Shoot Girdling Improves Rooting Performance of Kalamata Olive Cuttings by Upregulating Carbohydrates, Polyamines and Phenolic Compounds




Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Sampling
2.2. Biochemical Analyses
2.2.1. Carbohydrate Determination
2.2.2. Polyamine Determination
2.2.3. Extraction of Total Phenolic Compounds
Estimation of Endogenous Total Phenols
Estimation of Endogenous Total o-Diphenols
Estimation of Endogenous Total Flavonoids
Estimation of Endogenous Total Flavanols
Estimation of HPLC Phenolic Compounds
2.3. Statistical Analysis
3. Results
3.1. Effect of Girdling, Part of Shoot and Season on the Rooting Ability of Olive Cuttings
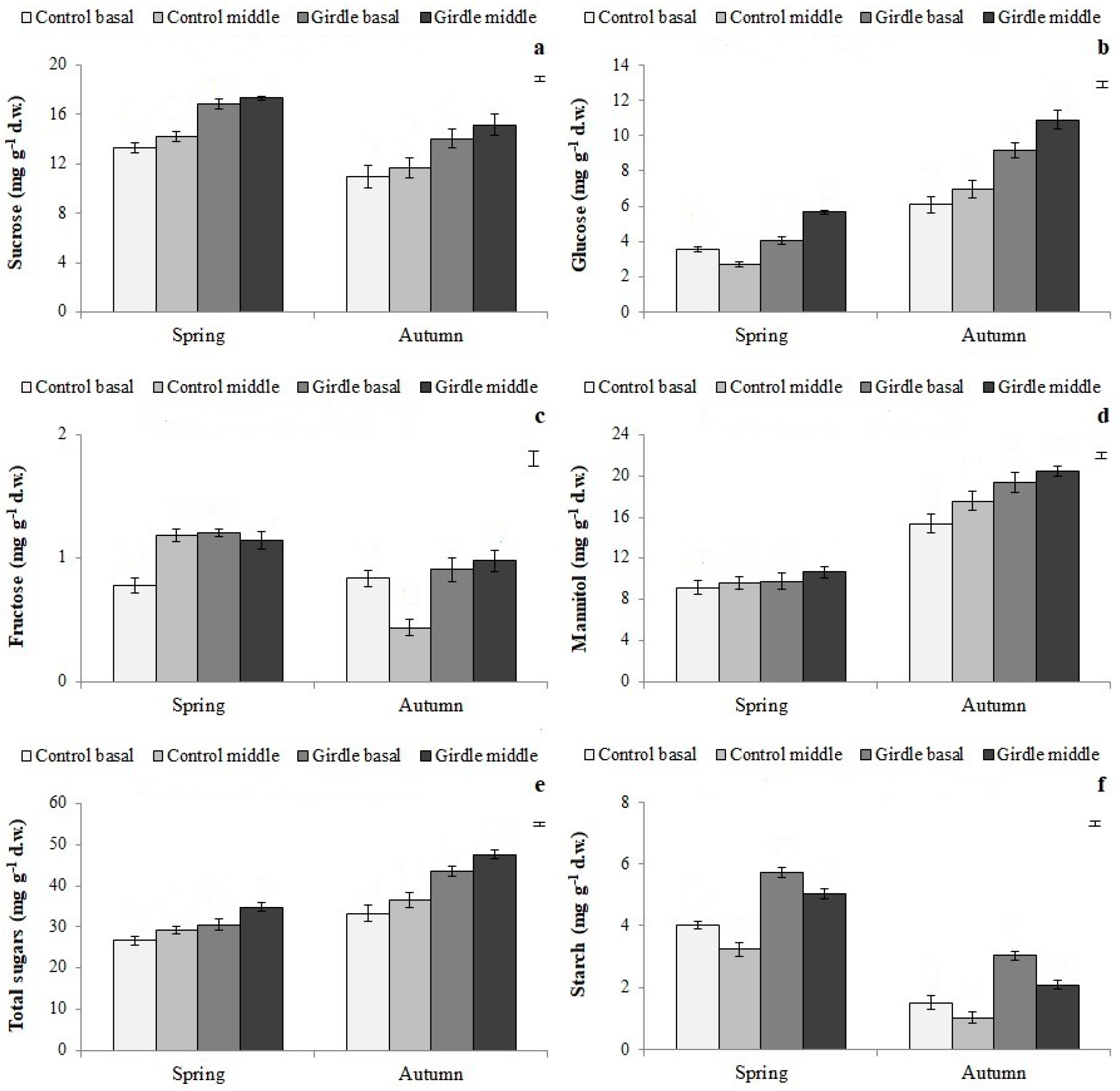
3.2. Carbohydrate Concentration in Cuttings
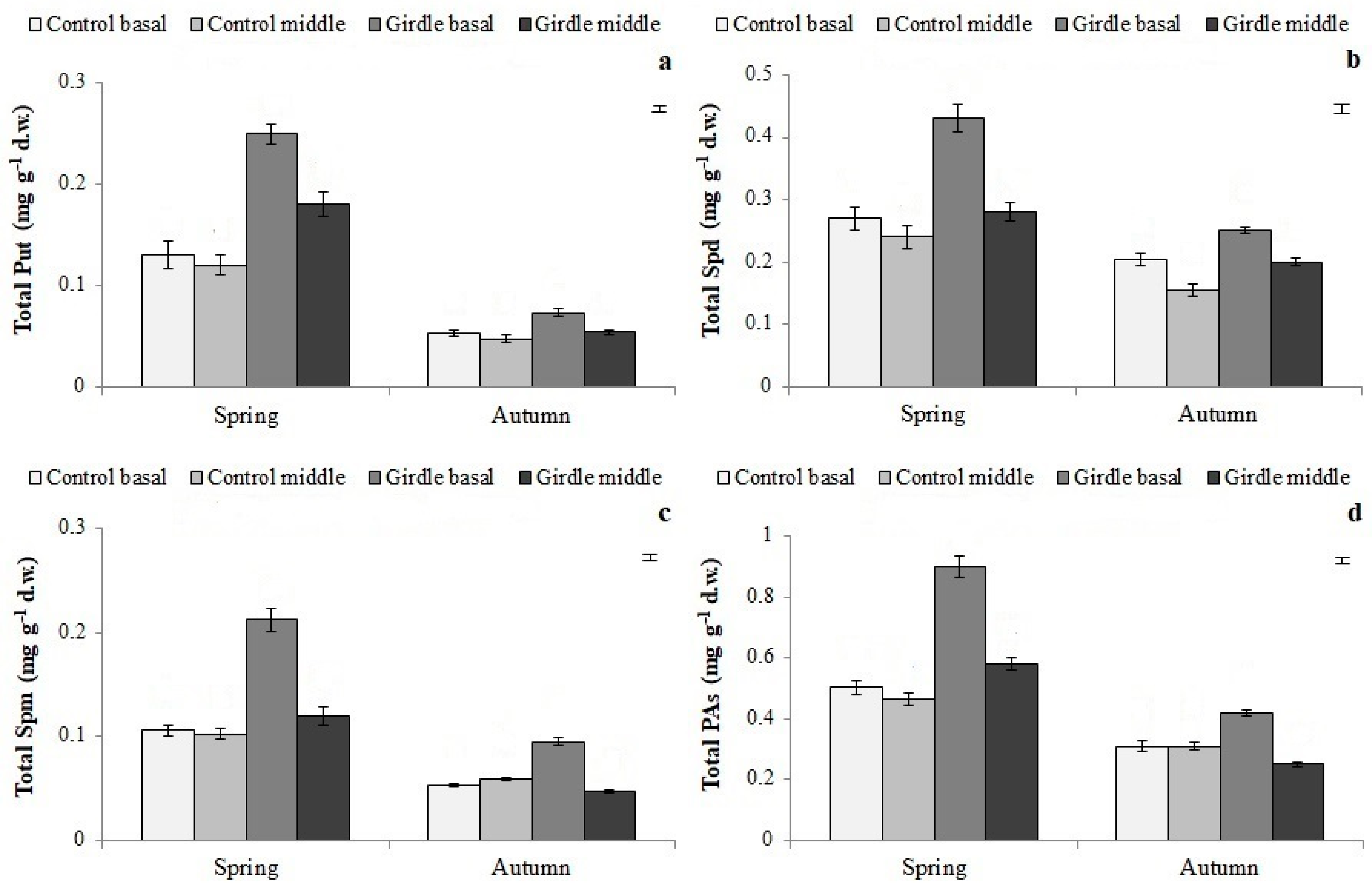
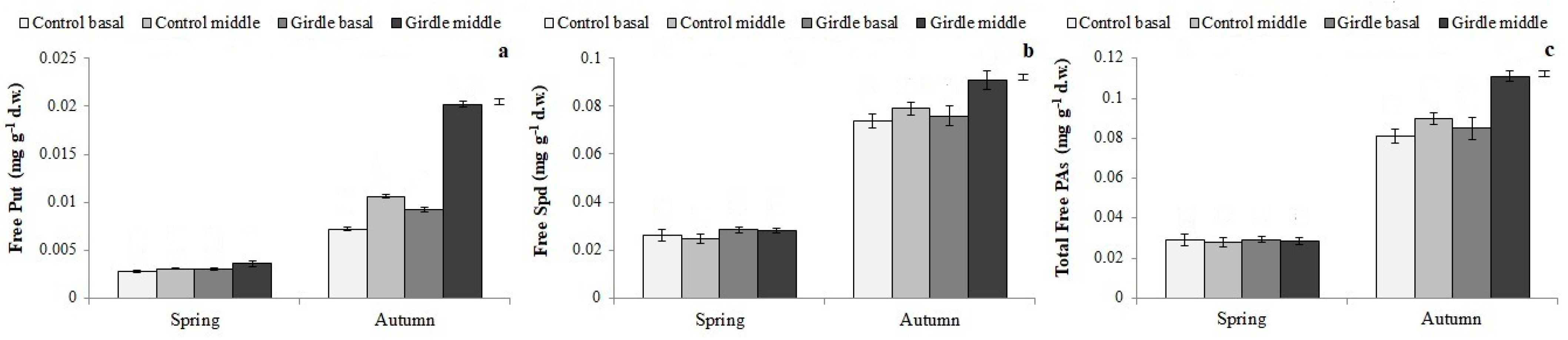
3.3. Polyamine Concentration in Cuttings
