
Grazing Management Targets for Tangolagrass Pastures

 ,
, 
Abstract
1. Introduction
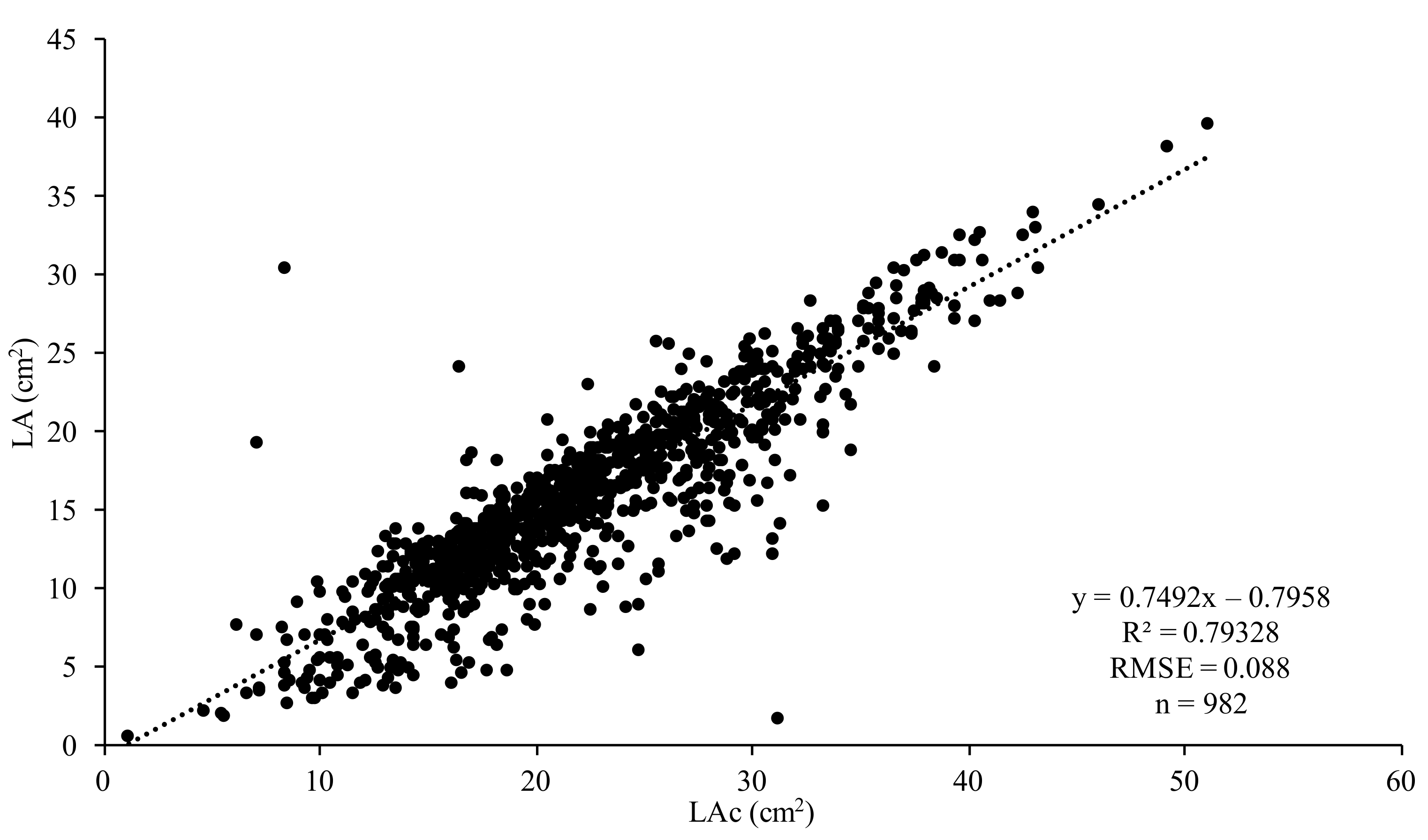
2. Materials and Methods
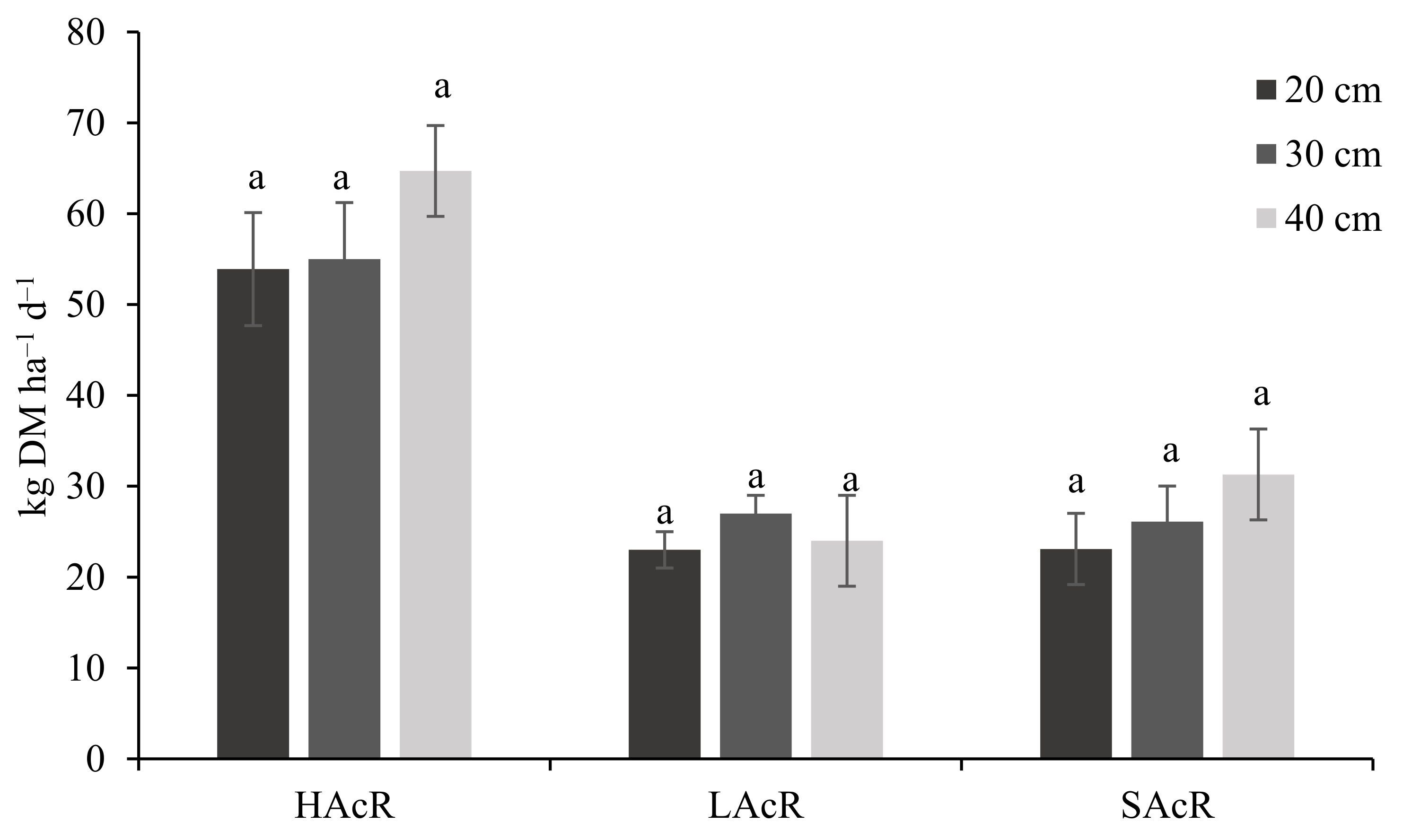
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Andrade, C.M.S.; Assis, G.M.L.; Fazolin, M.; Gonçalves, R.C.; Sales, M.F.L.; Valentin, J.F.; Estrela, J.L.V. (Eds.) Capim-Tangola: Gramínea Forrageira Recomendada Para Solos de Baixa Permeabilidade do Acre; Embrapa Acre: Rio Branco, Brazil, 2009. [Google Scholar]
- Queiroz, D.S.; Casagrande, D.R.; Moura, G.S.; Silva, E.A.; Viana, M.C.M.; Ruas, J.R.M. Espécies forrageiras para produção de leite em solos de várzea. R. Bras. Zootec. 2012, 41, 271–280. [Google Scholar] [CrossRef]
- Bircham, J.S.; Hodgson, J. The influence of sward condition on rates of herbage growth and senescence in mixed swards under continuous stocking management. Grass Forage Sci. 1983, 38, 323–331. [Google Scholar] [CrossRef]
- Parsons, A.J.; Johnson, I.R.; Harvey, A. Use of a model to optimize the interaction between frequency and severity of intermittent defoliation and to provide a fundamental comparison of the continuous and intermittent defoliation of grass. Grass Forage Sci. 1988, 43, 49–59. [Google Scholar] [CrossRef]
- Sbrissia, A.F.; Duchini, P.G.; Zanini, G.D.; Santos, G.T.; Padilha, D.A.; Schmitt, D. Defoliation strategies in pastures submitted to intermittent stocking method: Underlying mechanisms buffering forage accumulation over a range of grazing heights. Crop Sci. 2018, 58, 945–954. [Google Scholar] [CrossRef]
- Sbrissia, A.F.; Schmitt, D.; Duchini, P.G.; Da Silva, S.C. Unravelling the relationship between a seasonal environment and the dynamics of forage growth in grazed swards. J. Agron. Crop Sci. 2020, 206, 630–639. [Google Scholar] [CrossRef]
- Zanella, P.G.; Junior, L.H.P.D.G.; Pinto, C.E.; Baldissera, T.C.; Werner, S.S.; Garagorry, F.C.; Jaurena, M.; Lattanzi, F.A.; Sbrissia, A.F. Grazing intensity drives plant diversity but does not affect forage production in a natural grassland dominated by the tussock-forming grass Andropogon lateralis Nees. Sci. Rep. 2021, 11, 16744. [Google Scholar] [CrossRef]
- Brougham, R.W. Interception of light by the foliage of pure and mixed stands of pasture plants. Aust. J. Agric. Res. 1958, 9, 39–52. [Google Scholar] [CrossRef]
