
Estimation of Dry Matter Production and Yield Prediction in Greenhouse Cucumber without Destructive Measurements

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
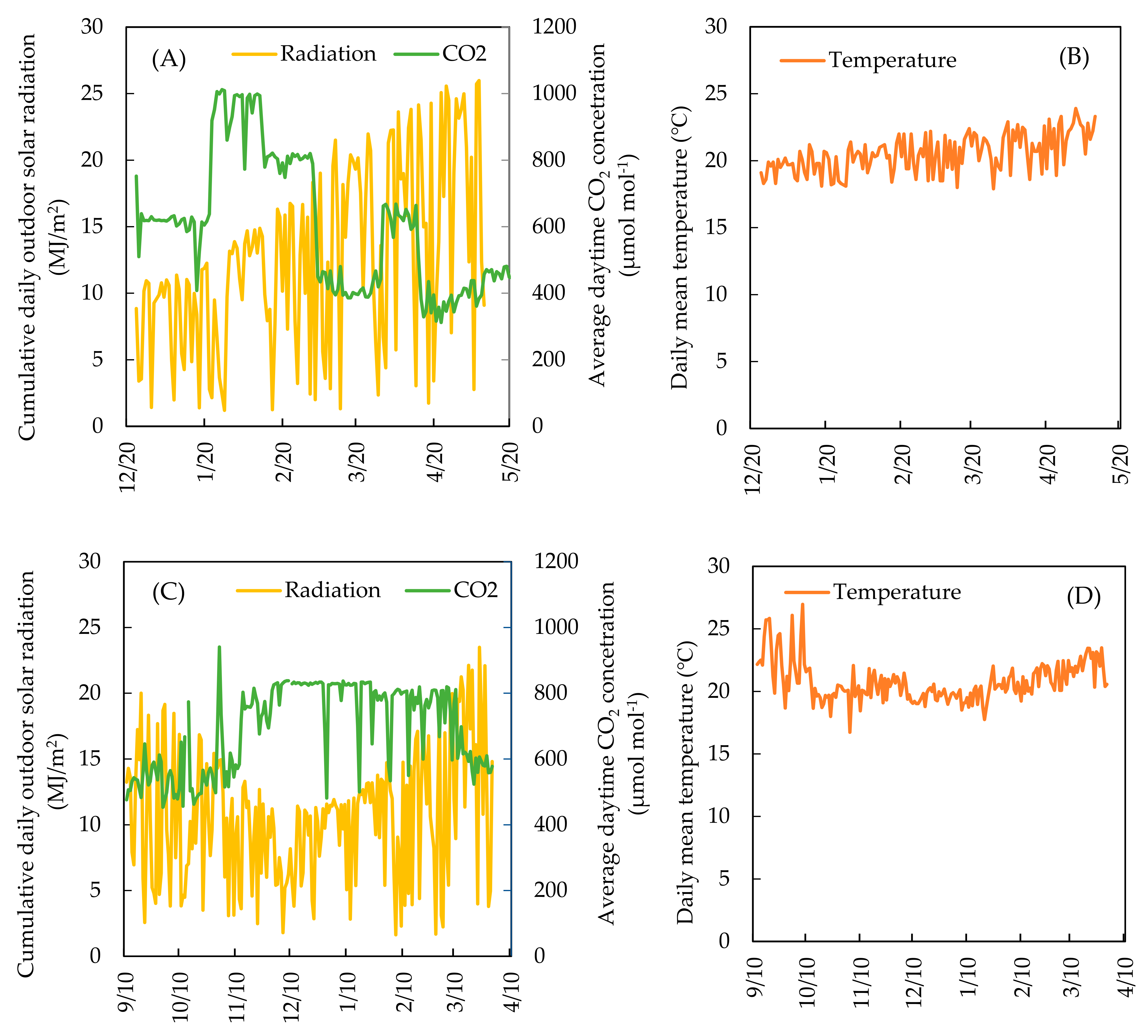
2.1. Plant Material and Growth Conditions
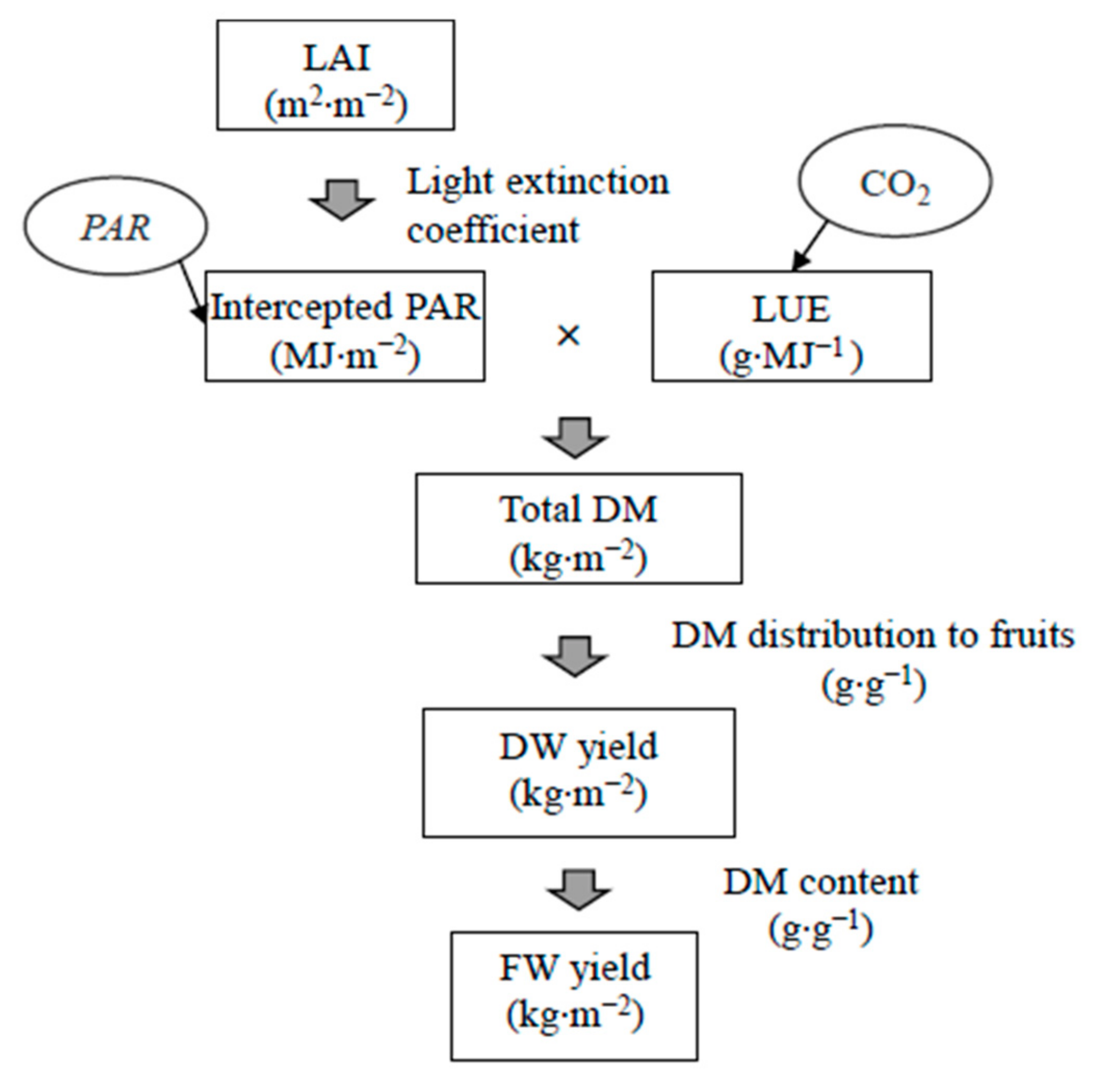
