Reduced Nitrogen Rate with Increased Planting Density Facilitated Grain Yield and Nitrogen Use Efficiency in Modern Conventional Japonica Rice


Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Site, Rice Variety, Field Design, and Crop Establishment
2.2. Sampling and Measurement
2.3. Formula Calculation and Statistical Analysis
3. Results
3.1. Grain Yield and Its Components
3.2. Shoot Biomass Weight and Accumulation, Harvest Index, and NSC Content
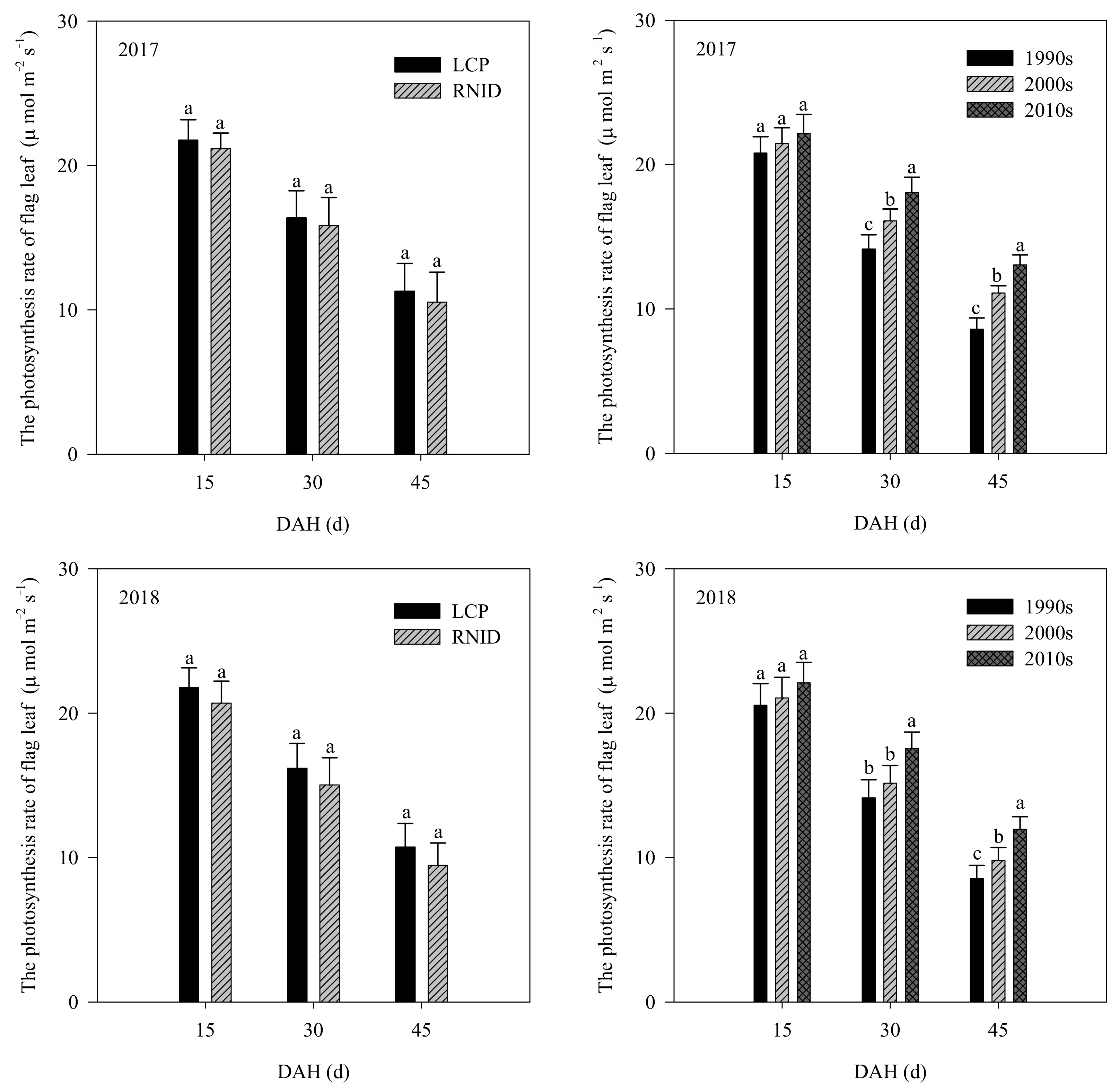
3.3. LAI, SPAD Values, and Leaf Photosynthetic Rate
3.4. N, P, and K Accumulation and Their Utilization Efficiencies
3.5. Correlation Analysis
4. Discussion
4.1. The Main Traits for the Progressive Yield Increase in Conventional japonica Rice since the 1990s
4.2. The Feasibility of RNID Facilitating Grain Yield and NUE of Modern Conventional Japonica Rice
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Emily, E. Rice by the numbers: A good grain. Nature 2014, 514, S50–S51. [Google Scholar]
- Bai, X.F.; Wu, B.; Xing, Y.Z. Yield-related QTLs and their applications in rice genetic improvement. J. Intergr. Plant Biol. 2012, 54, 300–311. [Google Scholar] [CrossRef]
- Yu, Y.Q.; Huang, Y.; Zhang, W. Changes in rice yields in China since 1980 associated with cultivar improvement, climate and crop management. Field Crop. Res. 2012, 136, 65–75. [Google Scholar] [CrossRef]
- Breseghello, F.; De Morais, O.P.; Pinheiro, P.V.; Silva, A.C.S.; De Castro, E.D.M.; Guimarães, É.P.; De Castro, A.P.; Pereira, J.A.; Lopes, A.D.M.; Utumi, M.M.; et al. Results of 25 years of upland rice breeding in Brazil. Crop Sci. 2011, 51, 914–923. [Google Scholar] [CrossRef]
- Futakuchi, K.; Senthilkumar, K.; Arouna, A.; Vandamme, E.; Diagne, M.; Zhao, D.L.; Manneh, B.; Saito, K. History and progress in genetic improvement for enhancing rice yield in sub-Saharan Africa. Field Crop. Res. 2021, 267, 108159. [Google Scholar] [CrossRef]
- Anzoua, K.G.; Junichi, K.; Toshihiro, H.; Kazuto, I.; Yutaka, J. Genetic improvements for high yield and low soil nitrogen tolerance in rice (Oryza Sativa L.) under a cold environment. Field Crop. Res. 2010, 116, 38–45. [Google Scholar] [CrossRef][Green Version]
- Liu, L.J.; Xiong, Y.W.; Bian, J.L.; Zhang, H.; Gu, J.F.; Wang, Z.Q.; Yang, J.C. Effect of genetic improvement of grain yield and nitrogen efficiency of mid-season indica rice cultivars. J. Plant Nutr. Soil Sci. 2015, 178, 297–305. [Google Scholar] [CrossRef]
