
Achieving the European Green “Deal” of Sustainable Grass Forage Production and Landscaping Using Fungal Endophytes

 ,
,  ,
,  ,
,
Abstract
1. Introduction
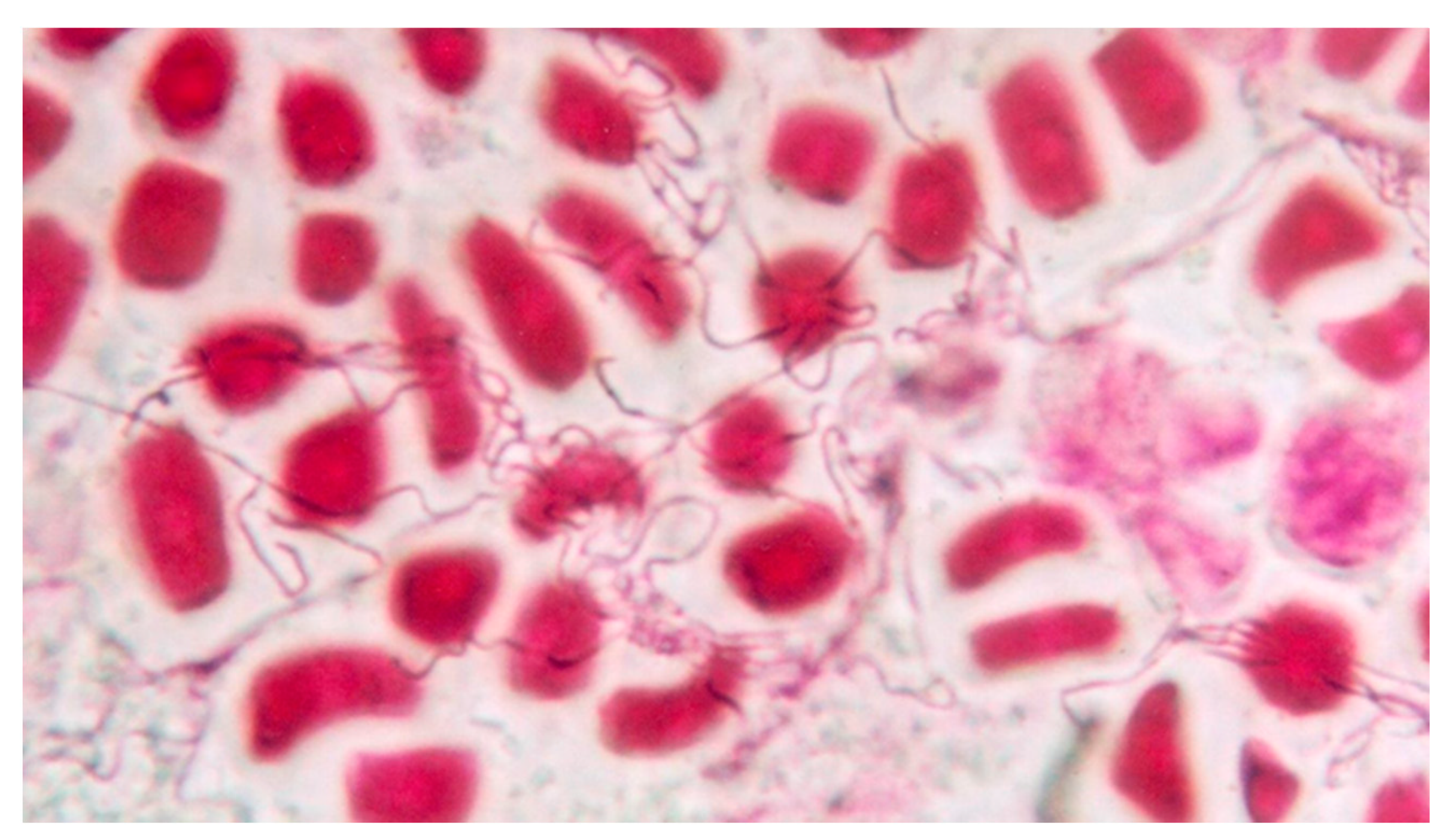
2. Epichloid Endophytes of Grasses
3. Grass-Endophyte Symbiosis Enhance Host Tolerance to Drought Stress
4. Mineral Nutrients from Soil Are More Efficiently Used by Grass–Endophyte Associations
5. Presence of Epichloid Endophytes Protects Grasses against Pathogens
6. Grasses with Epichloid Endophytes Are More Resistant to Pest Infestation
7. Forage Quality Can Be Negatively Affected by Endophytes
8. Symbiotically Modified Grasses
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Eriksson, O.; Cousins, S.A.O.; Bruun, H.H. Land-use history and fragmentation of traditionally managed grassland in Scandinavia. J. Veg. Sci. 2002, 13, 743–748. [Google Scholar] [CrossRef]
- Huyghe, C.; De Vliegher, A.; Van Gils, B.; Peeters, A. Grasslands and herbivore production in Europe and effects of common policies. In Les Editions Quae; Editions Quae: Versailles, France, 2014; p. 320. [Google Scholar] [CrossRef]
- Gunnarsson, B.; Knez, I.; Sang, M.H. Effects of biodiversity and environment-related attitude on perception of urban greenspace. Urban Ecosyst. 2017, 20, 37–49. [Google Scholar] [CrossRef]
- Clay, K.; Schardl, C. Evolutionary origins and ecological consequences of endophyte symbiosis with grasses. Am. Nat. 2002, 160, S99–S127. [Google Scholar] [CrossRef]
- West, C.P. Physiology and drought tolerance of endophyte-infected grasses. In Biotechnology of Endophytic Fungi of Grasses; Bacon, C.W., White, J.F., Jr., Eds.; CRC Press: Boca Raton, FL, USA, 1994; pp. 87–99. [Google Scholar]
- Tian, P.; Nan, Z.; Li, C.; Spangenberg, G. Effect of endophyte Neotyphodium lolii on susceptibility and host physiological response of perennial ryegrass to fungal pathogens. Eur. J. Plant Pathol. 2008, 122, 593–602. [Google Scholar] [CrossRef]
- Johnson, M.C.; Dahlman, D.L.; Siegel, M.R.; Bush, L.P.; Latch, G.C.; Potter, D.A.; Varney, D.R. Insect feeding deterrents in endophyte-infected tall fescue. Appl. Environ. Microbiol. 1985, 49, 568–571. [Google Scholar] [CrossRef] [PubMed]
- Bush, L.P.; Wilkinson, H.H.; Schardl, C.L. Bioprotective alkaloids of grass–fungal endophyte symbioses. Plant Physiol. 1997, 114, 1–7. [Google Scholar] [CrossRef]
- Leuchtmann, A. Systematics, distribution, and host specificity of grass endophytes. Nat. Toxins 1992, 1, 150–162. [Google Scholar] [CrossRef]
- Zabalgogeazcoa, I.; Vazquez de Aldana, B.R.; García Ciudad, A.; García Criado, B. Fungal endophytes in grasses from semiarid permanent grasslands of Western Spain. Grass Forage Sci. 2003, 58, 94–97. [Google Scholar] [CrossRef]
- Schardl, C.L.; Leuchtman, A.; Spiering, M.J. Symbioses of grasses with seed borne fungal endophytes. Annu. Rev. Plant Biol. 2004, 55, 315–340. [Google Scholar] [CrossRef] [PubMed]
- Lembicz, M.; Górzyńska, K.; Leuchtmann, A. Choke disease, caused by Epichloë bromicola, in the grass Agropyron repens in Poland. Plant Dis. 2010, 94, 1372. [Google Scholar] [CrossRef] [PubMed]
- Żurek, M.; Wiewióra, B.; Żurek, G.; Prończuk, M. Occurrence of endophyte fungi on grasses in Poland—Review. Fungal Ecol. 2012, 5, 353–356. [Google Scholar] [CrossRef]
- Faeth, S.H. Are endophytic fungi defensive plant mutualists? Oikos 2002, 98, 25–36. [Google Scholar] [CrossRef]
- Young, C.A.; David, E.H.; Mc Culley, R.L. Fungal endophytes of tall fescue and perennial ryegrass: Pasture friend or foe? J. Anim Sci. 2013, 91, 2379–2394. [Google Scholar] [CrossRef]
- Christensen, M.J.; Leuchtmann, A.; Rowan, D.D.; Tapper, B.A. Taxonomy of Acremonium endophytes of tall fescue (Festuca arundinacea), meadow fescue (Festuca pratensis) and perennial ryegrass (Lolium perenne). Mycol. Res. 1993, 97, 1083–1092. [Google Scholar] [CrossRef]
- Leuchtmann, A.; Bacon, C.W.; Schardl, C.L.; White, J.F., Jr.; Tadych, M. Nomenclatural realignment of Neotyphodium species with genus Epichloë. Mycologia 2014, 106, 202–215. [Google Scholar] [CrossRef] [PubMed]
- Zabalgogeazcoa, I.; Vázquez de Aldana, B.R.; Criado, B.G.; Ciudad, A.G. The infection of Festuca rubra by the fungal endophyte Epichloë festucae in Mediterranean permanent grasslands. Grass Forage Sci. 1999, 54, 91–95. [Google Scholar] [CrossRef]
- Saikkonen, K.; Ahlholm, J.; Helander, M.; Lehtimäki, S.; Niemeläinen, O. Endophytic fungi in wild and cultivated grasses in Finland. Ecography 2000, 23, 360–366. [Google Scholar] [CrossRef]