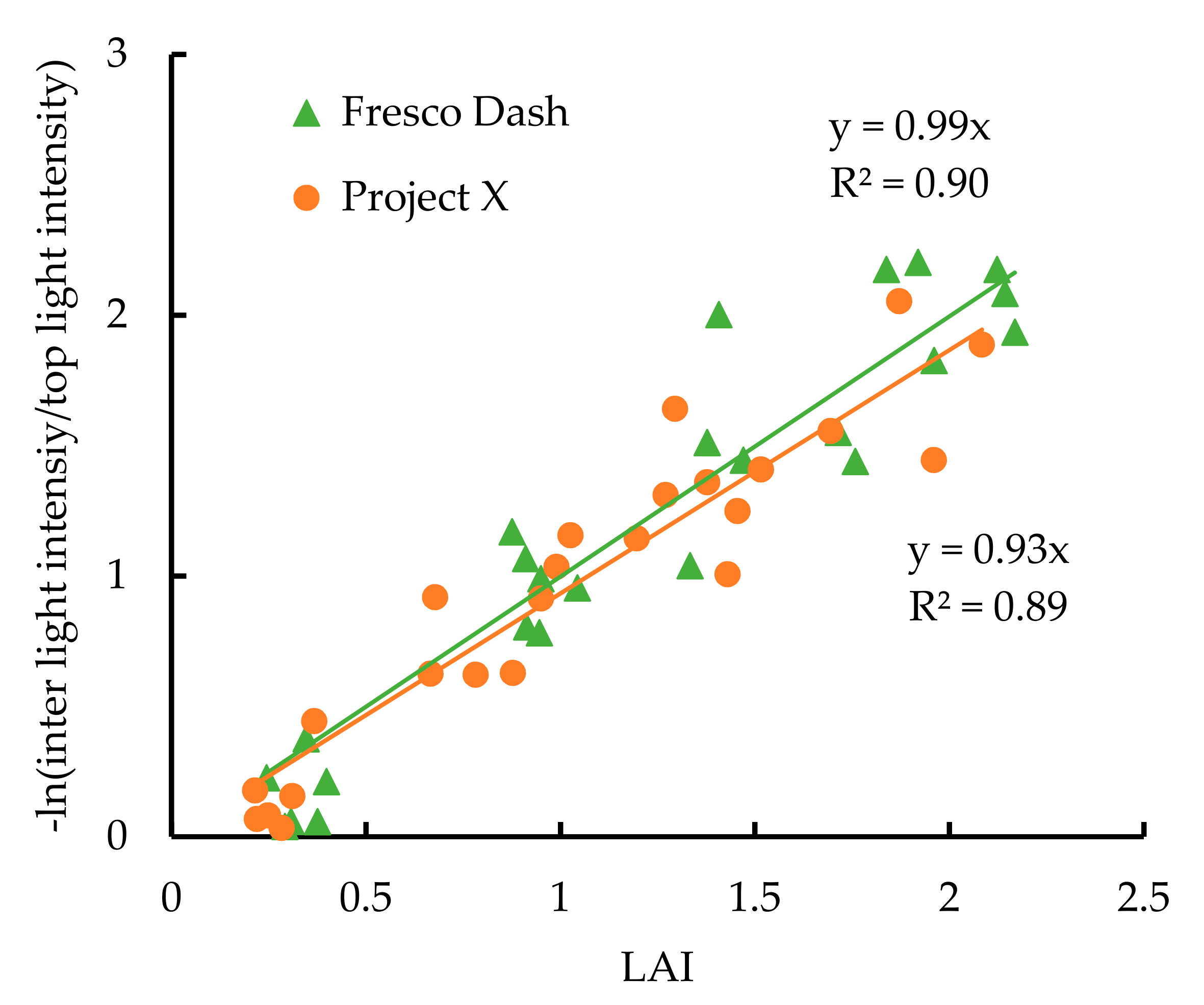
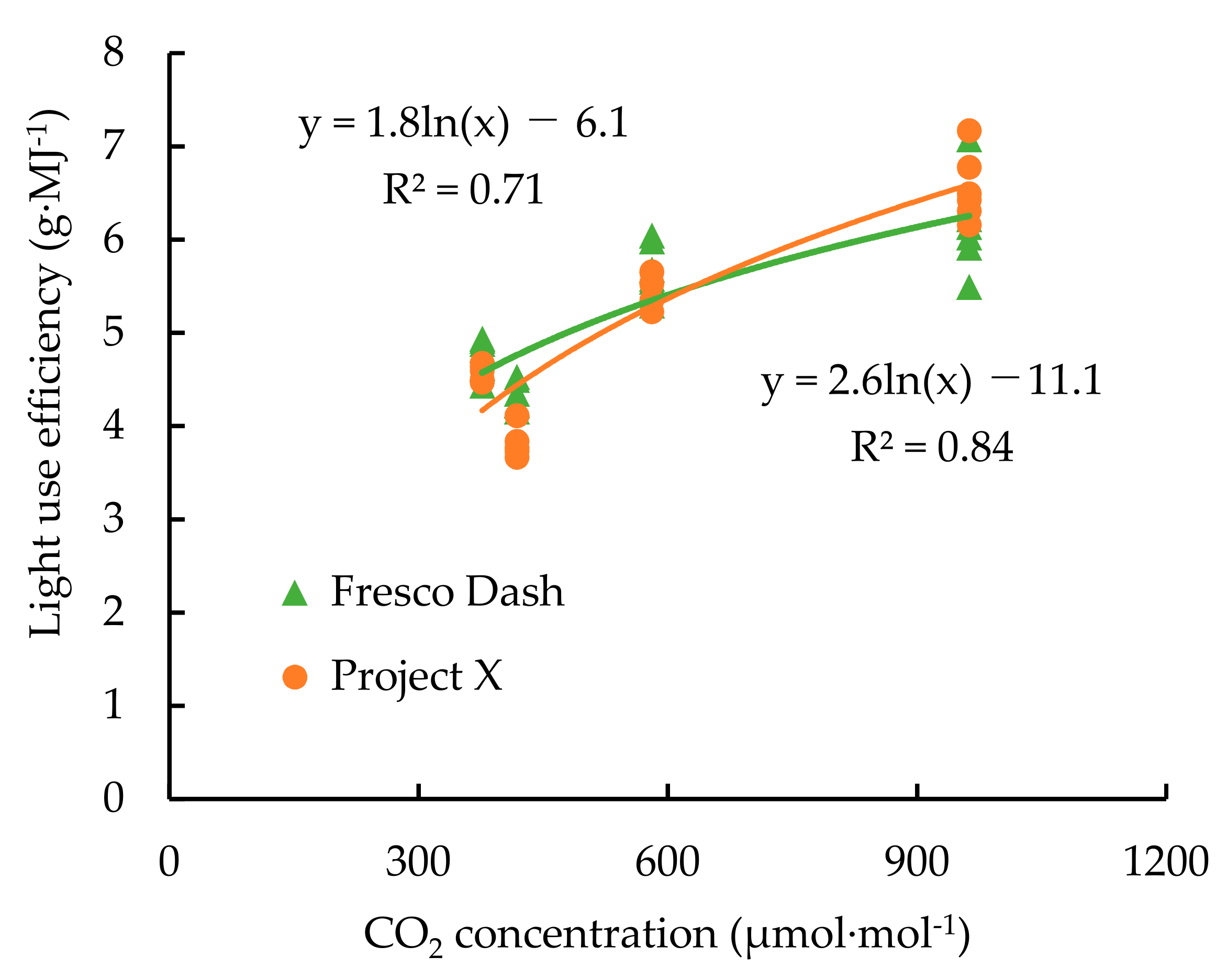
2.2. Empirical Growth Model and Plant Growth Measurement