- Carnevalli, R.A.; Da Silva, S.C.; Bueno, A.A.O.; Uebele, M.C.; Bueno, F.O.; Hodgson, J.; Silva, G.; Morais, J.P.G. Herbage production and grazing losses in Panicum maximum cv. Mombaça under four grazing managements. Trop. Grassl. 2006, 40, 165–176. [Google Scholar]
- Zanini, G.D.; Santos, G.T.; Sbrissia, A.F. Frequencies and intensities of defoliation in Aruana guineagrass swards: Morphogenetic and structural characteristics. R. Bras. Zootec. 2012, 41, 1848–1857. [Google Scholar] [CrossRef]
- Migliorini, F.; Soares, A.B.; Schmitt, D.; Pontes, L.S.; Assmann, T. Morphogenetic and structural characteristics of Alexandergrass pastures under continuous stocking. Ciênc. Anim. Bras. 2018, 19, e50830. [Google Scholar] [CrossRef]
- Da Silva, S.C.; Sbrissia, A.F.; Pereira, L.E.T. Ecophysiology of C4 forage grasses—Understanding plant growth for optimising their use and management. Agriculture 2015, 5, 598–625. [Google Scholar] [CrossRef]
- Dong, M.; Pierdominici, M.G. Morphology and growth of stolons and rhizomes in three clonal grasses, as affected by different light supply. Vegetatio 1995, 116, 25–32. [Google Scholar] [CrossRef]
- Wilen, C.A.; Holt, J.S. Spatial Growth of kikuyugrass (Pennisetum clandestinum). Weed Sci. 1996, 44, 323–330. [Google Scholar] [CrossRef]
- Silva, V.J.; Pedreira, C.G.; Sollenberger, L.E.; Carvalho, M.S.; Tonato, F.; Basto, D.C. Seasonal herbage accumulation and nutritive value of irrigated ‘Tifton 85′, Jiggs, and Vaquero bermudagrasses in response to harvest frequency. Crop Sci. 2015, 55, 2886–2894. [Google Scholar] [CrossRef]
- Wallau, M.O.; Sollenberger, L.E.; Vendramini, J.M.; Gomide, C.A.; Mullenix, M.K.; Quesenberry, K.H. Performance of limpograss breeding lines under various grazing management strategies. Crop Sci. 2016, 56, 3345–3353. [Google Scholar] [CrossRef]
- Benvenutti, M.A.; Gordon, I.J.; Poppi, D.P. The effect of the density and physical properties of grass stems on the foraging behaviour and instantaneous intake rate by cattle grazing an artificial reproductive tropical sward. Grass Forage Sci. 2006, 61, 272–281. [Google Scholar] [CrossRef]
- Mezzalira, J.C.; Carvalho, P.C.F.; Fonseca, L.; Bremm, C.; Cangiano, C.; Gonda, H.L.; Laca, E.A. Behavioural mechanisms of intake rate by heifers grazing swards of contrasting structures. Appl. Anim. Behav. Sci. 2014, 153, 1–9. [Google Scholar] [CrossRef]
