
Emergence of Fungicide Sensitivity in Leptosphaeria maculans Isolates Collected from the Czech Republic to DMI Fungicides


Abstract
1. Introduction
2. Materials and Methods
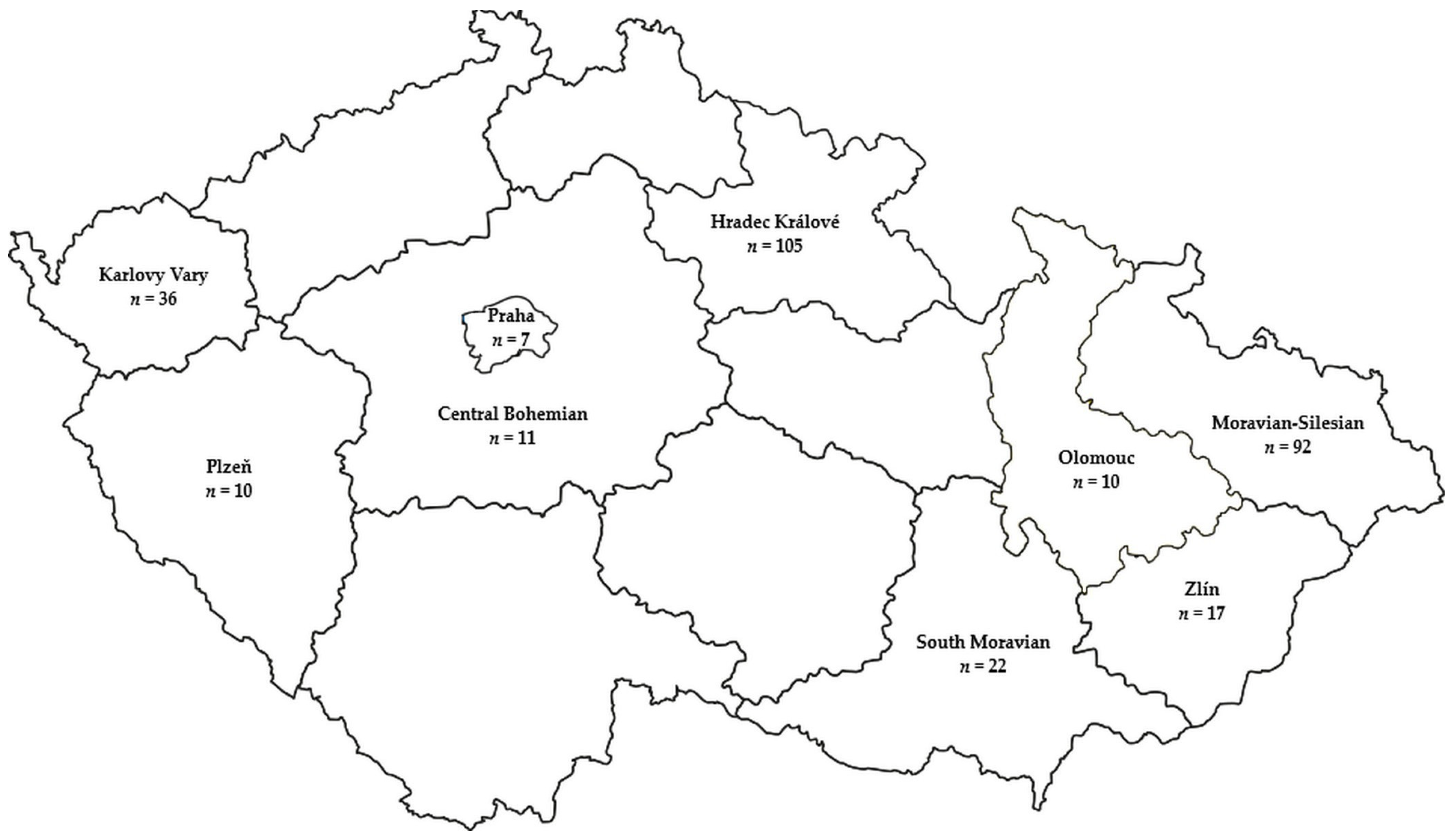
2.1. Sample Collection and Pathogen Isolation
2.2. Confirmation of L. maculans Isolates
2.3. Fungicides
2.4. In Vitro Sensitivity of L. maculans isolates to Tetraconazole, Metconazole, and Prochloraz Using the Mycelium Growth Plate Method