- Peng, S.; Laza, R.C.; Visperas, R.M.; Sanico, A.L.; Cassman, K.G.; Khush, G.S. Grain yield of rice cultivars and lines developed in the Philippines since 1966. Crop Sci. 2000, 40, 307–314. [Google Scholar] [CrossRef]
- Zhu, G.L.; Ren, Z.; Liu, Y.Q.; Lu, F.G.; Gu, L.F.; Shi, Y.; Liu, J.W.; Zhou, G.S.; Nimir, N.E.A.; Mohapatra, P.K. Optimization of leaf properties and plant phenotype through yield-based genetic improvement of rice over a period of seventy years in the Yangtze River Basin of China. Food Energy Secur. 2020, 9, e223. [Google Scholar] [CrossRef]
- Meng, T.Y.; Ge, J.L.; Zhang, X.B.; Chen, X.; Zhou, G.S.; Wei, H.H. Improvements in plant morphology facilitating progressive yield increases of japonica inbred rice since the 1980s in East China. Agriculture 2021, 11, 834. [Google Scholar] [CrossRef]
- Wu, Z.H.; Xu, K.Z.; Zhao, Y.J.; He, X.L.; Wang, X.L.; Ling, F.L. Changes of some agronomic traits in japonica rice varieties during forty-seven years of genetic improvement in Jilin province, China. Chin. J. Rice Sci. 2007, 21, 507–512. (In Chinese) [Google Scholar]
- Zhang, H.C.; Zhang, J.; Gong, J.L.; Chang, Y.; Li, M.; Gao, H.; Dai, Q.G.; Huo, Z.Y.; Xu, K.; Wei, H.Y. The productive advantages and formation mechanisms of “indica rice to japonica rice”. Sci. Agric. Sin. 2013, 46, 686–704. [Google Scholar]
- Wang, C.L.; Zhang, Y.D.; Zhu, Z.; Chen, T.; Zhao, Q.Y.; Zhong, W.G.; Yang, J.; Yao, S.; Zhou, L.H.; Zhao, L.; et al. Research progress on the breeding of japonica super rice varieties in Jiangsu Province, China. J. Integr. Agric. 2017, 16, 992–999. [Google Scholar] [CrossRef]
- Parry, M.A.J.; Hawkesford, M.J. Food security: Increasing yield and improving resource use efficiency. Proc. Nutr. Soc. 2010, 69, 592–600. [Google Scholar] [CrossRef]
- Hwakesford, M.J. Genetic variation in traits for nitrogen use efficiency in wheat. J. Exp. Bot. 2017, 68, 2627–2632. [Google Scholar] [CrossRef]
- Tian, Z.W.; Li, Y.; Liang, Z.H.; Guo, H.; Cai, J.; Jiang, D.; Cao, W.X.; Dai, T.B. Genetic improvement of nitrogen uptake and utilization of winter wheat in the Yangtze River Basin of China. Field Crop. Res. 2016, 196, 251–260. [Google Scholar] [CrossRef]
- Haegele, J.W.; Cook, K.A.; Nichols, D.M.; Below, F.E. Changes in nitrogen use traits associated with genetic improvement for grain yield of maize hybrids released in different decades. Crop Sci. 2013, 53, 1256–1268. [Google Scholar] [CrossRef]
- Mueller, S.M.; Messina, C.D.; Vyn, T.J. Simultaneous gains in grain yield and nitrogen efficiency over 70 years of maize genetic improvement. Sci. Rep. 2019, 9, 9095. [Google Scholar] [CrossRef] [PubMed]