- Euclides, V.P.B.; Carpejani, G.C.; Montagner, D.B.; Nascimento Júnior, D.; Barbosa, R.A.; Difante, G.S. Maintaining post-grazing sward height of Panicum maximum (cv. Mombaça) at 50 cm led to higher animal performance compared with post-grazing height of 30 cm. Grass Forage Sci. 2018, 73, 174–182. [Google Scholar] [CrossRef]
- Congio, G.F.S.; Batalha, C.D.A.; Chiavegato, M.B.; Berndt, A.; Oliveira, P.P.A.; Frighetto, R.T.S.; Maxwell, T.M.R.; Gregorini, P.; Da Silva, S.C. Strategic grazing management towards sustainable intensification at tropical pasture-based dairy systems. Sci. Total Environ. 2018, 636, 872–880. [Google Scholar] [CrossRef]
- Alvares, C.A.; Stape, J.L.; Sentelhas, P.C.; Gonçalves, J.L.M.; Sparovek, G. Köppen’s climate classification map for Brazil. Meteorol. Z. 2013, 22, 711–728. [Google Scholar] [CrossRef]
- Empresa Brasileira de Pesquisa Agropecuária (EMBRAPA). Sistema Brasileiro de Classificação de Solos; EMBRAPA Solos: Rio de Janeiro, Brazil, 2006. [Google Scholar]
- Comissão de Química e Fertilidade do SoloRS/SC—CQFS. Manual de Adubação e Calagem para os Estados do Rio Grande do Sul e Santa Catarina; Sociedade Brasileira de Fertilidade do Solo: Porto Alegre, Brazil, 2004. [Google Scholar]
- Martins, C.D.M.; Schmitt, D.; Duchini, P.G.; Miqueloto, T.; Sbrissia, A.F. Defoliation intensity and leaf area index recovery in defoliated swards: Implications for forage accumulation. Sci. Agric. 2021, 78, e20190095. [Google Scholar] [CrossRef]
- Schmitt, D.; Padilha, D.A.; Dias, K.M.; Gabriela, T.S.; Rodolfo, G.R.; Zanini, G.D.; Sbrissia, A.F. Chemical composition of two warm-season perennial grasses subjected to proportions of defoliation. Grassl. Sci. 2019, 65, 171–178. [Google Scholar] [CrossRef]
- Ankon Technology. Neutral Detergent Fiber in Feeds—Filter Bag Technique (for A200 and A200I). Method 6. ANKOM Technol. Macedon NY 2014. Available online: https://www.ankom.com/sites/default/files/document-files/Method_6_NDF_A200.pdf (accessed on 5 January 2022).
- Ankon Technology. Acid Detergent Fiber in Feeds—Filter Bag Technique (for A200 and A200I). Method 5. ANKOM Technol. Macedon NY 2014. Available online: https://www.ankom.com/sites/default/files/document-files/Method_5_ADF_A200.pdf (accessed on 5 January 2022).