2.5. In Vitro Sensitivity of Conidiospores of L. maculans to Tetraconazole, Metconazole, and Prochloraz Using the Microtiter Plate Method
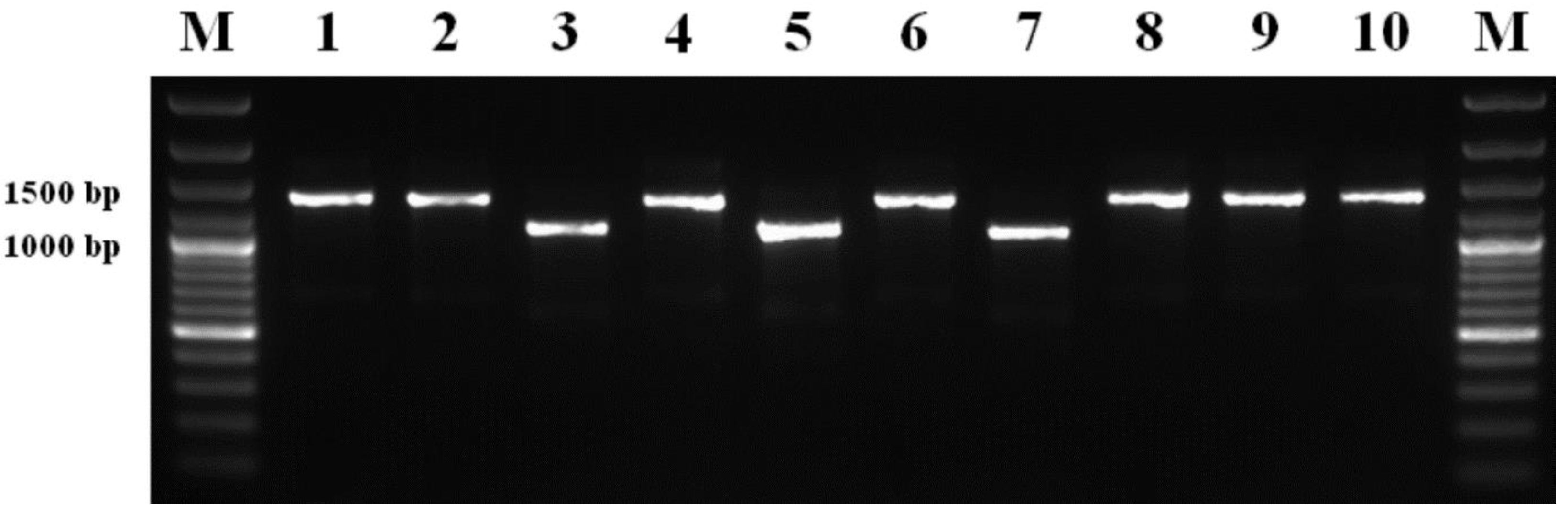
2.6. PCR Amplification of ERG11 Regulatory Region for Molecular Detection of Insertions Conferring Resistance to DMI
2.7. Data Analysis
3. Results
3.1. Mycelium Growth Plate Assay
3.2. Microtiter Plate Assay
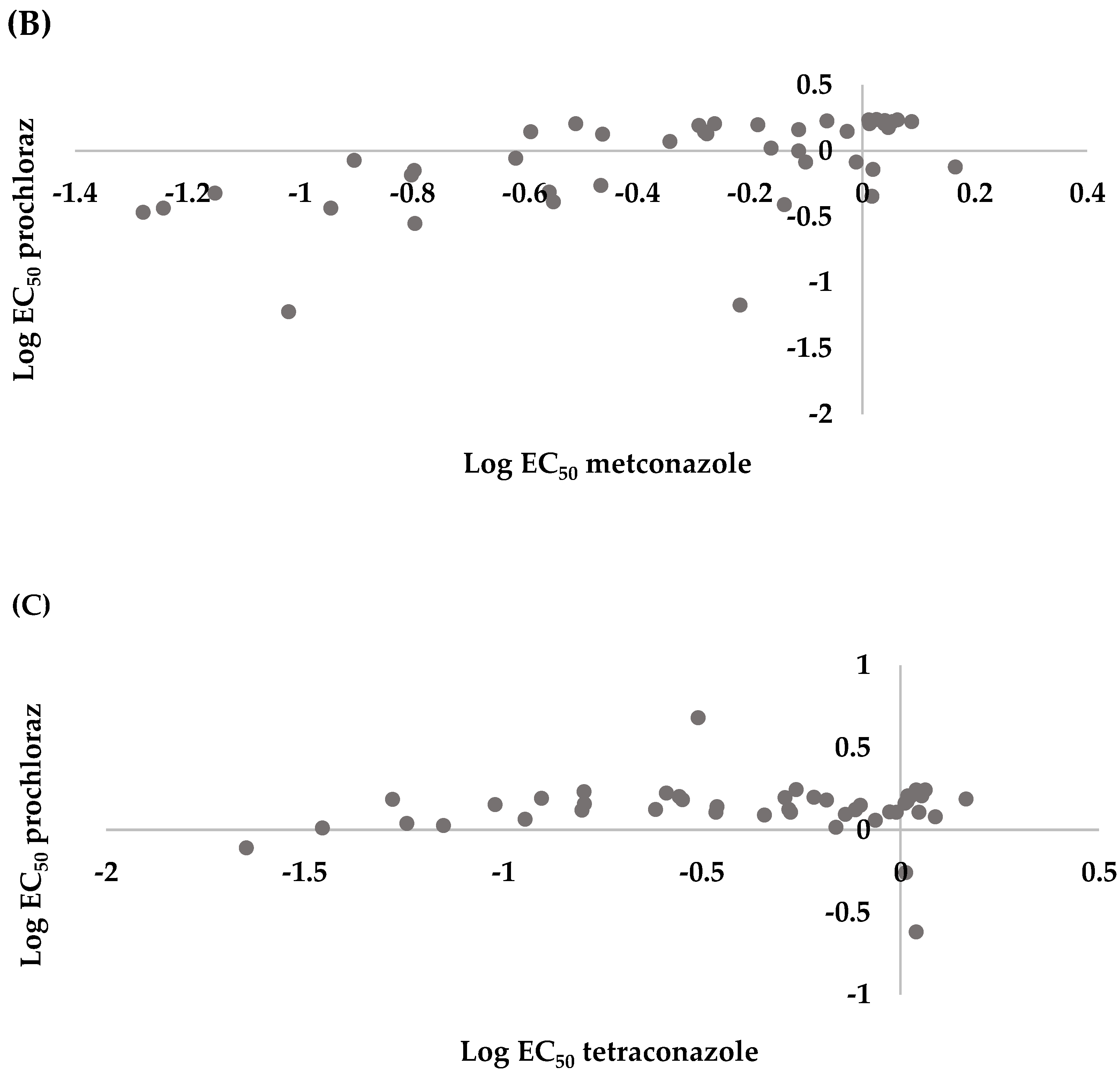
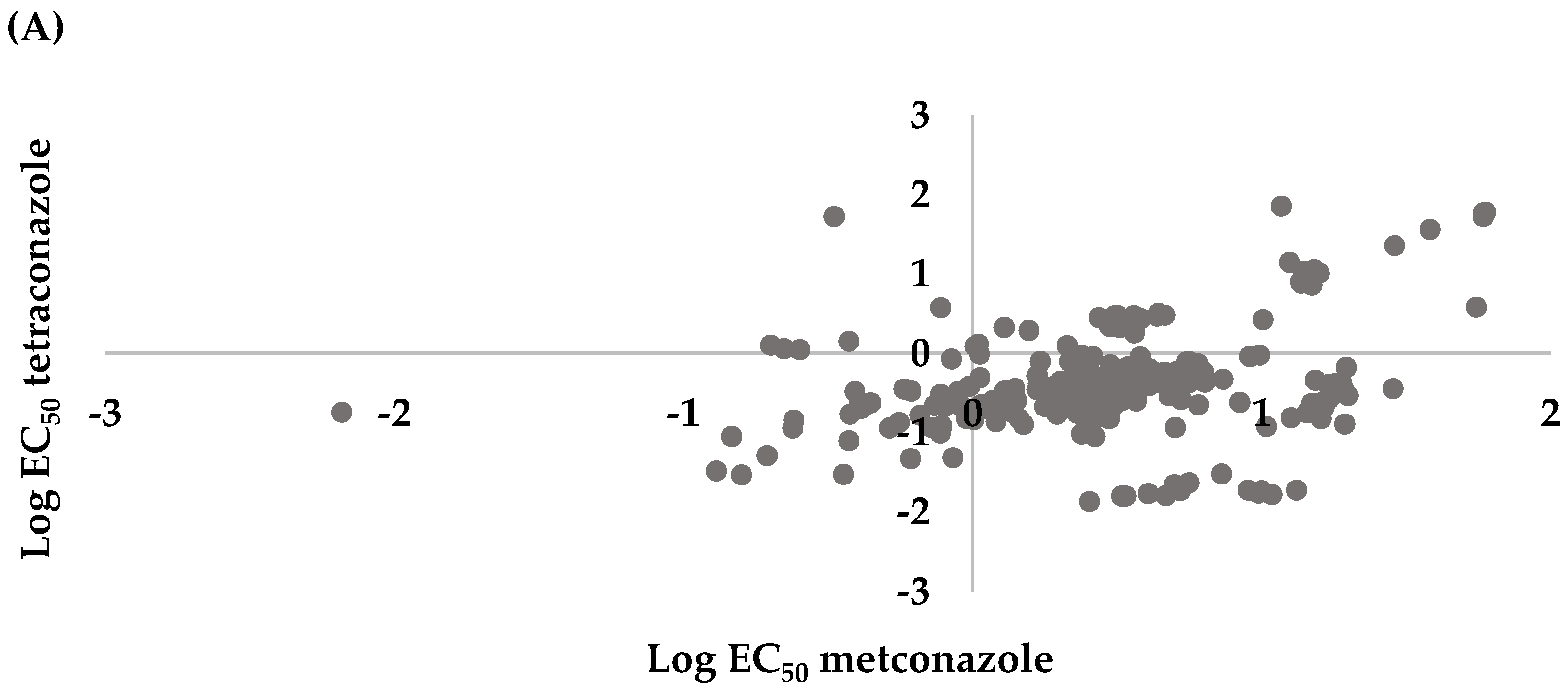
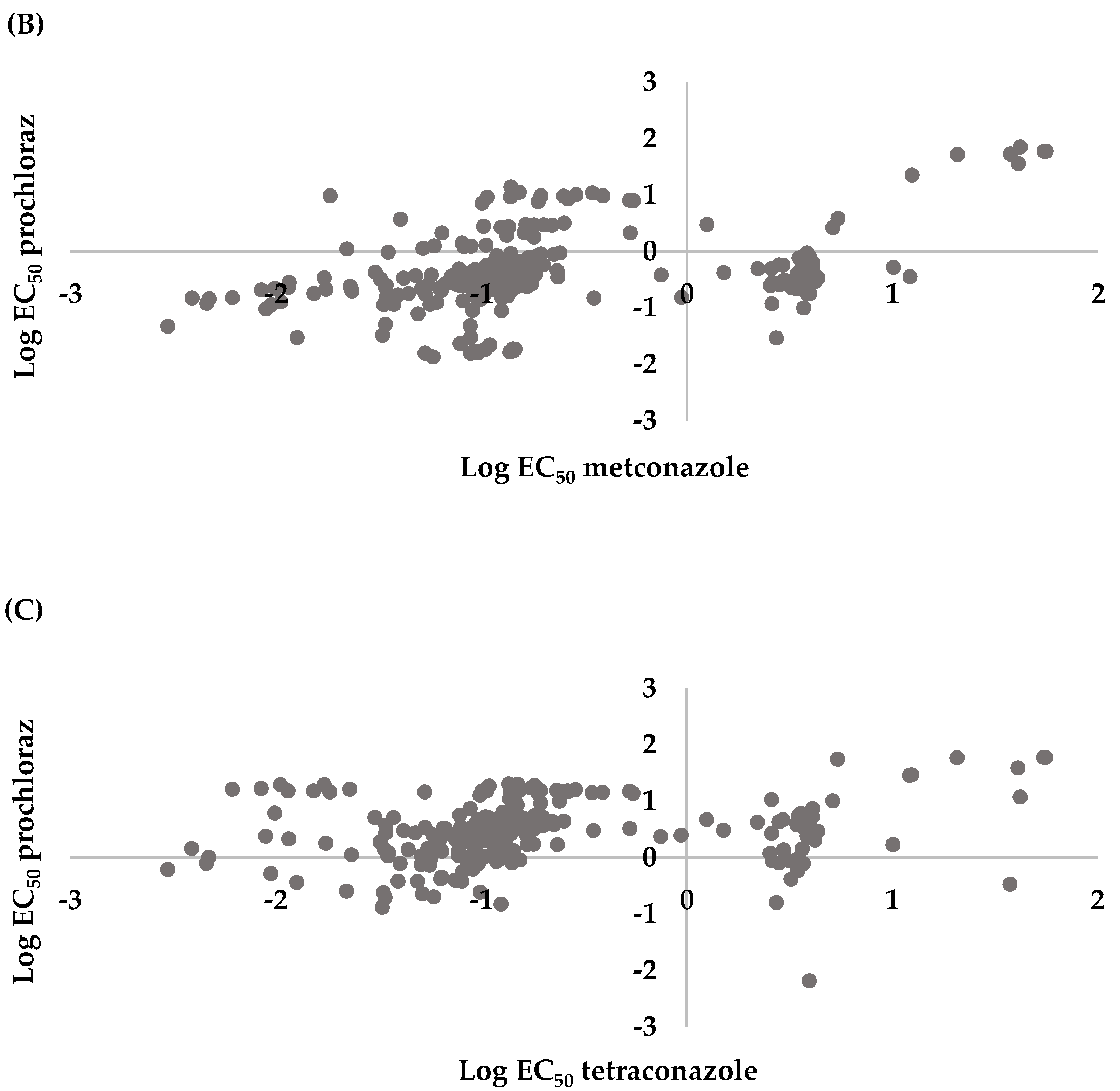
3.3. Cross Sensitivity