- Saikkonen, K.; Wäli, P.R.; Helander, M.; Faeth, S.H. Evolution of endophyte-plant symbioses. Trends Plant Sci. 2004, 9, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Saikkonen, K.; Lehtonen, P.; Helander, M.; Koricheva, J.; Faeth, S.H. Model systems in ecology: Dissecting the grass-endophyte literature. Trends Plant Sci. 2006, 11, 428–433. [Google Scholar] [CrossRef] [PubMed]
- Marshall, D.; Tunali, B.; Nelson, R.L. Occurrence of fungal endophytes in species of wild Triticum. Crop Sci. 1999, 39, 1507–1512. [Google Scholar] [CrossRef]
- Tunali, B.; Marshall, D. Antagonistic effect of endophytes against several root-rot pathogens of wheat. In Durum Wheat Improvement in the Mediterranean Region: New Challenges; Royo, C., Nachit, M.M., Di Fonzo, N., Araus, J.L., Eds.; CIHEAM-IAMZ, Options Méditerranéennes: Série A; Séminaires Méditerranéens; CIHEAM: Zaragoza, Spain, 2000; Volume 40, pp. 381–386. [Google Scholar]
- Simpson, W.R.; Faville, M.J.; Moraga, R.A.; Williams, W.M.; McManus, M.T.; Johnson, W.R. Epichloë fungal endophytes and the formation of synthetic symbioses in Hordeeae (=Triticeae) grasses. J. Syst. Evol. 2014, 56, 794–806. [Google Scholar] [CrossRef]
- Wilson, A.D. Clavicipitaceous anamorphic endophytes in Hordeum germplasm. Plant Pathol. J. 2007, 6, 1–13. [Google Scholar] [CrossRef]
- Card, S.D.; Faville, M.J.; Simpson, W.R.; Johnson, R.D.; Voisey, C.R.; De Bonth, A.C.M.; Hume, D.E. Mutualistic fungal endophytes in the Triticeae—Survey and description. Microbiol. Ecol. 2014, 88, 94–106. [Google Scholar] [CrossRef] [PubMed]
- Moon, C.D.; Craven, K.D.; Leuchtmann, A.; Clement, S.L.; Schardl, C.L. Prevalence of inter specific hybrids amongst asexual fungal endophytes of grasses. Mol. Ecol. 2004, 13, 1455–1467. [Google Scholar] [CrossRef] [PubMed]
- Selosse, M.A.; Schardl, C.L. Fungal endophytes of grasses: Hybrids rescued by vertical transmission? An evolutionary perspective. New Phytol. 2007, 173, 452–458. [Google Scholar] [CrossRef] [PubMed]
- Malinowski, D.P.; Belesky, D.P. Adaptations of endophyte-infected cool-season grasses to environmental stresses: Mechanisms of drought and mineral stress tolerance. Crop Sci. 2000, 40, 923–940. [Google Scholar] [CrossRef]
- Malinowski, D.P.; Belesky, D.P.; Lewis, G.C. Abiotic stresses in endophytic grasses. In Neotyphodium in Cool-Season Grasses; Roberts, C., West, C.P., Spiers, D., Eds.; Blackwell Publishing Ltd.: Oxford, UK, 2005; pp. 187–199. [Google Scholar] [CrossRef]
- Cheplick, G.P.; Faeth, S. Ecology and Evolution of the Grass Endophyte Symbiosis; Oxford University Press: New York, NY, USA, 2009. [Google Scholar] [CrossRef]
- Arraudeau, M.A. Breeding strategies for drought resistance. In Drought Resistance in Cereals; Baker, F.W.C., Ed.; CAB International: Wallingford, UK, 1989; pp. 107–116. [Google Scholar]
- Elmi, A.A.; West, C.P. Endophyte infection effects on stomatal conductance, osmotic adjustment and drought recovery of tall fescue. New Phytol. 1995, 131, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Buck, G.W.; West, C.P.; Elbersen, H.W. Endophyte effect on drought tolerance in diverse Festuca species. In Neotyphodium/Grass Interactions, 1st ed.; Bacon, C.W., Hill, N.S., Eds.; Plenum Press: New York, NY, USA, 1997; pp. 141–143. [Google Scholar] [CrossRef]
- Malinowski, D.; Leuchtmann, A.; Schmidt, D.; Nösberger, J. Growth and water status in meadow fescue is affected by Neotyphodium and Phialophora species endophytes. Agron. J. 1997, 89, 673–678. [Google Scholar] [CrossRef]
- Malinowski, D.; Leuchtmann, A.; Schmidt, D.; Nösberger, J. Symbiosis with Neotyphodium uncinatum endophyte may increase the competitive ability of meadow fescue. Agron J. 1997, 89, 833–839. [Google Scholar] [CrossRef]
- Johnson, R.C.; Tieszen, L.L. Carbon isotope discrimination, water relations, and gas exchange in temperate grass species and accessions. In Stable Isotopes and Plant Carbon Water Relations; Ehleringer, J.R., Hall, A.E., Farquhar, G.D., Eds.; Academic Press, Inc.: San Diego, CA, USA, 1993; pp. 281–296. [Google Scholar] [CrossRef]
- Eerens, J.P.J.; Lucas, R.J.; Easton, H.S.; White, J.G.H. Influence of the endophyte Neotyphodium lolii on morphology, physiology, and alkaloid synthesis of perennial ryegrass during high temperature and water stress. New Zealand J. Agric. Res. 1998, 41, 219–226. [Google Scholar] [CrossRef]
- Richardson, M.D.; Chapman, G.W., Jr.; Hoveland, C.S.; Bacon, C.W. Sugar alkohols in endophyte infected tall fescue. Crop Sci. 1992, 32, 1060–1061. [Google Scholar] [CrossRef]