- Littell, R.C.; Stroup, W.W.; Milliken, G.A.; Wolfinger, R.D.; Schabenberger, O. (Eds.) SAS for Mixed Models; SAS Institute Inc.: Cary, NC, USA, 2006. [Google Scholar]
- Portes Silva, G.; Da Silva, S.C.; Escobar-Guttiérrez, A.; Lemaire, G.; Louarn, G. Stem elongation in Pennisetum purpureum results from a fixed pattern of vegetative development potentially enhanced by the initiation of flowering. Grass Forage Sci. 2019, 74, 708–719. [Google Scholar] [CrossRef]
- Pereira, L.E.T.; Paiva, A.J.; Geremia, E.V.; Silva, S.C. Regrowth patterns of elephant grass (Pennisetum purpureum Schum) subjected to strategies of intermittent stocking management. Grass Forage Sci. 2015, 70, 195–204. [Google Scholar] [CrossRef]
- Rodolfo, G.R.; Schmitt, D.; Dias, K.M.; Sbrissia, A.F. Levels of defoliation and regrowth dynamics in elephant grass swards. Cienc. Rural 2015, 45, 1299–1304. [Google Scholar] [CrossRef][Green Version]
- Matthew, C.; Lemaire, G.; Sackville-Hamilton, N.R.; Hernández Garay, A. A modified self- thinning equation to describe size/density relationships for defoliated swards. Ann. Bot. 1995, 76, 579–587. [Google Scholar] [CrossRef]
- Hernández Garay, A.; Matthew, C.; Hodgson, J. Tiller size/density compensation in perennial ryegrass miniature swards subject to differing defoliation heights and a proposed productivity index. Grass For. Sci. 1999, 54, 347–356. [Google Scholar] [CrossRef]
- Sbrissia, A.F.; Da Silva, S.C. Compensação tamanho/densidade populacional de perfilhos em pastos de capim-marandu. R. Bras. Zootec. 2008, 37, 35–47. [Google Scholar] [CrossRef]
- Martínez Calsina, L.; Agnusdei, M.G.; Assuero, S.G.; Pérez, H. Size⁄density compensation in Chloris gayana Kunth cv. Fine Cut subjected to different defoliation regimes. Grass For. Sci. 2012, 67, 255–262. [Google Scholar] [CrossRef]
- Da Trindade, J.K.; Da Silva, S.C.; Souza Júnior, S.J.; Giacomini, A.A.; Zeferino, C.V.; Guarda, V.D.A.; Carvalho, P.C.F. Composição morfológica da forragem consumida por bovinos de corte durante o rebaixamento do capim-marandu submetido a estratégias de pastejo rotativo. Pesqui. Agropecu. Bras. 2007, 42, 883–890. [Google Scholar] [CrossRef]
- Rouquette, F.M. Invited Review: The roles of forage management, forage quality, and forage allowance in grazing research. Prof. Anim. Sci. 2016, 32, 10–18. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Month | Average Temperature (°C) | Monthly Precipitation (mm) | ||