3.4. Differences between Fungicide Sensitivity Monitoring Methods
3.5. Promoter Insertions
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Russell, P.E. Fungicide Resistance: Occurrence and Management. J. Agric. Sci. 1995, 124, 317–323. [Google Scholar] [CrossRef]
- Parker, J.E.; Warrilow, A.G.; Price, C.L.; Mullins, J.G.; Kelly, D.E.; Kelly, S.L. Resistance to Antifungals that Target CYP51. J. Chem. Biol. 2014, 7, 143–161. [Google Scholar] [CrossRef] [PubMed]
- Marichal, P.; Gorrens, J.; Laurijssens, L.; Vermuyten, K.; Van Hove, C.; Le Jeune, L.; Verhasselt, P.; Sanglard, D.; Borgers, M.; Ramaekers, F.C.; et al. Accumulation of 3-Ketosteroids Induced by Itraconazole in Azole-Resistant Clinical Candida albicans Isolates. Antimicrob. Agents Chemother. 1999, 43, 2663–2670. [Google Scholar] [CrossRef]
- Czech Statistical Office. Areas under Crops. Available online: https://vdb.czso.cz/vdbvo2/faces/en/index.jsf?page=vystup-objektparametry&katalog=30840&z=T&sp=A&skupId=346&pvo=ZEM02A (accessed on 22 October 2021).