- Elbersen, H.W.; West, C.P. Growth and water relations of field grown tall fescue as influenced by drought and endophyte. Grass Forage Sci. 1996, 51, 333–342. [Google Scholar] [CrossRef]
- Bayat, F.; Mirlohi, A.; Khodambashi, M. Effects of endophytic fungi on some drought tolerance mechanisms of tall fescue in a hydroponics culture. Russ. J. Plant Physiol. 2009, 56, 510–516. [Google Scholar] [CrossRef]
- Nagabhyru, P.; Dinkins, R.D.; Wood, C.L.; Bacon, C.W.; Schardl, C.L. Tall fescue endophyte effects on tolerance to water deficit stress. BMC Plant Biol. 2013, 13, 127. [Google Scholar] [CrossRef]
- White, R.H.; Engelke, M.C.; Morton, S.J.; Johnson-Cicalese, J.M.; Ruemmele, B.A. Acremonium endophyte effects on tall fescue drought tolerance. Crop Sci. 1992, 32, 1392–1396. [Google Scholar] [CrossRef]
- Malinowski, D. Rhizomatous Ecotypes and Symbiosis with Endophytes as New Possibilities of Improvement in Competitive Ability of Meadow Fescue (Festuca pratensis Huds). Ph.D. Thesis, Swiss Federal Institute of Technology, Zurich, Switzerland, 1995. [Google Scholar] [CrossRef]
- Barker, D.J.; Sullivan, C.Y.; Moser, L.E. Water deficit effects on osmotic potential, cell wall elasticity and proline in five forage grasses. Agron. J. 1993, 85, 270–275. [Google Scholar] [CrossRef]
- Abernethy, G.A.; McManus, M.T. Biochemical responses to an imposed water deficit in mature leaf tissue of Festuca arundinacea. Environ. Exp. Bot. 1998, 40, 17–28. [Google Scholar] [CrossRef]
- West, C.P.; Carson, R.D.; Guerber, C.A.; de los Reyes, B. Endophyte effects on antioxidants and membrane leakage in tall fescue during drought. In Proceedings of the 4th International Symposium on the Molecular Breeding of Forage and Turf, a Satellite Workshop of the XXth International Grassland Congress, Aberystwyth, Wales, 3–7 July 2005; Academic Publishers: Wageningen, The Netherlands, 2005; pp. 217–218. [Google Scholar]
- White, J.F.; Torres, M.S. Is plant endophyte mediated defensive mutualism the result of oxidative stress protection? Physiol. Plant. 2009, 138, 440–446. [Google Scholar] [CrossRef] [PubMed]
- Qawasmeh, A.; Obied, H.K.; Raman, A.; Wheatley, W. Influence of fungal endophyte infection on phenolic content and antioxidant activity in grasses: Interaction between Lolium perenne and different strains of Neotyphodium lolii. J. Agric. Food Chem. 2012, 60, 3381–3388. [Google Scholar] [CrossRef] [PubMed]
- Torres, M.S.; White, J.F., Jr.; Zhang, X.; Hinton, D.M.; Bacon, C.W. Endophyte-mediated adjustments in host morphology and physiology and effects on host fitness traits in grasses. Fungal Ecol. 2012, 5, 322–330. [Google Scholar] [CrossRef]
- Volaire, F. Drought survival, summer dormancy and dehydrin accumulation in contrasting cultivars of Dactylis glomerata. Physiol. Plant. 2002, 116, 42–51. [Google Scholar] [CrossRef]
- Allagulova, C.R.; Gimalov, F.R.; Shakirova, F.M.; Vakhitov, V.A. The plant dehydrins: Structure and putative functions. Biochemistry 2003, 68, 945–951. [Google Scholar] [CrossRef] [PubMed]
- Hume, D.E.; Ryan, G.D.; Gibert, A.; Helander, M.; Mirlohi, A.; Sabzalian, M.R. Epichloë Fungal Endophytes for Grassland Ecosystems. In Sustainable Agriculture Reviews, Sustainable Agriculture Reviews 19; Lichtfouse, E., Ed.; Springer International Publishing: Cham, Switzerland, 2016. [Google Scholar] [CrossRef]
- Belesky, D.P.; Wilkinson, S.R.; Pallas, J.E., Jr. Response of four tall fescue cultivars grown at two nitrogen levels to low soil water availability. Crop Sci. 1982, 22, 93–97. [Google Scholar] [CrossRef]
- Belesky, D.P.; Wilkinson, S.R.; Evans, J.J. Amino acid composition of fractions of ‘Kentucky-31′ tall fescue as affected by N fertilization and mild water stress. Plant Soil. 1984, 81, 257–267. [Google Scholar] [CrossRef]
- Lyons, P.C.; Evans, J.J.; Bacon, C.W. Effects of the fungal endophyte Acremonium coenophialum on nitrogen accumulation and metabolism in tall fescue. Plant Physiol. 1990, 92, 726–732. [Google Scholar] [CrossRef]
- Malinowski, D.P.; Alloush, G.A.; Belesky, D.P. Evidence for chemical changes on the root surface of tall fescue in response to infection with the fungal endophyte Neotyphodium coenophialum. Plant Soil. 1998, 205, 1–12. [Google Scholar] [CrossRef]
- Malinowski, D.P.; Belesky, D.P. Tall fescue aluminum tolerance is affected by Neotyphodium coenophialum endophyte. J. Plant Nutr. 1999, 22, 1335–1349. [Google Scholar] [CrossRef]