- Bingham, I.J.; Karley, A.J.; White, P.J.; Thomas, W.T.B.; Russell, J.R. Analysis of improvements in nitrogen use efficiency associated with 75 years of spring barley breeding. Eur. J. Agron. 2012, 42, 49–58. [Google Scholar] [CrossRef]
- Muurinen, S.; Slafer, G.A.; Peltonen-Sainio, P. Breeding effects on nitrogen use efficiency of spring cereals under northern conditions. Crop Sci. 2006, 46, 561–568. [Google Scholar] [CrossRef]
- Rochester, I.J.; Constable, G.A. Improvements in nutrient uptake and nutrient use-efficiency in cotton cultivars released between 1973 and 2006. Field Crop. Res. 2015, 173, 14–21. [Google Scholar] [CrossRef]
- Wei, H.H.; Meng, T.Y.; Ge, J.L.; Zhang, X.B.; Shi, T.Y.; Ding, E.H.; Lu, Y.; Li, X.Y.; Tao, Y.; Chen, Y.L.; et al. Reduced nitrogen application rate with dense planting improves rice grain yield and nitrogen use efficiency: A case study in east China. Crop J. 2021, 9, 954–961. [Google Scholar] [CrossRef]
- Zheng, H.B.; Chen, Y.W.; Chen, Q.M.; Li, B.; Zhang, Y.S.; Jia, W.; Mo, W.W.; Tang, Q.Y. High-density planting with lower nitrogen application increased early rice production in a double-season rice system. Agron. J. 2020, 112, 205–214. [Google Scholar] [CrossRef]
- Hou, W.F.; Khan, M.R.; Zhang, J.L.; Lu, J.W.; Ren, T.; Cong, R.H.; Li, X.K. Nitrogen rate and plant density interaction enhances radiation interception, yield and nitrogen use efficiency of mechanically transplanted rice. Agric. Ecosyst. Environ. 2019, 269, 183–192. [Google Scholar] [CrossRef]
- Tian, G.L.; Gao, L.M.; Kong, Y.L.; Hu, X.Y.; Xie, K.L.; Zhang, R.Q.; Ling, N.; Shen, Q.R.; Guo, S.W. Improving rice population productivity by reducing nitrogen rate and increasing plant density. PLoS ONE 2017, 12, e0182310. [Google Scholar]
- Huang, M.; Chen, J.N.; Cao, F.B.; Zou, Y.B. Increased hill density can compensate for yield loss from reduced nitrogen input in machine-transplanted double-cropped rice. Field Crop. Res. 2018, 221, 333–338. [Google Scholar] [CrossRef]
- Chen, J.; Zhu, X.C.; Xie, J.; Deng, G.Q.; Tu, T.H.; Guan, X.J.; Yang, Z.; Huang, S.; Chen, X.M.; Qiu, C.F.; et al. Reducing nitrogen application with dense planting increases nitrogen use efficiency by maintaining root growth in a double-rice cropping system. Crop J. 2021, 9, 805–815. [Google Scholar] [CrossRef]
- Huang, M.; Zou, Y.B. Reducing environmental risk of nitrogen by popularizing mechanically dense transplanting for rice production in China. J. Integr. Agric. 2020, 19, 2362–2366. [Google Scholar] [CrossRef]