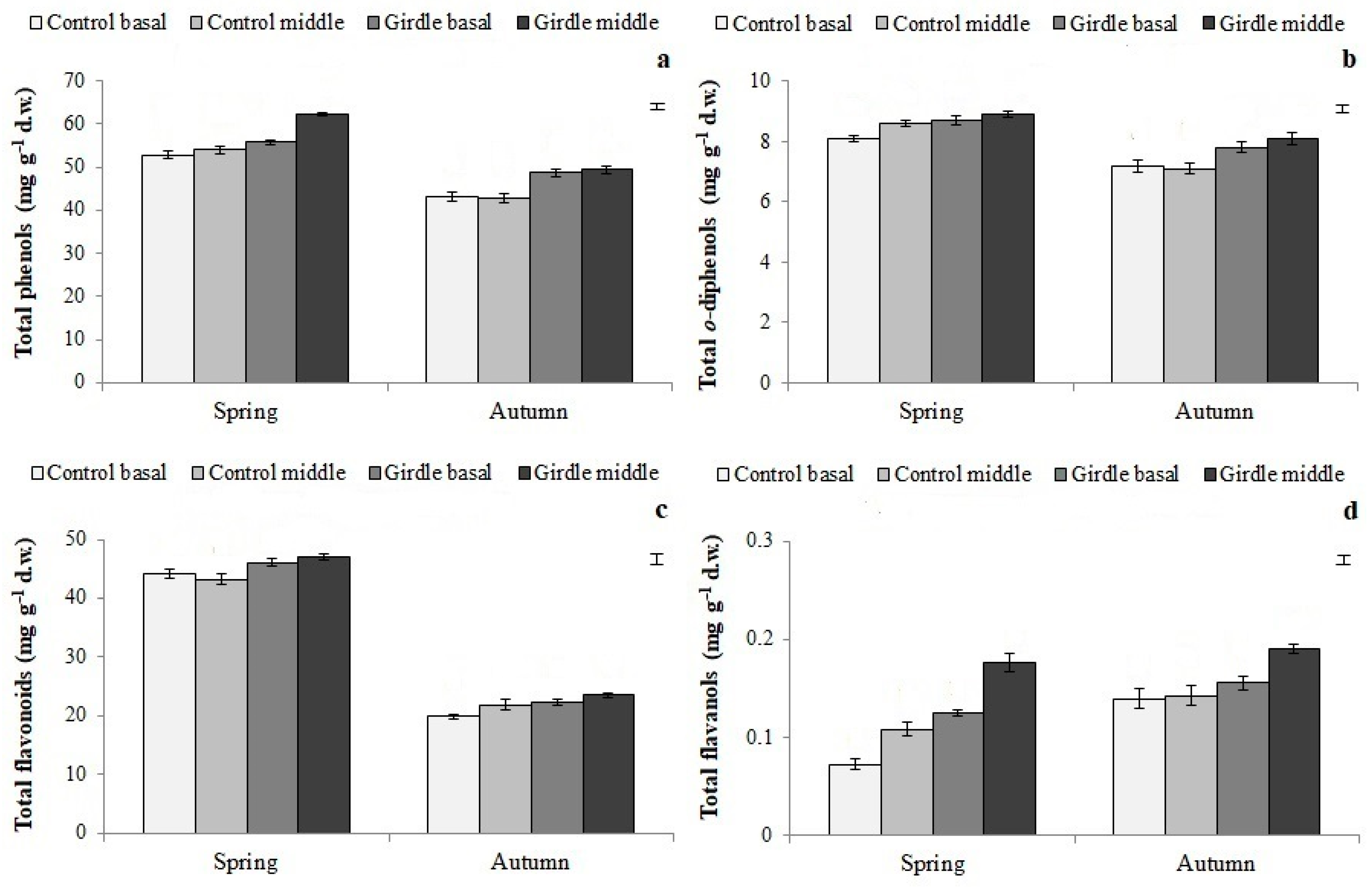
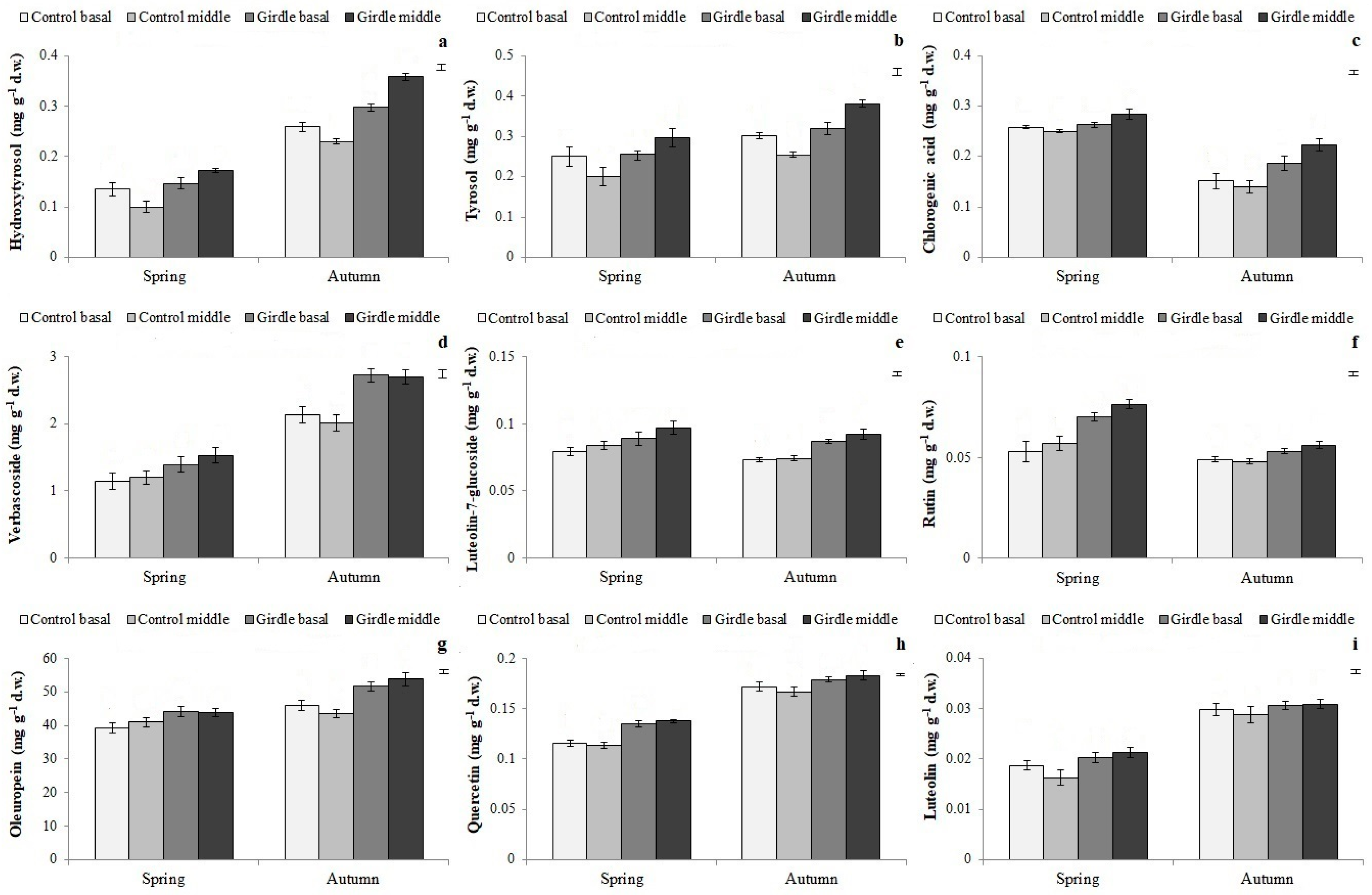
3.4. Phenolic Compound Concentration in Cuttings
3.5. Principal Component Analysis (PCA)
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fabbri, A.; Bartolini, G.; Lambardi, M.; Kailis, S.G. Olive Propagation Manual; Landlinks Press: Collingwood, Australia, 2004. [Google Scholar]
- Fontanazza, G. Olivicoltura Intensiva Meccanizzata; Edagricole: Bologna, Italy, 1993; p. 103. [Google Scholar]
- Wiesman, Z.; Lavee, S. Enhancement of stimulatory effects on rooting of olive cultivar stem cuttings. Sci. Hort. 1995, 62, 189–198. [Google Scholar] [CrossRef]
- Hartmann, H.T.; Kester, D.E.; Davies, F.T.; Geneve, R.L. Plant Propagation Principles and Practices, 7th ed.; Prentice Hall: Saddle River, NJ, USA, 2002. [Google Scholar]