- Malinowski, D.P.; Belesky, D.P. Neotyphodium coenophialum endophyte infection affects the ability of tall fescue to use sparingly available phosphorus. J. Plant Nutr. 1999, 22, 835–853. [Google Scholar] [CrossRef]
- Malinowski, D.P.; Alloush, G.A.; Belesky, D.P. Leaf endophyte Neotyphodium coenophialum modifies mineral uptake in tall fescue. Plant Soil. 2000, 227, 115–126. [Google Scholar] [CrossRef]
- Malinowski, D.P.; Zuo, H.; Belesky, D.P.; Alloush, G.A. Evidence for copper binding by extracellular root exudates of tall fescue but not perennial ryegrass infected with Neotyphodium spp. endophytes. Plant Soil. 2004, 267, 1–12. [Google Scholar] [CrossRef]
- Zabalgogeazcoa, I.; García Ciudad, A.; Vázquez de Aldana, B.R.; García Criado, B. Effects of the infection by the fungal endophyte Epichloë festucae in the growth and nutrient content of Festuca rubra. Eur. J. Agron. 2006, 24, 374–384. [Google Scholar] [CrossRef]
- Latch, G.C.M.; Christensen, M.J.; Samuels, G.J. Five endophytes of Lolium and Festuca in New Zealand. Mycotaxon 1984, 20, 535–550. [Google Scholar]
- Guo, J.; McCulley, R.L.; McNear, D.H., Jr. Tall fescue cultivar and fungal endophyte combinations influence plant growth and root exudate composition. Front. Plant Sci. 2015, 6, 183. [Google Scholar] [CrossRef]
- Ren, A.Z.; Gao, Y.B.; Zhou, F. Response of Neotyphodium lolii infected perennial ryegrass to phosphorus deficiency. Plant Soil Environ. 2007, 53, 113–119. [Google Scholar] [CrossRef]
- Soleimani, M.; Hajabbasi, M.A.; Afyuni, M.; Mirlohi, A.; Borggaard, O.K.; Holm, P.E. Effect of endophytic fungi on cadmium tolerance and bioaccumulation by Festuca arundinacea and Festuca pratensis. Int. J. Phytoremediation. 2010, 12, 535–549. [Google Scholar] [CrossRef] [PubMed]
- Mirzahosseini, Z.; Shabani, L.; Sabzalian, M.R.; Sharifi, T.M. Effect of Neotyphodium endophyte symbiosis on growth, nickel uptake and photosynthetic pigments in two genotypes of tall fescue. J. Plant Process. Funct. 2014, 2, 25–37. [Google Scholar]
- Zamani, N.; Sabzalian, M.R.; Khoshgoftarmanesh, A.; Afyuni, M. Neotyphodium endophyte changes phytoextraction of zinc in Festuca arundinacea and Lolium perenne. Int. J. Phytoremediation 2014, 17, 456–463. [Google Scholar] [CrossRef] [PubMed]
- Shimanuki, T.; Sato, T. Occurrence of the choke disease on timothy caused by Epichloë typhina (Pers.ex Fr.) Tul. in Hokkaido and location of the endophytic mycelia with plant tissue (JE). Res. Bull. Hok-kaido Natl. Agric. Exp Stn 1983, 138, 87–97. [Google Scholar]
- Vincelli, P.; Powell, A.J. Reaction of perennial ryegrass varieties to red thread. Biol. Cultural Tests Control Plant Dis. 1991, 6, 102. [Google Scholar]
- Schmidt, D. Du nouveau sur les endophytes de la fétuque des prés. Rev. Suisse Agric. 1994, 26, 57–63. [Google Scholar]
- Koshino, H.; Togiya, S.; Yoshihara, T.; Sakamura, S. Four fungitoxic C-18 hydroxy unsaturated fatty acids from stromata of Epichloë typhina. Tetrahedron Lett. 1987, 28, 73–76. [Google Scholar] [CrossRef]
- Pańka, D. Susceptibility of meadow fescue (Festuca pratensis Huds.) infected with endophyte Neotyphodium uncinatum to pathogens. In Recent Advances in Genetics and Breeding of the Grasses; Zwierzykowski, Z., Kosmala, A., Eds.; Institute of Plant Genetics PAS: Poznań, Poland, 2005; pp. 191–195. [Google Scholar]
- Clarke, B.B.; White, J.F., Jr.; Hurley, R.H.; Torres, M.S.; Sun, S.; Huff, D.R. Endophyte-mediated suppression of dollar spot dieases in fine fescues. Plant Dis. 2006, 90, 994–998. [Google Scholar] [CrossRef]
- Lehtonen, P.M.; Helander, M.; Siddiqui, S.A.; Lehto, K.; Saikkonen, K. Endophytic fungi decrease plant virus infections in meadow ryegrass (Lolium pratense). Bio. Lett. 2006, 2, 620–623. [Google Scholar] [CrossRef][Green Version]
- Wäli, P.R.; Helander, M.; Nissinen, O.; Saikkonen, K. Susceptibility of endophyte-infected grasses to winter pathogens (snow molds). Can. J. Bot. 2006, 84, 1043–1051. [Google Scholar] [CrossRef]
- Reddy, M.N.; Faeth, S.H. Damping-off of Festuca arizonica caused by Fusarium. Am. J. Plant Sci. 2010, 1, 104–105. [Google Scholar] [CrossRef]
- Wiewióra, B.; Żurek, G.; Żurek, M. Endophyte-mediated disease resistance in wild populations of perennial ryegrass (Lolium perenne). Fungal Ecol. 2015, 15, 1–8. [Google Scholar] [CrossRef]
- Christensen, M.J. Antifungal activity in grassess infected with Acremonium and Epichloë endophytes. Australas. Plant Pathol. 1996, 25, 186–191. [Google Scholar] [CrossRef]