- Mazáková, J.; Urban, J.; Zouhar, M.; Ryšánek, P. Analysis of Leptosphaeria Species Complex Causing Phoma Leaf Spot and Stem Canker of Winter Oilseed Rape (Brassica napus) in the Czech Republic. Crop Pasture Sci. 2017, 68, 254–264. [Google Scholar] [CrossRef]
- Fitt, B.D.L.; Brun, H.; Barbetti, M.J.; Rimmer, S.R. World-Wide Importance of Phoma Stem Canker (Leptosphaeria maculans and L. biglobosa) on Oilseed Rape (Brassica napus). Eur. J. Plant Path. 2006, 114, 3–15. [Google Scholar] [CrossRef]
- Shoemaker, R.A.; Brun, H. The Teleomorph of the Weakly Aggressive Segregate of Leptosphaeria maculans. Can. J. Bot. 2001, 79, 412–419. [Google Scholar] [CrossRef]
- West, J.S.; Kharbanda, P.D.; Barbetti, M.J.; Fitt, B.D.L. Epidemiology and Management of Leptosphaeria maculans (Phoma Stem Canker) on Oilseed Rape in Australia, Canada and Europe. Plant Path. 2001, 50, 10–27. [Google Scholar] [CrossRef]
- Central Institute for Supervising and Testing in Agriculture. Available online: http://eagri.cz/public/app/eagriapp/POR/Vyhledavani.aspx?type=0&vyhledat=A&stamp=1566295194615 (accessed on 23 July 2021).
- Berry, P.M.; Spink, J.H. Understanding the Effect of a Triazole with Anti-Gibberellin Activity on the Growth and Yield of Oilseed Rape (Brassica napus). J. Agric. Sci. 2009, 147, 273–285. [Google Scholar] [CrossRef]
- Davis, T.D.; Steffens, G.L.; Sankhla, N. Triazole Plant Growth Regulators. Hortic. Rev. 1988, 10, 63–105. [Google Scholar]
- Fletcher, R.A.; Hofstra, G.; Gao, J. Comparative Fungitoxic and Plant Growth Regulating Properties of Triazole Derivatives. Plant Cell Physiol. 1986, 27, 367–371. [Google Scholar]
- Luster, D.G.; Miller, P.A. Triazole Plant Growth Regulator Binding to Native and Detergent-Solubilized Plant Microsomal Cytochrome P450. Pestic. Biochem. Physiol. 1993, 46, 27–39. [Google Scholar] [CrossRef]
- Coules, A.E.; Lunn, G.D.; Rossal, S. Disease and Canopy Control in Oilseed Rape Using Triazole Fungicides. In Proceedings of the BCPC Conference: Pests and Diseases, Brighton, UK, 18–21 November 2002. [Google Scholar]
- Zamani-Noor, N.; Knüfer, J. Effects of Host Plant Resistance and Fungicide Application on Phoma Stem Canker, Growth Parameters and Yield of Winter Oilseed Rape. Crop Prot. 2018, 112, 313–321. [Google Scholar] [CrossRef]
- Hollomon, D.W. Fungicide Resistance: Facing the Challenge. Plant Prot. Sci. 2015, 51, 170–176. [Google Scholar] [CrossRef]
- Ma, Z.; Michailides, T.J. Advances in Understanding Molecular Mechanisms of Fungicide Resistance and Molecular Detection of Resistant Genotypes in Phytopathogenic Fungi. Crop Prot. 2005, 24, 853–863. [Google Scholar] [CrossRef]
- Brent, K.J.; Hollomon, D.W. Fungicide Resistance in Crop Pathogens: How Can it Be Managed, 2nd ed.; Fungicide Resistance Action Committee; Crop Life International: Brussels, Belgium, 2007; p. 60. [Google Scholar]
- Eckert, M.R.; Rossall, S.; Selley, A.; Fitt, B.D.L. Effects of Fungicides on in Vitro Spore Germination and Mycelial Growth of the Phytopathogens Leptosphaeria maculans and L. biglobosa (Phoma Stem Canker of Oilseed Rape). Pest Manag. Sci. 2010, 66, 396–405. [Google Scholar] [CrossRef]
- Sewell, T.R.; Hawkins, N.J.; Stotz, H.U.; Huang, Y.J.; Kelly, S.L.; Kelly, D.E.; Fraaije, B.; Fitt, B.D.L. Azole Sensitivity in Leptosphaeria Pathogens of Oilseed Rape: The Role of Lanosterol 14α-Demethylase. Sci. Rep. 2017, 7, 15849. [Google Scholar] [CrossRef] [PubMed]