|---|---|---|---|---|
| 2018 | 2019 | 2018 | 2019 | |
| January | 24.6 | 27.1 | 120.6 | 112.9 |
| February | 24.5 | 24.8 | 30.5 | 105.9 |
| March | 24.7 | 22.8 | 99.9 | 76.9 |
| April | 24.1 | 22.4 | 23.1 | 426.1 |
| May | 20.6 | 12.7 | 111.4 | 357.8 |
| June | 17.1 | 12.6 | 125.4 | 23.7 |
| Block | pH | OM 1 | P | K | H + Al | CEC |
|---|---|---|---|---|---|---|
| % | mg dm−3 | cmolc dm−3 | ||||
| I | 6.4 | 1.4 | 2.0 | 88 | 6.5 | 5.54 |
| II | 6.4 | 1.5 | 1.9 | 55 | 8.1 | 5.62 |
| III | 6.2 | 2.0 | 2.5 | 74 | 11.4 | 6.37 |
| Variable | Height (cm) | SEM | ||
|---|---|---|---|---|
| 20 | 30 | 40 | ||
| TPD 1 | 1190 A | 1290 A | 1130 A | 97.4 |
| LAI pre | 1.9 A | 2.1 A | 2.5 A | 0.22 |
| LAI post | 0.8 A | 0.5 A | 0.6 A | 0.33 |
| Variable | Height (cm) | SEM | ||
|---|---|---|---|---|
| 20 | 30 | 40 | ||
| Pre-grazing | ||||
| HM 1 | 3430 B | 3710 B | 4470 A | 201 |
| Leaves | 25 A | 22 A | 22 A | 1.31 |
| Stems | 45 A | 47 A | 49 A | 1.98 |
| Senesc. tissues | 26 A | 28 A | 27 A | 2.49 |
| Post-grazing | ||||
| HM | 1820 B | 2310 A | 2280 A | 165 |
| Leaves | 9 A | 8 A | 8 A | 1.74 |
| Stems | 48 A | 48 A | 50 A | 1.82 |
| Senesc. tissues | 38 A | 40 A | 39 A | 1.85 |
| Variable | Height (cm) | SEM | ||
|---|---|---|---|---|
| 20 | 30 | 40 | ||
| NDF 1 | 55.0 A | 57.2 A | 57.9 A | 1.97 |
| ADF | 23.7 A | 25.2 A | 26.0 A | 0.95 |
| CP | 22.4 A | 24.7 A | 22.7 B | 0.83 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mocelin, N.G.; Schmitt, D.; Zanini, G.D.; Camacho, P.A.G.; Sbrissia, A.F. Grazing Management Targets for Tangolagrass Pastures. Agriculture 2022, 12, 279. https://doi.org/10.3390/agriculture12020279
Mocelin NG, Schmitt D, Zanini GD, Camacho PAG, Sbrissia AF. Grazing Management Targets for Tangolagrass Pastures. Agriculture. 2022; 12(2):279. https://doi.org/10.3390/agriculture12020279
Chicago/Turabian StyleMocelin, Nemora Guliane, Daniel Schmitt, Guilherme Doneda Zanini, Pedro Antonio Garzón Camacho, and André Fischer Sbrissia. 2022. "Grazing Management Targets for Tangolagrass Pastures" Agriculture 12, no. 2: 279. https://doi.org/10.3390/agriculture12020279
APA StyleMocelin, N. G., Schmitt, D., Zanini, G. D., Camacho, P. A. G., & Sbrissia, A. F. (2022). Grazing Management Targets for Tangolagrass Pastures. Agriculture, 12(2), 279. https://doi.org/10.3390/agriculture12020279