- Zhu, X.C.; Zhang, J.; Zhang, Z.P.; Deng, A.X.; Zhang, W.J. Dense planting with less basal nitrogen fertilization might benefit rice cropping for high yield with less environmental impacts. Eur. J. Agron. 2016, 75, 50–59. [Google Scholar] [CrossRef]
- Hu, Y.J.; Qian, H.J.; Cao, W.W.; Xing, Z.P.; Zhang, H.C.; Dai, Q.G.; Huo, Z.Y.; Xu, K.; Wei, H.Y.; Guo, B.W. Effects of different mechanical transplantation methods and density on yield and its components of different panicle-typed rice. Chin. J. Rice Sci. 2016, 30, 493–506. (In Chinese) [Google Scholar]
- Xing, Z.P.; Wu, P.; Zhu, M.; Qian, H.J.; Cao, W.W.; Hu, Y.J.; Guo, B.W.; Wei, H.Y.; Xu, K.; Dai, Q.G.; et al. Effect of mechanized planting methods on plant type and lodging resistance of different rice varieties. Trans. CSAE 2017, 33, 52–62. (In Chinese) [Google Scholar]
- Nakano, H.; Morita, S.; Kitagawa, H.; Wada, H.; Takahashi, M. Grain yield response to planting density in forage rice with a large number of spikelets. Crop Sci. 2012, 52, 345–350. [Google Scholar] [CrossRef]
- Inthapanya, P.; Sipaseuth; Sihavong, P.; Sihathep, V.; Chanphengsay, M.; Fukai, S.; Basnayake, J. Genotype differences in nutrient uptake and utilisation for grain yield production of rainfed lowland rice under fertilised and non-fertilised conditions. Field Crop. Res. 2000, 65, 57–68. [Google Scholar] [CrossRef]
- Li, X.T.; Cheng, H.T.; Wang, N.; Yu, C.M.; Qu, L.Y.; Cao, P.; Hu, N.; Liu, T.; Lyu, W.Y. Critical factors for grain filling of erect panicle type japonica rice cultivars. Agron. J. 2013, 105, 1404–1410. [Google Scholar] [CrossRef]
- Mohapatra, P.K.; Panigrahi, R.; Turner, N.C. Chapter Five—Physiology of spikelet development on the rice panicle: Is manipulation of apical dominance crucial for grain yield improvement? Adv. Agron. 2011, 110, 333–359. [Google Scholar]
- Fu, P.H.; Wang, J.; Zhang, T.; Huang, J.L.; Peng, S.B. High nitrogen input causes poor grain filling of spikelets at the panicle base of super hybrid rice. Field Crop. Res. 2019, 244, 107635. [Google Scholar] [CrossRef]
- Panigrahi, R.; Kariali, E.; Panda, B.B.; Lafarge, T.; Mohapatra, P.K. Controlling the trade-off between spikelet number and grain filling: The hierarchy of starch synthesis in spikelets of rice panicle in relation to hormone dynamics. Funct. Plant Biol. 2019, 46, 507–523. [Google Scholar] [CrossRef]
- Keipp, K.; Hütsch, B.W.; Schubert, S. How does the harvest index affect water-use efficiency and nutrient-utilization efficiency of sunflowers (Helianthus annuus L.)? J. Agron. Crop Sci. 2019, 205, 519–532. [Google Scholar] [CrossRef]