- Van de Wouw, A.P.; Elliott, V.L.; Chang, S.; López-Ruiz, F.J.; Marcroft, S.J.; Idnurm, A. Identification of Isolates of the Plant Pathogen Leptosphaeria maculans with Resistance to the Triazole Fungicide Fluquinconazole Using a Novel in Planta Assay. PLoS ONE 2017, 12, e0188106. [Google Scholar]
- Yang, Y.; Marcroft, S.J.; Forsyth, L.M.; Zhao, J.; Ziqin, L.; Van de Wouw, A.P.; Idnurm, A. Sterol Demethylation Inhibitor Fungicide Resistance in Leptosphaeria maculans is Caused by Modifications in the Regulatory Region of ERG11. Plant Dis. 2020, 104, 1280–1290. [Google Scholar] [CrossRef]
- Price, C.L.; Parker, J.E.; Warrilow, A.G.S.; Kelly, D.E.; Kelly, S.L. Azole Fungicides Understanding Resistance Mechanisms in Agricultural Fungal Pathogens. Pest Manag. Sci. 2015, 8, 1054–1058. [Google Scholar] [CrossRef]
- Huang, Y.J.; Hood, J.; Eckert, M.R.; Stonard, J.F.; Cools, H.J.; King, G.J.; Rosall, S.; Ashworth, F.; Fitt, B.D.L. Effects of Fungicide on Growth of Leptosphaeria maculans and L. biglobosa in Relation to Development of Phoma Stem Canker on Oilseed Rape (Brassica napus). Plant Path. 2011, 60, 607–620. [Google Scholar] [CrossRef]
- Williams, R.H.; Fitt, B.D.L. Differentiating A and B Groups of Leptosphaeria maculans, Causal Agent of Stem Canker (Blackleg) of Oilseed Rape. Plant Pathol. 1999, 48, 161–175. [Google Scholar] [CrossRef]
- Liu, S.Y.; Liu, Z.; Fitt, B.D.L.; Evans, N.; Foster, S.J.; Huang, Y.-J.; Latunde-Dada, A.O.; Lucas, J.A. Resistance to Leptosphaeria maculans (Phoma Stem Canker) in Brassica napus (Oilseed Rape) Induced by L. biglobosa and Chemical Defence Activators in Field and Controlled Environments. Plant Pathol. 2006, 55, 401–412. [Google Scholar] [CrossRef]
- Mahuku, G.S.; Hall, R.; Goodwin, P.H. Co-Infection and Induction of Systemic Acquired Resistance by Weakly and Highly Virulent Isolates of Leptosphaeria maculans in Oilseed Rape. Physiol. Mol. Plant Pathol. 1996, 49, 61–72. [Google Scholar] [CrossRef]
- Pijls, C.F.N.; Shaw, M.W.; Parker, A.A. Rapid Test to Evaluate in Vitro Sensitivity of Septoria tritici to Flutriafol, Using a Microtitre Plate Reader. Plant Pathol. 1994, 43, 726–732. [Google Scholar] [CrossRef]
- Wong, F.P.; Midland, S.L. Sensitivity Distributions of California Populations of Colletotrichum cereale to the DMI Fungicides Propiconazole, Myclobutanil, Tebuconazole, and Triadimefon. Plant Dis. 2007, 91, 1547–1555. [Google Scholar] [CrossRef] [PubMed]
- Wong, F.P.; Wilcox, W.F. Sensitivity to Azoxystrobin among Isolates of Uncinula necator: Baseline Distribution and Relationship to Myclobutanil Sensitivity. Plant Dis. 2002, 86, 394–404. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ishii, H.; Hollomon, D.W. Fungicide Resistance in Plant Pathogens: Principles and a Guide to Practical Management, 1st ed.; Springer: Berlin/Heilderberg, Germany, 2015; p. 490. [Google Scholar]
- Fraser, M.; Hwang, S.-F.; Ahmed, H.U.; Akhavan, A.; Stammler, G.; Barton, W.; Strelkov, S.E. Sensitivity of Leptosphaeria maculans to Pyraclostrobin in Alberta, Canada. Can. J. Plant Sci. 2016, 97, 83–91. [Google Scholar] [CrossRef]
- Wang, Y.; Akhavan, A.; Hwang, S.-F.; Strelkov, S.E. Decreased Sensitivity of Leptosphaeria maculans to Pyraclostrobin in Alberta, Canada. Plant Dis. 2020, 104, 2462–2468. [Google Scholar] [CrossRef]
- Van de Wouw, A.P.; Scanlan, J.L.; Marcroft, S.J.; Smith, A.J.; Sheedy, E.M.; Perndt, N.W.; Harrison, C.E.; Forsyth, L.M.; Idnurm, A. Fungicide Sensitivity and Resistance in the Blackleg Fungus, Leptosphaeria maculans, across Canola Growing Regions in Australia. Crop Pasture Sci. 2021, 72, 994–1007. [Google Scholar] [CrossRef]