2.3. Model Validation
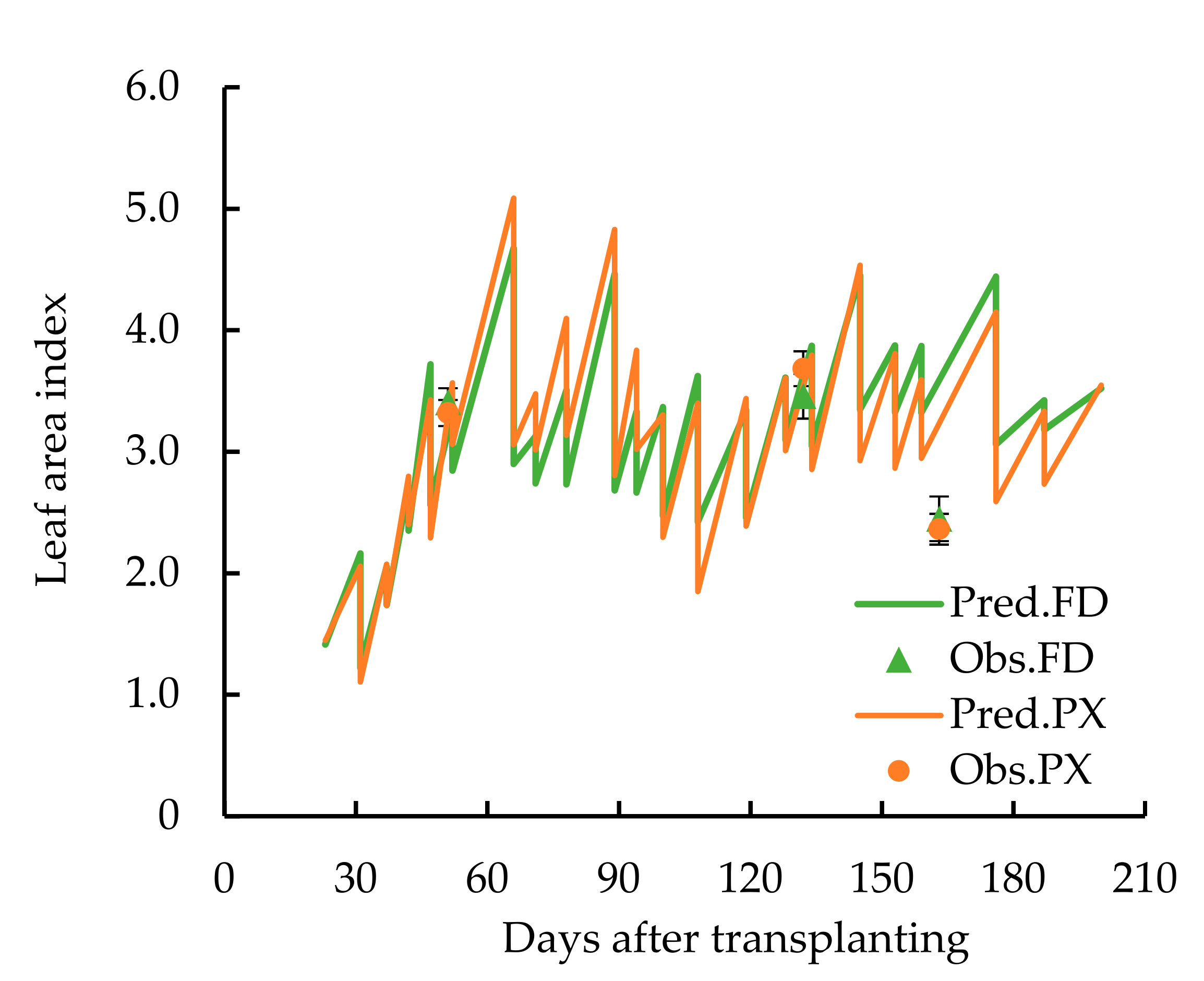
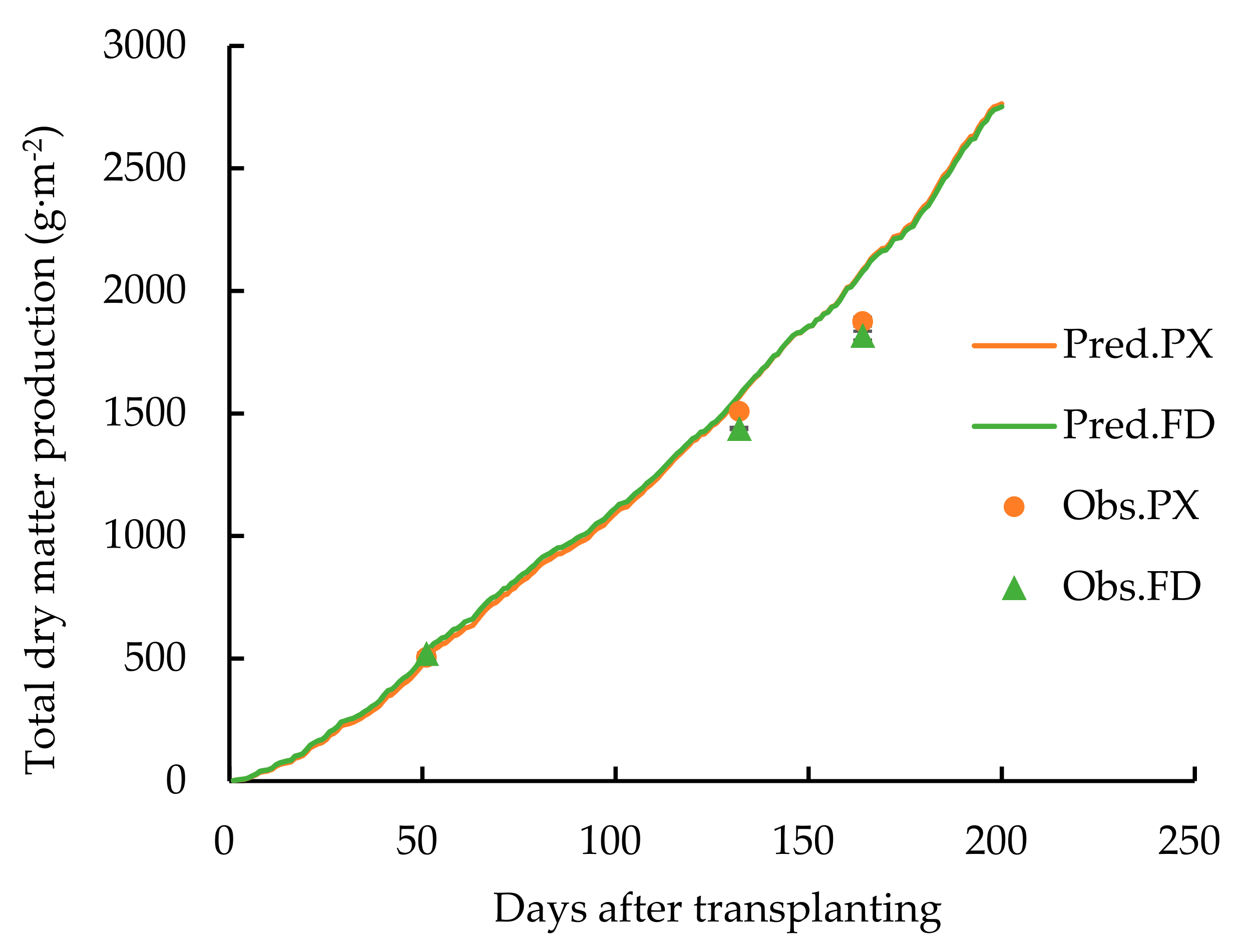
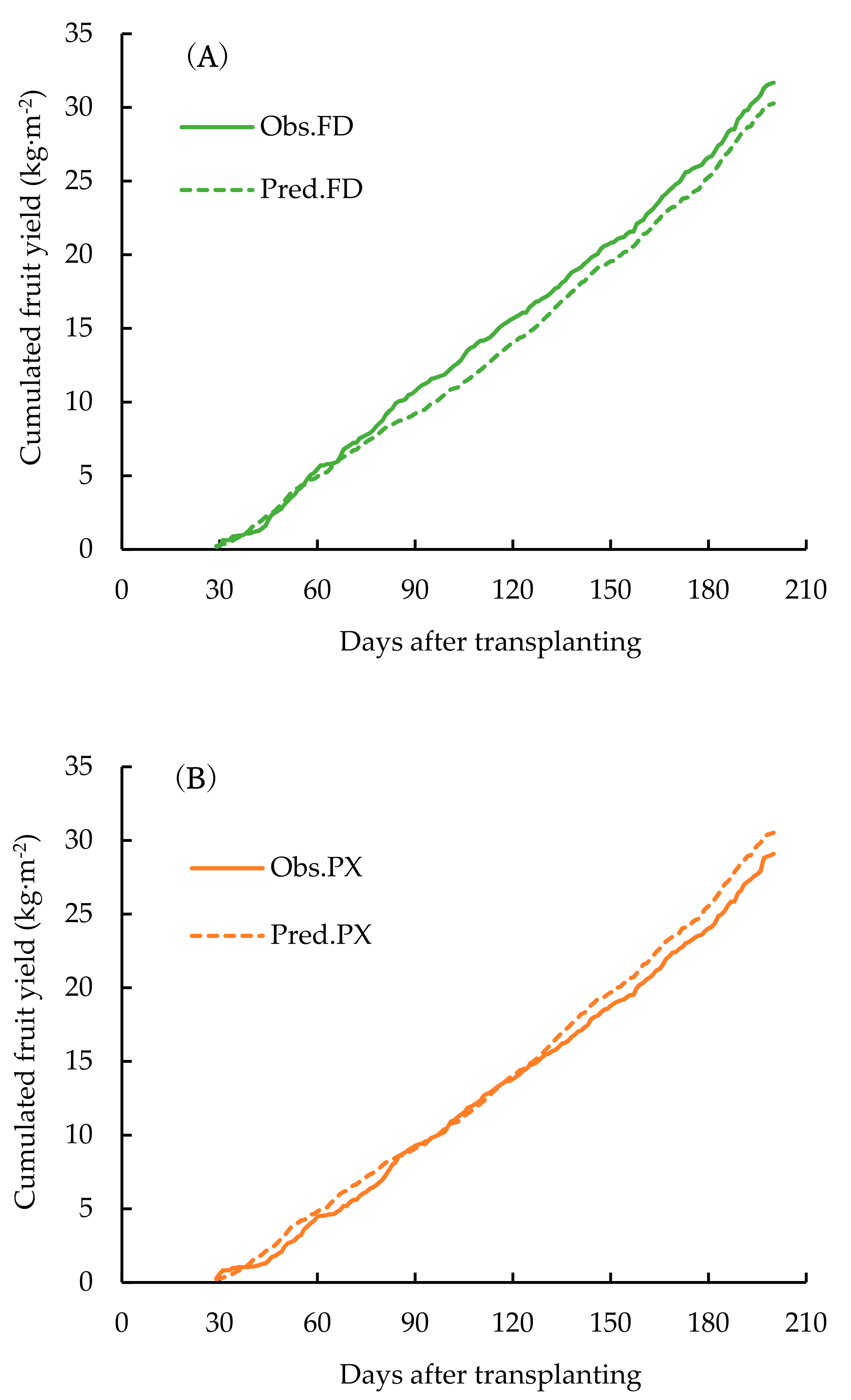
3. Result
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- MAFF. 2020. Available online: https://www.maff.go.jp/j/seisan/ryutu/engei/NextGenerationHorticulture/ (accessed on 12 March 2021).
- FAOSTAT. 2018. Available online: http://faostat (accessed on 14 May 2019).