- Sebastiani, L.; Tognetti, R. Growing season and hydrogen peroxide effects on root induction and development Olea europaea L. (cvs ‘Frantoio’ and ‘Gentile di Larino’) cuttings. Sci. Hort 2004, 100, 75–82. [Google Scholar] [CrossRef]
- Avidan, B.; Lavee, S. Physiological aspects of the rooting ability of olive cultivars. Acta Hort. 1978, 79, 93–101. [Google Scholar] [CrossRef]
- Sghir, S.; Belkoura, I.; Quazzani, N. Variability in the rooting ability of varieties of olive (Olea europaea L.). Olivae 2003, 96, 20–24. [Google Scholar]
- Sankhla, N.; Upadhyaya, A. Polyamines and adventitious root formation. In Adventitious Root Formation in Cuttings; Davis, T.D., Haissig, B.E., Sankhla, N., Eds.; Dioscorides Press: Portland, OR, USA, 1988; pp. 202–213. [Google Scholar]
- Cristofori, V.; Rouphael, Y.; Rugini, E. Collection time, cutting age, IBA and putrescine effects on root formation in Corylus avellana L. cuttings. Sci. Hort. 2010, 124, 189–194. [Google Scholar] [CrossRef]
- Denaxa, N.-K.; Vemmos, S.N.; Roussos, P.A.; Kostelenos, G. The effect of IBA, NAA and carbohydrates on rooting capacity of leafy cuttings in three olive cultivars (Olea europaea L.). Acta Hort. 2010, 924, 101–109. [Google Scholar] [CrossRef]
- Basak, U.C.; Das, A.B.; Das, P. Metabolic changes during rooting in stem cuttings of five mangrove species. Plant. Growth Regul. 1995, 17, 141–148. [Google Scholar] [CrossRef]
- Stoltz, L.; Hess, C. The effect of girdling upon root initiation: Carbohydrates and amino acids. Proc. Am. Soc. Hort. Sci. 1966, 89, 734–743. [Google Scholar]
- Goldschmidt, E.E.; Huber, S.C. Regulation of photosynthesis by end-product accumulation in leaves of plants storing starch, sucrose and hexose sugars. Plant. Physiol. 1992, 99, 1443–1448. [Google Scholar] [CrossRef]