- Pańka, D. Infestation of tall fescue (Festuca arundinaceae Schreb.) with Neotyphodium coenophialum and its influence on growth of chosen microorganisms in vitro. Acta Agrobot. 2005, 58, 369–380. [Google Scholar] [CrossRef][Green Version]
- Pańka, D. Occurrence of Neotyphodium lolii and its antifungal properties. Phytopathol. Pol. 2008, 48, 5–12. [Google Scholar]
- Burpee, L.L.; Bouton, J.H. Effect of eradication of the endophyte Acremonium coenophialum on epidemics of Rhizoctonia blight in tall fescue. Plant Dis. 1993, 77, 157–159. [Google Scholar] [CrossRef]
- Trevathan, L.E. Performance of endophyte-free and endophyte infected tall fescue seedlings in soil infected with Cochliobolus sativus. Can. J. Plant Pathol. 1996, 18, 415–418. [Google Scholar] [CrossRef]
- Hamilton, C.E.; Faeth, S.H. Asexual Neotyphodium endophytes in Arizona fescue: A test of the seed germination and pathogen resistance hypothesis. Symbiosis 2005, 38, 69–85. [Google Scholar]
- Pańka, D.; West, C.P.; Guerber, C.A.; Richardson, M.D. Susceptibility of tall fescue to Rhizoctonia zeae infection as affected by endophyte symbiosis. Ann. Appl. Biol. 2013, 163, 257–268. [Google Scholar] [CrossRef]
- Moy, M.; Belanger, F.C.; Duncan, R.; Freehof, A.; Leary, C.; Meyer, W.; Sullivan, R.; White, J.F. Identification of epiphyllous mycelial nets on leaves of grasses infected by Clavicipitaceous endophytes. Symbiosis 2000, 28, 291–302. [Google Scholar]
- Moy, M.; Li, H.M.; Sullivan, R.; White, J.F., Jr.; Belanger, F.C. Endophytic fungal β-1,6 gluconase expression in the infected host grass. Plant Physiol. 2002, 130, 1298–1308. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Neill, S.J.; Desikan, R.; Clarke, A.; Hurst, R.D.; Hancock, J.T. Hydrogen peroxide and nitric oxide as signaling molecules in plants. J. Exp. Bot. 2002, 53, 1237–1247. [Google Scholar] [CrossRef]
- Hohnjec, N.; Vieweg, M.F.; Pühler, A.; Becker, A.; Küster, H. Overlaps in the transcriptional profiles of Medicago truncatula roots inoculated with two different Glomus fungi provide insights into the genetic program activated during arbuscular mycorrhiza. Plant Physiol. 2005, 137, 1283–1301. [Google Scholar] [CrossRef] [PubMed]
- Ford, V.L.; Kirkpatrick, T.L. Effects of Acremonium coenophialum in tall fescue on host disease and insect resistance and allelopathy to Pinus taeda seedlings. Ark. Agric. Exp. Stn. Special Rep. 1989, 140, 29–34. [Google Scholar]
- Stewart, T.M.; Mercer, C.F.; Grant, J.L. Development of Meloidogyne nassi on endophyte-infected and endophyte-free perennial ryegrass. Australas. Plant Pathol. 1993, 22, 40–41. [Google Scholar] [CrossRef]
- Eerens, J.P.J.; Visker, M.H.P.W.; Lucas, R.J.; Easton, H.S.; White, J.G.H. Influence of the ryegrass endophyte (Neotyphodium lolii) in a cool moist environment: IV. Plant Parasitic nematodes. New Zealand J. Agric. Res. 1998, 41, 209–217. [Google Scholar] [CrossRef][Green Version]
- Pennell, C.G.; Ball, O.J.-P. The effects of Neotyphodium endophytes in tall fescue on pasture mealy bug (Balanococcus poae). In Proceedings of the New Zealand Plant Protection Conference, Auckland, New Zealand, 10–12 August 1999; Volume 52, pp. 259–263. [Google Scholar]
- Elmi, A.A.; West, C.P.; Robbins, R.T.; Kirkpatrik, T.L. Endophyte effects on reproduction of root-knot nematode (Meloidogyne merylandi) and osmotic adjustment in tall fescue. Grass Forage Sci. 2000, 55, 166–172. [Google Scholar] [CrossRef]
- Pennell, C.G.; David, E.H.; Ball, O.J.; Easton, H.S.; Lyons, T.B. Effects of Neotyphodium lolii infection in ryegrass on root aphid and pasture mealy bug. In Proceedings of the 4th International Neotyphodium/Grass Interactions Symposium, Soest, Germany, 27–29 September 2000; Paul, V.H., Dapprich, P.D., Eds.; Universitat-Gesamthochschule Paderborn: Paderborn, Germany, 2000; pp. 465–469. [Google Scholar]
- Popay, A.J.; Baltus, J.G.; Pennell, C.G. Insect resistance in perennial ryegrass infected with toxin-free Neotyphodium endophytes. In Proceedings of the 4th International Neotyphodium/Grass Interactions Symposium, Soest, Germany, 27–29 September 2000; Paul, V.H., Dapprich, P.D., Eds.; Universitat-Gesamthochschule Paderborn: Paderborn, Germany, 2000; pp. 187–193. [Google Scholar]
- Siegel, M.R.; Bush, L.P. Defensive chemicals in grass-fungal endophyte associations. Recent Adv. Phytochem. 1996, 30, 81–119. [Google Scholar]