- Van Ittersum, M.K.; Cassman, K.G.; Grassini, P.; Wolf, J.; Tittonell, P.; Hochman, Z. Yield gap analysis with local to global relevance—A review. Field Crops Res. 2013, 143, 4–17. [Google Scholar] [CrossRef] [Green Version]
- Lin, D.; Wei, R.; Xu, L. An Integrated Yield Prediction Model for Greenhouse Tomato. Agronomy 2019, 9, 873. [Google Scholar] [CrossRef] [Green Version]
- Dayan, E.; Vankeulen, H.; Jones, J.W.; Zipori, I.; Shmuel, D.; Challa, H. Development, Calibration and Validation of a Greenhouse Tomato Growth-Model.1. Description of the Model. Agr. Syst. 1993, 43, 145–163. [Google Scholar] [CrossRef]
- Vanthoor, B.; de Visser, P.; Stanghellini, C.; van Henten, E. A methodology for model-based greenhouse design: Part 2, description and validation of a tomato yield model. Biosyst. Eng. 2011, 110, 378–395. [Google Scholar] [CrossRef]
- Lopez, A.K.; Benavides, M.A.; Gonzalez, M.S.; Maldonado, A.J.; Chinas, S.P.; Morelos, M.A. Artificial neural network modeling of greenhouse Tomato yield and aerial dry matter. Agriculture 2020, 10, 97. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.-L.; Chung, S.-C.; Fu, W.-L.; Huang, C.-C. Artificial intelligence approaches to predict growth, harvest day, and quality of lettuce (Lactuca sativa L.) in a IoT-enabled greenhouse system. Biosyst. Eng. 2021, 212, 77–105. [Google Scholar] [CrossRef]
- Wang, Y.; Xiao, R.; Yin, Y.; Liu, T. Prediction of Tomato Yield in Chinese-Style Solar Greenhouses Based on Wavelet Neural Networks and Genetic Algorithms. Information 2021, 12, 336. [Google Scholar] [CrossRef]
- Gong, L.; Yu, M.; Jiang, S.; Cutsuridis, V.; Pearson, S. Deep Learning Based Prediction on Greenhouse Crop Yield Combined TCN and RNN. Sensors 2021, 21, 4537. [Google Scholar] [CrossRef]
- Louarn, G.; Song, Y. Two decades of functional-structural plant modelling: Now addressing fundamental questions in systems biology and predictive ecology. Ann. Bot. 2020, 126, 501–509. [Google Scholar] [CrossRef]
- Wen, W.; Wang, Y.; Wu, S.; Liu, K.; Gu, S.; Guo, X. 3D phytomer-based geometric modelling method for plants—The case of maize. AoB Plants 2021, 13, plab055. [Google Scholar] [CrossRef]
- Pao, Y.-C.; Kahlen, K.; Chen, T.-W.; Wiechers, D.; Stützel, H. How does structure matter? Comparison of canopy photosynthesis using one- and three-dimensional light models: A case study using greenhouse cucumber canopies. Silico Plants 2021, diab031. [Google Scholar] [CrossRef]
- Van der Meer, M.; de Visser, P.H.B.; Heuvelink, E.; Marcelis, L.F.M. Row orientation affects the uniformity of light absorption, but hardly affects crop photosynthesis in hedgerow tomato crops. Silico Plants 2021, 3, diab025. [Google Scholar] [CrossRef]
- O’Sullivan, H.; Raumonen, P.; Kaitaniemi, P.; Perttunen, J.; Sievänen, R. Integrating terrestrial laser scanning with functional–structural plant models to investigate ecological and evolutionary processes of forest communities. Ann. Bot. 2021, 128, 663–684. [Google Scholar] [CrossRef] [PubMed]
- Marcelis, L.; Gijzen, H. Evaluation under commercial conditions of a model of prediction of the yield and quality of cucumber fruits. Sci. Hortic. 1998, 76, 171–181. [Google Scholar] [CrossRef]
- Saito, T.; Kawasaki, Y.; Ahn, D.-H.; Ohyama, A.; Higashide, T. Prediction and Improvement of Yield and Dry Matter Production Based on Modeling and Non-destructive Measurement in Year-round Greenhouse Tomatoes. Hortic. J. 2020, 89, 425–431. [Google Scholar] [CrossRef] [Green Version]
- Saito, T.; Mochizuki, Y.; Kawasaki, Y.; Ohyama, A.; Higashide, T. Estimation of Leaf Area and Light-use Efficiency by Non-destructive Measurements for Growth Modeling and Recommended Leaf Area Index in Greenhouse Tomatoes. Hortic. J. 2020, 89, 445–453. [Google Scholar] [CrossRef]
