Use as Citrus Fruit Postharvest Diseases Control. Agriculture 2022, 12, 701. https://doi.org/10.3390/agriculture12050701
Khamsaw P, Lumsangkul C, Karunarathna A, Onsa NE, Kawichai S, Chuttong B, Sommano SR. Recovery of Orange Peel Essential Oil from ‘Sai-Namphaung’ Tangerine Fruit Drop Biomass and Its Potential Use as Citrus Fruit Postharvest Diseases Control. Agriculture. 2022; 12(5):701. https://doi.org/10.3390/agriculture12050701
Chicago/Turabian StyleKhamsaw, Pattarapol, Chompunut Lumsangkul, Anuruddha Karunarathna, Nuttacha Eva Onsa, Sawaeng Kawichai, Bajaree Chuttong, and Sarana Rose Sommano. 2022. "Recovery of Orange Peel Essential Oil from ‘Sai-Namphaung’ Tangerine Fruit Drop Biomass and Its Potential Use as Citrus Fruit Postharvest Diseases Control" Agriculture 12, no. 5: 701. https://doi.org/10.3390/agriculture12050701
APA StyleKhamsaw, P., Lumsangkul, C., Karunarathna, A., Onsa, N. E., Kawichai, S., Chuttong, B., & Sommano, S. R. (2022). Recovery of Orange Peel Essential Oil from ‘Sai-Namphaung’ Tangerine Fruit Drop Biomass and Its Potential Use as Citrus Fruit Postharvest Diseases Control. Agriculture, 12(5), 701. https://doi.org/10.3390/agriculture12050701