- Siegel, M.R.; Bush, L.P. Toxin production in grass/endophyte association. In The Mycota. V. Plant relationships, Part A; Carroll, G.C., Tudzynski, P., Eds.; Springer: Berlin, Germany, 1997; pp. 185–208. [Google Scholar]
- Raisbeck, M.F.; Rottinghaus, G.E.; Kendall, J.D. Effects of naturally occurring mycotoxins on ruminants. In Mycotoxins and Animal Foods; Smith, J.E., Henderson, R.S., Eds.; CRC Press: Boca Raton, FL, USA, 1991; pp. 647–677. [Google Scholar]
- Thompson, F.N.; Stuedemann, J.A. Pathophysiology of fescue toxicosis. Agric. Ecosyst. Environ. 1993, 44, 263–281. [Google Scholar] [CrossRef]
- Tor-Agbidye, J.; Blythe, L.L.; Craig, A.M. Correlation of endophyte toxins (ergovaline and lolitrem B) with clinical disease: Fescue foot and perennial ryegrass staggers. Vet. Hum. Toxicol. 2001, 43, 140–146. [Google Scholar] [PubMed]
- Eich, E.; Pertz, H. Antimicrobial and antitumor effects of ergot alkaloids and their derivatives. In Ergot: The Genus Claviceps; Kren, V., Cvak, L., Eds.; Academic Publishers: Amsterdam, The Netherlands, 1999; pp. 441–449. [Google Scholar]
- Wilkinson, H.H.; Siegel, M.R.; Blankenship, J.D.; Mallory, A.C.; Bush, L.P.; Schardl, C.L. Contribution of fungal loline alkaloids to protection from aphids in a grass endophyte mutualism. Mol. Plant Microbe Interact. 2000, 13, 1027–1033. [Google Scholar] [CrossRef] [PubMed]
- Spiering, M.J.; Moon, C.D.; Wilkinson, H.H.; Schardl, C.L. Gene clusters for insecticidal loline alkaloids in the grass–endophytic fungus Neotyphodium uncinatum. Genetics 2005, 169, 1403–1414. [Google Scholar] [CrossRef] [PubMed]
- Huizing, H.J.; Van der Molen, W.; Kloek, W.; Den Nijs, A.P.M. Detection of lolines in endophyte-containing meadow fescue in the Netherlands and the effect of elevated temperature on induction of lolines in endophyte-infected perennial ryegrass. Grass Forage Sci. 1999, 46, 441–445. [Google Scholar] [CrossRef]
- Riedell, W.E.; Kieckhefer, R.E.; Petroski, R.J.; Powell, R.G. Naturally occurring and synthetic loline alkaloid derivatives: Insect feeding behavior modification and toxicity. J. Entomol. Sci. 1991, 26, 122–129. [Google Scholar] [CrossRef]
- Rowan, D.D.; Hunt, M.B.; Gaynor, D.L. Peramine, a novel insect feeding deterrent from ryegrass infected with the endophyte Acremonium loliae. J. Chem. Soc. Chem. Commun. 1986, 142, 935–936. [Google Scholar] [CrossRef]
- Latch, G.C.M. Physiological interactions of endophytic fungi and their hosts: Biotic stress tolerance imparted to grasses by endophytes. Agric. Ecosyst. Environ. 1993, 44, 143–156. [Google Scholar] [CrossRef]
- Siegel, M.; Latch, G.; Bush, L.; Fannin, F.; Rowan, D.; Tapper, B. Fungal endophyte infected grasses: Alkaloid accumulation and aphid response. J. Chem. Ecol. 1990, 16, 3301–3315. [Google Scholar] [CrossRef]
- Vázquez de Aldana, B.R.; Zabalgogeazcoa, I.; Rubio de Casas, R.; García Ciudad, A.; García Criado, B. Relationships between the genetic distance of Epichloë festucae isolates and the ergovaline and peramine content of their Festuca rubra hosts. Ann. Appl. Biol. 2010, 156, 51–61. [Google Scholar] [CrossRef]
- Lehtonen, P.; Helander, M.; Saikkonen, K. Are endophyte-mediated effects on herbivores conditional on soil nutrients? Oecology 2005, 142, 38–45. [Google Scholar] [CrossRef]
- Tapper, B.A.; Lane, G.A. Janthitrems found in a Neotyphodium endophyte of perennial ryegrass. In Proceedings of the 5th International Symposium on Neotyphodium/Grass Interactions, 23–26 May 2004; Kallenbach, R., Rosenkrans, C.J., Lock, T.R., Eds.; University of Arkansas Press: Fayetteville, AR, USA, 2004; p. 301. [Google Scholar]
- Babu, J.V.; Popay, A.J.; Miles, C.O.; Wilkins, A.L.; di Menna, M.E.; Finch, S.C. Identification and structure elucidation of janthitrems A and D from Penicillium janthinellum and determination of the tremorgenic and anti-insect activity of janthitrems A and B. J. Agric. Food Chem. 2018, 66, 13116–13125. [Google Scholar] [CrossRef] [PubMed]
- Ball, O.J.P.; Christensen, M.J.; Prestidge, R.A. Effect of selected isolates of Acremonium endophytes on adult black beetle (Heteronychus arator) feeding. N. Z. Plant Prot. 1994, 47, 227–231. [Google Scholar] [CrossRef]
- Popay, A.J.; Wyatt, R.T. Resistance to argentine stem weevil in perennial ryegrass infected with endophytes producing different alkaloids. N. Z. Plant Prot. 1995, 48, 229–236. [Google Scholar] [CrossRef]
- Jensen, J.G.; Popay, A.J. Perennial ryegrass infected with AR37 endophyte reduces survival of porina larvae. N. Z. Plant Prot. 2004, 57, 323–328. [Google Scholar] [CrossRef]