- Morita, S.; Nakano, H. Nonstructural carbohydrate content in the stem at full heading contributes to high performance of ripening in heat-tolerant rice cultivar Nikomaru. Crop Sci. 2011, 51, 818–828. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Cultivation Treatment | Nitrogen Application Rate and Timing (kg ha−1) | Planting Density | ||||||
|---|---|---|---|---|---|---|---|---|
| Total Rate | One Day before Transplanting | One Week after Transplanting | Panicle Initiation | Penultimate Leaf Appearance | Hill Spacing | Seedling per Hill | Seeding Number per m2 | |
| LCP | 300 | 90 | 90 | 60 | 60 | 25 cm × 13.3 cm | 4 | 120 |
| RNID | 255 | 76.5 | 76.5 | 51 | 51 | 25 cm × 11.0 cm | 4 | 144 |
| Varietal Type | Variety | Year of Release | Cross Information | Cumulative Planting Area (×104 hm2) |
|---|---|---|---|---|
| 1990s | Wuyujing 3 | 1992 | Zhongdan 1/79-51 × Zhongdan 1/Yangjing 1 | 608 |
| Zaofeng 9 | 1997 | Wufujing/Zhongdan 1 × Nonglin 205 | 104 | |
| Wuyunjing 8 | 1999 | Xiangnuo 9121/Jia 48 × Bing 815 | 88 | |
| 2000s | Wujing 15 | 2004 | Zaofeng 9/Chunjiang03jing × Wuyunjing 7 | 88 |
| Nanjing 44 | 2007 | Nanjing 38 | 45 | |
| Yangjing 4038 | 2008 | Zhenxiang 24/Wuyunjing 8 × Chang 9363 | 22 | |
| 2010s | Nanjing 9108 | 2013 | Wuxiangjing 14 × Guandong 194 | 111 |
| Zhendao 18 | 2013 | Zhendao 99 × Wuyunjing 7 | 17 | |
| Wuyunjing 31 | 2015 | Yun 2608 × Fan 103 | 10 |
| Year | Treatment | Grain Yield (t ha−1) | Panicles per m2 | Spikelets per Panicle | Spikelets per m2 | Filled-Grain Percentage (%) | Grain Weight (mg) | |
|---|---|---|---|---|---|---|---|---|
| 2017 | Cultivation treatment | LCP | 11.1 a | 303 b | 155 a | 46.6 a | 89.4 a | 27.0 a |
| RNID | 10.9 a | 313 a | 146 b | 45.4 a | 89.9 a | 27.0 a | ||
| Varietal type | 1990s | 10.6 c | 327 a | 131 c | 42.6 c | 90.0 a | 27.4 a | |
| 2000s | 11.0 b | 311 b | 149 b | 46.3 b | 90.0 a | 27.2 a | ||
| 2010s | 11.5 a | 287 c | 172 a | 49.2 a | 88.9 a | 26.5 b | ||
| 2018 | Cultivation treatment | LCP | 11.1 a | 310 b | 153 a | 47.0 a | 88.6 a | 26.8 a |
| RNID | 10.9 b | 322 a | 143 b | 45.8 a | 89.9 a | 26.9 a | ||
| Varietal type | 1990s | 10.6 c | 336 a | 135 c | 45.3 c | 90.3 a | 27.2 a | |
| 2000s | 11.0 b | 319 a | 145 b | 46.2 b | 89.9 a | 26.8 ab | ||
| 2010s | 11.5 a | 293 b | 164 a | 47.8 a | 87.7 b | 26.6 b | ||
| Year | Treatment | Shoot Biomass Weight (t ha−1) | Shoot Biomass Accumulation (t ha−1) | Harvest Index | ||||