- Higashide, T.; Heuvelink, E. Physiological and Morphological Changes Over the Past 50 Years in Yield Components in Tomato. J. Am. Soc. Hortic. Sci. 2009, 134, 460–465. [Google Scholar] [CrossRef] [Green Version]
- Kaneko, S.; Higashide, T.; Yasuba, K.-I.; Ohmori, H.; Nakano, A. Effects of Planting Stage and Density of Tomato Seedlings on Growth and Yield Component in Low-truss Cultivation. Hortic. Res. 2015, 14, 163–170. [Google Scholar] [CrossRef] [Green Version]
- Maeda, K.; Ahn, D.-H. A Review of Japanese Greenhouse Cucumber Research from the Perspective of Yield Components. Hortic. J. 2021, 90, 263–269. [Google Scholar] [CrossRef]
- Cho, Y.Y.; Oh, S.; Oh, M.M.; Son, J.E. Estimation of individual leaf area, fresh weight, and dry weight of hydroponically grown cucumbers (Cucumis sativus L.) using leaf length, width, and SPAD value. Sci. Hortic. 2007, 111, 330–334. [Google Scholar] [CrossRef]
- Monsi, M.; Saeki, T. On the factor light in plant communities and its importance for matter production. Ann. Bot. 2005, 95, 549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohtani, Y. Effective radiation, micrometeorological phenomena. In Agricultural Meteorology Glossary; Society for Agricultural Meteorology of Japan: Tokyo, Japan, 1997; pp. 106–107. (In Japanese) [Google Scholar]
- Ahn, D.; Higashide, T.; Iwasaki, Y.; Kawasaki, Y.; Nakano, A. Estimation of leaf area index of cucumbers (Cucumis sαtivus L.) trained on a high -wir. Bull. Natl. Inst. Veg. Tea Sci. 2015, 14, 23–29. [Google Scholar]
- Nederhoff, E.M. Effects of CO2 Concentration on Photosynthesis, Transpiration and Production of Greenhouse Fruit Vegetable Crops; Wageningen University and Research: Wageningen, The Netherlands, 1994. [Google Scholar]
- Kleinendorst, A.; Veen, B. Responses of young cucumber plants to root and shoot temperatures. Neth. J. Agric. Sci. 1983, 31, 47–61. [Google Scholar] [CrossRef]
- Marcelis, L. Effects of Sink Demand on Photosynthesis in Cucumber. J. Exp. Bot. 1991, 42, 1387–1392. [Google Scholar] [CrossRef]
- Alsadon, A.A.; Al-Helal, I.M.; Ibrahim, A.A.; Abdel-Ghany, A.M.; Al-Zaharani, S.M.; Gulrez, S.K.H. Growth response of cucumber under greenhouses covered with plastic films. J. Anim. Plant Sci. 2016, 26, 139–148. [Google Scholar]
- Heuvelink, E.; Kierkels, T. Plant Physiology in Greenhouses; Horti-Text: Woerden, The Netherlands, 2015. [Google Scholar]
- Shimomoto, K.; Takayama, K.; Takahashi, N.; Nishina, H.; Inaba, K.; Isoyama, Y.; Oh, S.-C. Real-time Monitoring of Photosynthesis and Transpiration of a Fully-grown Tomato Plant in Greenhouse. Environ. Control. Biol. 2020, 58, 65–70. [Google Scholar] [CrossRef]
- Wang, R.; Sun, Z.; Yang, D.; Ma, L. Simulating cucumber plant heights using optimized growth functions driven by water and accumulated temperature in a solar greenhouse. Agric. Water Manag. 2021, 259, 107170. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maeda, K.; Ahn, D.-H. Estimation of Dry Matter Production and Yield Prediction in Greenhouse Cucumber without Destructive Measurements. Agriculture 2021, 11, 1186. https://doi.org/10.3390/agriculture11121186
Maeda K, Ahn D-H. Estimation of Dry Matter Production and Yield Prediction in Greenhouse Cucumber without Destructive Measurements. Agriculture. 2021; 11(12):1186. https://doi.org/10.3390/agriculture11121186
Chicago/Turabian StyleMaeda, Kazuya, and Dong-Hyuk Ahn. 2021. "Estimation of Dry Matter Production and Yield Prediction in Greenhouse Cucumber without Destructive Measurements" Agriculture 11, no. 12: 1186. https://doi.org/10.3390/agriculture11121186
APA StyleMaeda, K., & Ahn, D.-H. (2021). Estimation of Dry Matter Production and Yield Prediction in Greenhouse Cucumber without Destructive Measurements. Agriculture, 11(12), 1186. https://doi.org/10.3390/agriculture11121186