- Popay, A.; Silvester, W.B.; Gerard, P.J. New endophyte isolate suppresses root aphid, Aploneura lentisci, in perennial ryegrass. In Proceedings of the 5th International Symposium on Neotyphodium/Grass Interactions 23–26 May 2004; Kallenbach, R., Rosenkrans, C.J., Lock, T.R., Eds.; University of Arkansas Press: Fayetteville, AR, USA, 2004; p. 317. [Google Scholar]
- Endophytes and Related Methods. WO2011082455A1, 2011. Available online: http://patents.google.com/patent/WO2011082455A1/en (accessed on 27 February 2021).
- Rowan, D.D. Lolitrems, peramine and paxilline: Mycotoxins of the ryegrass/endophyte interaction. Agric. Ecosyst. Environ. 1993, 44, 103–122. [Google Scholar] [CrossRef]
- Di Menna, M.E.; Finch, S.C.; Popay, A.J.; Smith, B.L. A review of the Neotyphodium lolii/Lolium perenne symbiosis and its associated effects on animal and plant health, with particular emphasis on ryegrass staggers. N. Z. Vet. J. 2012, 60, 315–328. [Google Scholar] [CrossRef]
- Strickland, J.R.; Spiers, D.E.; Oliver, J.W.; Aiken, G.E. Physiological basis for fescue toxicosis. In Tall Fescue for the Twenty-First Century; Fribourg, H.A., Hannaway, D.B., West, C.P., Eds.; Agron. monograph; American Society of Agronomy, Crop Science Society of America, Soil Science Society of America: Madison, WI, USA, 2009; pp. 203–227. [Google Scholar]
- Waller, J.C. Endophyte effects on cattle. In Tall Fescue for the Twenty-First Century; Fribourg, H.A., Hannaway, D.B., West, C.P., Eds.; Agron. monograph; American Society of Agronomy, Crop Science Society of America, Soil Science Society of America: Madison, WI, USA, 2009; pp. 289–310. [Google Scholar]
- Oldenburg, E. Effects of endophyte-infected perennial ryegrass on the performance of sheep. Rev. Med. Vet. 1998, 149, 638. [Google Scholar]
- Lewis, G.C. Significance of endophyte toxicosis and current practices in dealing with the problem in Europe. In Neotyphodium/Grass Interactions; Bacon, C.W., Hill, N.S., Eds.; Plenum Press: New York, NY, USA, 1997; pp. 377–382. [Google Scholar]
- Lewis, G.C. An overview of the symbiotic association between Neotyphodium/Epichloë fungal endophytes and forage grasses. Asp. Appl. Biol. 2001, 63, 203–212. [Google Scholar]
- Bony, S.; Delatour, P. Relevance and impact of grass endophyte toxins in Europe. In Proceedings of the 4th International Neotyphodium/Grass Interactions Symposium, Soest, Germany, 27–29 September 2000; Paul, V.H., Dapprich, P.D., Eds.; Universitat-Gesamthochschule Paderborn: Paderborn, Germany, 2000; pp. 207–218. [Google Scholar]
- Zabalgogeazcoa, I.; Bony, S. Neotyphodium research and application in Europe. In Neotyphodium in Cool-Season Grasses; Roberts, C., West, C.P., Spiers, D., Eds.; Blackwell Publishing: Ames, IA, USA, 2005; pp. 23–33. [Google Scholar]
- Ball, O.J.; Prestidge, R.A.; Sprosen, J.M. Interrelationships between Acremonium lolii, peramine, and lolitrem B in perennial ryegrass. Appl. Environ. Microbiol. 1995, 61, 1527–1533. [Google Scholar] [CrossRef] [PubMed]
- Keogh, R.G.; Tapper, B.A.; Fletcher, R.H. Distributions of the fungal endophyte Acremonium lolii, and of the alkaloids lolitrem B and peramine, within perennial ryegrass. New Zealand J. Agric. Res. 1996, 39, 121–127. [Google Scholar] [CrossRef]
- Easton, H.S.; Latch, G.C.M.; Tapper, B.A.; Ball, O.J.P. Ryegrass host genetic control of concentrations of endophyte-derived alkaloids. Crop Sci. 2002, 42, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Żurek, G.; Wiewióra, B.; Żurek, M.; Łyszczarz, R. Environmental effect on Epichloë endophyte occurrence and ergovaline concentration in wild populations of forage grasses in Poland. Plant Soil 2017, 410, 383–399. [Google Scholar] [CrossRef]
- Gundel, P.E.; Pérez, L.I.; Helander, M.; Saikkonen, K. Symbiotically modified organisms: Nontoxic fungal endophytes in grasses. Trends Plant Sci. 2013, 18, 420–427. [Google Scholar] [CrossRef]
- Bouton, J.; Easton, S. Endophytes in forage cultivars. In Neotyphodium in Cool-Season Grasses; Roberts, C.A., West, C.P., Spiers, D.E., Eds.; Blackwell Publishing: Ames, Iowa, USA, 2005; pp. 327–340. [Google Scholar]
- Brilman, L.A. Endophytes in turfgrass cultivars. In Neotyphodium in Cool-Season Grasses; Roberts, C.A., West, C.P., Spiers, D.E., Eds.; Blackwell Publishing: Ames, Iowa, USA, 2005; pp. 341–349. [Google Scholar]
- MVS Mountain View Seeds. Available online: https://www.mtviewseeds.com/forage-tall-fescue (accessed on 2 March 2021).