- Délye, C.; Laigret, F.; Corio-Costet, M.F. A Mutation in the 14 Alpha-Demethylase Gene of Uncinula necator that Correlates with Resistance to a Sterol Biosynthesis Inhibitor. Appl. Environ. Microbiol. 1997, 63, 2966–2970. [Google Scholar] [CrossRef] [PubMed]
- Erickson, E.O.; Wilcox, W.F. Distributions of Sensitivities to Three Sterol Demethylation Inhibitor Fungicides among Populations of Uncinula necator Sensitive and Resistant to Triadimefon. Phytopathology 1997, 87, 784–791. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Fraaije, B.A.; Cools, H.J.; Kim, S.H.; Motteram, J.; Clark, W.S.; Lucas, J.A. A Novel Substitution I381V in the Sterol 14α-Demethylase (CYP51) of Mycosphaerella graminicola Is Differentially Selected by Azole Fungicides. Mol. Plant Pathol. 2007, 8, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Ghosoph, J.M.; Schmidt, L.S.; Margosan, D.A.; Smilanick, J.L. Imazalil Resistance Linked to a Unique Insertion Sequence in the PdCYP51 Promoter Region of Penicillium digitatum. Postharvest Biol. Technol. 2007, 44, 9–18. [Google Scholar] [CrossRef]
- Omrane, S.; Sghyer, H.; Audéon, C.; Lanen, C.; Duplaix, C.; Walker, A.S.; Fillinger, S. Fungicide Efflux and the MgMFS1 Transporter Contribute to the Multidrug Resistance Phenotype in Zymoseptoria tritici Field Isolates. Environ. Microbiol. 2015, 17, 2805–2823. [Google Scholar] [CrossRef] [PubMed]
- Stergiopoulos, I.; Van Nistelrooy, J.G.; Kema, G.H.; De Ward, M.A. Multiple Mechanisms Account for Variation in Base-Line Sensitivity to Azole Fungicides in Field Isolates of Mycosphaerella graminicola. Pest Manag. Sci. 2003, 59, 1333–1343. [Google Scholar] [CrossRef]
- Dyer, P.S.; Hansen, J.; Delaney, A.; Lucas, J.A. Genetic Control of Resistance to the Sterol 14alpha-Demethylase Inhibitor Fungicide Prochloraz in the Cereal Eyespot Pathogen Tapesia yallundae. Appl. Environ. Microbial. 2000, 66, 4599–4604. [Google Scholar] [CrossRef]
- Smith, F.D.; Parker, D.M.; Köller, W. Sensitivity Distribution of Venturia ynaequalis to Sterol Demethylation Inhibitor Flusilazole: Baseline Sensitivity and Implications for Resistance Monitoring. Phytopathology 1991, 81, 392–396. [Google Scholar] [CrossRef]
- Wang, W.; Fang, Y.; Imran, M.; Hu, Z.; Zhang, S.; Huang, Z.; Liu, X. Characterization of the Field Fludioxonil Resistance and Its Molecular Basis in Botrytis cinerea from Shanghai Province in China. Microorganisms 2021, 9, 266. [Google Scholar] [CrossRef] [PubMed]
- Ishii, H.; Cools, H.J.; Nishimura, K.; Borghi, L.; Kikuhara, K.; Yamaoka, Y. DMI-Fungicide Resistance in Venturia nashicola, the Causal Agent of Asian Pear Scab—How Reliable Are Mycelial Growth Tests in Culture? Microorganisms 2021, 9, 1377. [Google Scholar] [CrossRef] [PubMed]
- Wedge, D.E.; Curry, K.J.; Kreiser, B.; Curry, A.; Abril, M.; Smith, B.J. Fungicide Resistance Profiles for 13 Botrytis cinerea Isolates from Strawberry in Southeastern Louisiana. Int. J. Fruit Sci. 2013, 13, 413–429. [Google Scholar] [CrossRef]
- Brent, K.J.; Hollomon, D.W. Fungicide Resistance: The Assessment of Risk Monograph, 2nd ed.; Global Crop Protection Federation: Brussels, Belgium, 1998; p. 53. [Google Scholar]
- Karaoglanidis, G.S.; Thanassoulopoulos, C.C. Cross-Resistance Patterns among Sterol Biosynthesis Inhibiting Fungicides (SBIs) in Cercospora beticola. Eur. J. Plant Pathol. 2003, 109, 929–934. [Google Scholar] [CrossRef]
- Thomas, A.; Langston, D.B., Jr.; Stevenson, K.L. Baseline Sensitivity and Cross-Resistance to Succinate-Dehydrogenase-Inhibiting and Demethylation Inhibiting Fungicides in Didymella bryoniae. Plant Dis. 2012, 96, 979–984. [Google Scholar] [CrossRef] [PubMed]