|---|---|---|---|---|---|---|---|---|
| Jointing | Heading | Maturity | From Jointing to Heading | From Heading to Maturity | ||||
| 2017 | Cultivation treatment | LCP | 5.5 a | 11.9 a | 20.1 a | 6.5 a | 8.1 a | 0.476 b |
| RNID | 5.4 a | 11.7 a | 19.6 b | 6.3 a | 7.9 a | 0.484 a | ||
| Varietal type | 1990s | 5.6 a | 11.7 a | 19.4 b | 6.1 b | 7.7 b | 0.471 b | |
| 2000s | 5.4 a | 11.8 a | 19.8 b | 6.5 a | 8.0 b | 0.482 a | ||
| 2010s | 5.4 a | 12.0 a | 20.4 a | 6.6 a | 8.4 a | 0.488 a | ||
| 2018 | Cultivation treatment | LCP | 5.5 a | 11.7 a | 19.9 a | 6.2 a | 8.2 a | 0.481 b |
| RNID | 5.3 a | 11.4 a | 19.3 b | 6.1 a | 7.9 b | 0.489 a | ||
| Varietal type | 1990s | 5.5 a | 11.4 a | 19.2 b | 5.9 a | 7.8 b | 0.477 b | |
| 2000s | 5.5 a | 11.6 a | 19.5 b | 6.1 a | 7.9 b | 0.488 a | ||
| 2010s | 5.3 a | 11.8 a | 20.1 a | 6.5 a | 8.5 a | 0.491 a | ||
| Year | Treatment | NSC Content in the Stem (g m−2) | NSC Remobilization Reserve (%) | ||
|---|---|---|---|---|---|
| Heading | Maturity | ||||
| 2017 | Cultivation treatment | LCP | 309 a | 160 a | 48.2 b |
| RNID | 258 b | 115 b | 55.4 a | ||
| Varietal type | 1990s | 250 b | 119 b | 52.4 a | |
| 2000s | 281 b | 138 ab | 50.9 a | ||
| 2010s | 321 a | 157 a | 51.1 a | ||
| 2018 | Cultivation treatment | LCP | 311 a | 158 a | 49.2 b |
| RNID | 268 b | 121 b | 54.9 a | ||
| Varietal type | 1990s | 258 b | 122 b | 52.7 a | |
| 2000s | 291 b | 142 ab | 51.2 a | ||
| 2010s | 320 a | 155 a | 51.6 a | ||
| Year | Treatment | LAI (m2 m−2) | |||
|---|---|---|---|---|---|
| Jointing | Heading | Maturity | |||
| 2017 | Cultivation treatment | LCP | 4.1 a | 7.4 a | 2.5 a |
| RNID | 3.7 b | 7.2 a | 2.2 a | ||
| Varietal type | 1990s | 3.3 b | 6.5 c | 1.9 c | |
| 2000s | 4.1 a | 7.2 b | 2.4 b | ||
| 2010s | 4.4 a | 8.2 a | 2.8 a | ||
| 2018 | Cultivation treatment | LCP | 4.0 a | 7.5 a | 2.5 a |
| RNID | 3.6 a | 7.2 b | 2.2 a | ||
| Varietal type | 1990s | 3.1 b | 6.7 c | 1.8 c | |
| 2000s | 4.0 a | 7.3 b | 2.4 b | ||
| 2010s | 4.3 a | 8.2 a | 2.9 a | ||
| Year | Treatment | Total Nutrient Accumulation (kg ha−1) | NUEg (kg Grain kg−1) | PUEg (kg Grain kg−1) | KUEg (kg Grain kg−1) | |||
|---|---|---|---|---|---|---|---|---|
| N | P | K | ||||||
| 2017 | Cultivation treatment | LCP | 237 a | 60.8 a | 282 a | 46.8 b | 182 a | 39.4 a |
| RNID | 230 a | 59.3 a | 278 a | 47.7 a | 186 a | 39.5 a | ||
| Varietal type | 1990s | 227 b | 58.6 b | 272 b | 46.8 a | 181 a | 38.9 b | |
| 2000s | 233 b | 60.1 ab | 281 a | 47.4 a | 184 a | 39.3 b | ||