- Pennington. Available online: https://www.pennington.com/all-products/agriculture/forage (accessed on 2 March 2021).
- Barenbrug New Zealand. Available online: https://www.barenbrug.co.nz/endophyte/endophyte-options.htm (accessed on 2 March 2021).
- Barenbrug Australia. Available online: https://barenbrug.com.au/forage-pasture/pasture-grasses/perennial-ryegrass (accessed on 2 March 2021).
- PGG Wrightson Seeds. Available online: https://www.pggwrightsonseeds.com/pasture/grasses (accessed on 2 March 2021).
- Spinoni, J.; Vogt, J.V.; Naumann, G.; Barbosa, P.; Dosio, A. Will drought events become more frequent and severe in Europe? Int. J. Climatol. 2017, 38, 1718–1736. [Google Scholar] [CrossRef]
- Patterson, J.D.; Lafaillette, F.; Wöster, S.; Roulund, N.; Charrier, S.; Gilliland, T.J. Impact of endophyte inoculation on the morphological identity of cultivars of Lolium perenne (L) and Festuca arundinacea (Schreb.). Sci. Rep. 2020, 10, 7729. [Google Scholar] [CrossRef] [PubMed]
- Kyriakides, S. The food chain: Paving the way to formulating a more sustainable, healthy and quality food policy. In Farm to Fork: Towards a Sustainable and Resilient Food System in Europe; Ulmann, L., Mai Thanh, C., Eds.; The European Files/Les Dossiers Europeenes: Brussels, Belgium, 2020; Volume 64, p. 6. [Google Scholar]
- Wojciechowski, J. Towards a sustainable and resilient food system in Europe—The key role of the Common Agricultural Policy. In Farm to Fork: Towards a sustainable and resilient food system in Europe; Ulmann, L., Mai Thanh, C., Eds.; The European Files/Les Dossiers Europeenes: Brussels, Belgium, 2020; Volume 64, p. 7. [Google Scholar]



{kind=link}
{kind=link}
| Grass Cultivar | Novel Endophyte | Reference |
|---|---|---|
| “Estancia” 1, “Teton II” 1 | Ark Shield | [135] |
| “Jesup” 1 | MaxQ (AR542) | [136] |
| “Jesup” 1, “Lacefield” 1, “Texoma” 1 | MaxQ II (AR584) | [136] |
| “Shogun” 2 | NEA | [137] |
| “Bealey” 2, “Rohan” 2, “Trojan” 2, “Impact 2” 2, “Kidman” 2 | NEA2 | [137,138] |
| “Maxsyn” 2, “Viscount” 2 | NEA4 | [137,138] |
| “Tyson” 2, “Governor” 2 | AR1 | [137] |
| “Base” 2, “Excess” 2, “Governor” 2, “Rely” 2, “Platform” 2, “Lush” 3 | AR37 | [137,139] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pańka, D.; Jeske, M.; Łukanowski, A.; Prus, P.; Szwarc, K.; Muhire, J.d.D. Achieving the European Green “Deal” of Sustainable Grass Forage Production and Landscaping Using Fungal Endophytes. Agriculture 2021, 11, 390. https://doi.org/10.3390/agriculture11050390
Pańka D, Jeske M, Łukanowski A, Prus P, Szwarc K, Muhire JdD. Achieving the European Green “Deal” of Sustainable Grass Forage Production and Landscaping Using Fungal Endophytes. Agriculture. 2021; 11(5):390. https://doi.org/10.3390/agriculture11050390
Chicago/Turabian StylePańka, Dariusz, Małgorzata Jeske, Aleksander Łukanowski, Piotr Prus, Katarzyna Szwarc, and Jean de Dieu Muhire. 2021. "Achieving the European Green “Deal” of Sustainable Grass Forage Production and Landscaping Using Fungal Endophytes" Agriculture 11, no. 5: 390. https://doi.org/10.3390/agriculture11050390
APA StylePańka, D., Jeske, M., Łukanowski, A., Prus, P., Szwarc, K., & Muhire, J. d. D. (2021). Achieving the European Green “Deal” of Sustainable Grass Forage Production and Landscaping Using Fungal Endophytes. Agriculture, 11(5), 390. https://doi.org/10.3390/agriculture11050390