- Ishii, H.; Bryson, P.K.; Kayamori, M.; Miyamoto, T.; Yamaoka, Y.; Schnabel, G. Cross-Resistance to the New Fungicide Mefentrifluconazole in DMI-Resistant Fungal Pathogens. Pestic. Biochem. Physiol. 2021, 171, 104737. [Google Scholar]
- Patel, R.; Mendrick, C.; Knapp, C.; Grist, R.; McNicholas, P. Clinical Evaluation of the Sensititre Yeast One Plate for Testing Susceptibility of Filamentous Fungi to Posaconazole. J. Clin. Microbiol. 2007, 45, 2000–2001. [Google Scholar] [CrossRef]
- Schnabel, G.; Jones, A.L. The 14α−Demethylase (CYP51A1) Gene Is Overexpressed in Venturia inaequalis Strains Resistant to Myclobutanil. Phytopathology 2001, 91, 102–110. [Google Scholar] [CrossRef]
- Ma, Z.; Proffer, T.J.; Jacobs, J.L.; Sundin, G.W. Overexpression of the 14alpha-Demethylase Target Gene (CYP51) Mediates Fungicide Resistance in Blumeriella jaapii. Appl. Environ. Microbiol. 2006, 72, 2581–2585. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region | Years | N | EC50 (µg mL−1) | ||
|---|---|---|---|---|---|
| Tetraconazole | Metconazole | Prochloraz | |||
| Central Bohemian | 2016–2017 | 3 | 1.277–1.542 | 0.754–1.5 | 0.7707–1.463 |
| Hradec Králové | 2017 | 6 | 0.2411–1.753 | 0.453–1.717 | 1.03–1.155 |
| Moravian-Silesian | 2016–2017 | 8 | 1.143–4.808 | 0.7088–1.729 | 0.1596–1.06 |
| Olomouc | 2016 | 7 | 1.028–1.531 | 0.3169–1.398 | 0.035–0.5293 |
| Praha | 2017 | 2 | 1.3116–1.436 | 0.281–0.6564 | 0.1578–0.16 |
| South Moravian | 2015–2016 | 5 | 1.065–1.332 | 0.0602–1.451 | 0.0706–0.7707 |
| Zlín | 2014–2017 | 14 | 0.7776–1.756 | 0.06905–1.671 | 0.0225–1.224 |
| Region | Years | N | EC50 (µg mL−1) | ||
|---|---|---|---|---|---|
| Tetraconazole | Metconazole | Prochloraz | |||
| Central Bohemian | 2016–2017 | 11 | 0.1471–14.74 | 0.02971–8.105 | 0.01266–0.9403 |
| Hradec Králové | 2017, 2019 | 100 | 0.2524–58.62 | 0.09946–52.58 | 0.003895–20.7 |
| Karlovy Vary | 2019 | 36 | 1.172–19.93 | 0.1278–7.582 | 0.00613–3.833 |
| Moravian-Silesian | 2016, 2017, 2019 | 88 | 0.006585–38.3 | 0.0136–70.69 | 0.004717–41.95 |
| Olomouc | 2016–2017 | 7 | 0.1953–14.77 | 0.05094–9.548 | 0.034–3.474 |
| Plzeň | 2017 | 9 | 0.8578–6.38 | 0.04847–2.959 | 0.01156–0.1804 |
| Praha | 2017 | 7 | 0.7815–14.08 | 0.1198–9.347 | 0.00459–5.133 |
| South Moravian | 2015, 2016, 2017 | 19 | 0.3739–59.41 | 0.07853–59.59 | 0.03428–56.04 |
| Zlín | 2016-2017 | 9 | 0.1302–58.84 | 0.03295–59.28 | 0.002977–54.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fajemisin, O.; Mazáková, J.; Ryšánek, P. Emergence of Fungicide Sensitivity in Leptosphaeria maculans Isolates Collected from the Czech Republic to DMI Fungicides. Agriculture 2022, 12, 237. https://doi.org/10.3390/agriculture12020237
Fajemisin O, Mazáková J, Ryšánek P. Emergence of Fungicide Sensitivity in Leptosphaeria maculans Isolates Collected from the Czech Republic to DMI Fungicides. Agriculture. 2022; 12(2):237. https://doi.org/10.3390/agriculture12020237
Chicago/Turabian StyleFajemisin, Olufadekemi, Jana Mazáková, and Pavel Ryšánek. 2022. "Emergence of Fungicide Sensitivity in Leptosphaeria maculans Isolates Collected from the Czech Republic to DMI Fungicides" Agriculture 12, no. 2: 237. https://doi.org/10.3390/agriculture12020237
APA StyleFajemisin, O., Mazáková, J., & Ryšánek, P. (2022). Emergence of Fungicide Sensitivity in Leptosphaeria maculans Isolates Collected from the Czech Republic to DMI Fungicides. Agriculture, 12(2), 237. https://doi.org/10.3390/agriculture12020237