| 2010s | 242 a | 61.5 a | 287 a | 47.6 a | 188 a | 40.2 a | ||
| 2018 | Cultivation treatment | LCP | 236 a | 59.8 a | 285 a | 47.1 a | 186 a | 39.0 a |
| RNID | 227 b | 58.1 a | 280 a | 47.8 a | 187 a | 38.7 a | ||
| Varietal type | 1990s | 227 b | 58.1 a | 278 b | 46.8 b | 183 a | 38.1 b | |
| 2000s | 231 ab | 58.8 a | 282 ab | 47.4 ab | 187 a | 38.8 b | ||
| 2010s | 238 a | 60.1 a | 289 a | 48.2 a | 191 a | 39.8 a | ||
| The Determined Parameters | Grain Yield | |
|---|---|---|
| LCP | RNID | |
| Spikelets per panicle | 0.86 ** | 0.78 ** |
| Shoot biomass weight at maturity | 0.90 ** | 0.93 ** |
| Harvest index | 0.73 ** | 0.81 ** |
| Shoot biomass accumulation from heading to maturity | 0.64 ** | 0.68 ** |
| NSC remobilization reserve | −0.80 ** | 0.72 ** |
| LAI at maturity | 0.69 ** | 0.84 ** |
| The SPAD of top three leaves at 24 DAH | 0.76 ** | 0.74 ** |
| The SPAD of top three leaves at 36 DAH | 0.68 ** | 0.85 ** |
| The photosynthetic rate of flag leaf at 30 DAH | 0.77 ** | 0.91 ** |
| The photosynthetic rate of flag leaf at 45 DAH | 0.81 ** | 0.88 ** |
| Total N accumulation | 0.88 ** | 0.94 ** |
| Total P accumulation | 0.52 * | 0.57 * |
| Total K accumulation | 0.89 ** | 0.76 ** |
| NUEg | 0.59 * | 0.71 ** |
| PUEg | 0.52 * | 0.78 ** |
| KUEg | 0.55 * | 0.84 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meng, T.; Chen, X.; Ge, J.; Zhang, X.; Zhou, G.; Dai, Q.; Wei, H. Reduced Nitrogen Rate with Increased Planting Density Facilitated Grain Yield and Nitrogen Use Efficiency in Modern Conventional Japonica Rice. Agriculture 2021, 11, 1188. https://doi.org/10.3390/agriculture11121188
Meng T, Chen X, Ge J, Zhang X, Zhou G, Dai Q, Wei H. Reduced Nitrogen Rate with Increased Planting Density Facilitated Grain Yield and Nitrogen Use Efficiency in Modern Conventional Japonica Rice. Agriculture. 2021; 11(12):1188. https://doi.org/10.3390/agriculture11121188
Chicago/Turabian StyleMeng, Tianyao, Xi Chen, Jialin Ge, Xubin Zhang, Guisheng Zhou, Qigen Dai, and Huanhe Wei. 2021. "Reduced Nitrogen Rate with Increased Planting Density Facilitated Grain Yield and Nitrogen Use Efficiency in Modern Conventional Japonica Rice" Agriculture 11, no. 12: 1188. https://doi.org/10.3390/agriculture11121188
APA StyleMeng, T., Chen, X., Ge, J., Zhang, X., Zhou, G., Dai, Q., & Wei, H. (2021). Reduced Nitrogen Rate with Increased Planting Density Facilitated Grain Yield and Nitrogen Use Efficiency in Modern Conventional Japonica Rice. Agriculture, 11(12), 1188. https://doi.org/10.3390/agriculture11121188