- Aslmoshtaghi, E.; Shahsavar, A.R. Endogenous soluble sugars, starch contents and phenolic compounds in easy- and difficult-to-root olive cuttings. J. Biol. Environ. Sci. 2010, 49, 83–86. [Google Scholar]
- Vemmos, S.N.; Papagiannopoulou, A.; Coward, S. Effects of shoot girdling on photosynthetic capacity, leaf carbohydrate, and bud abscission in pistachio (Pistacia vera L.). Photosynthetica 2012, 50, 35–48. [Google Scholar] [CrossRef]
- Usta, S.S. The research on rooting ability of olive cuttings (Olea europaea L. cv. Domat). Acta Hort. 1999, 474, 63–66. [Google Scholar] [CrossRef]
- Wilson, P.J. Contributions of the stem to rooting in Eucalyptus grandis Hill ex Maid. stem cuttings. J. Hort. Sci. Biotechnol. 1994, 69, 1009–1017. [Google Scholar] [CrossRef]
- Negi, V.; Rana, S.; Hannah, P. Effect of auxins application on Rhododendron arboreum Smith pre-conditioned (girdled) stem cuttings. Environ. Ecol. 2017, 35, 3532–3534. [Google Scholar]
- Denaxa, N.-K.; Vemmos, S.N.; Roussos, P.A. The role of endogenous carbohydrates and seasonal variation in rooting ability of cuttings of an easy and a hard to root olive cultivars (Olea europaea L.). Sci. Hort. 2012, 143, 19–28. [Google Scholar] [CrossRef]
- Vemmos, S.N. Carbohydrate content of inflorescent buds of defruited and fruiting pistachio (Pistachia vera L) branches in relation to biennal bearing. J. Hort. Sci. Biotech. 1999, 74, 94–100. [Google Scholar] [CrossRef]
- Denaxa, N.-K.; Roussos, P.A.; Vemmos, S.N. The possible role of polyamines to the recalcitrance of ‘Kalamata’ olive leafy cuttings to root. J. Plant. Growth Regul. 2014, 33, 579–589. [Google Scholar] [CrossRef]
- Pedrol, N.; Tiburcio, A. Polyamines determination by TLC and HPLC. In Handbook of Plant Ecophysiology Techniques; Reigosa Roger, M.J., Ed.; Kluwer Academic Publishers: Amsterdam, The Netherlands, 2001; pp. 335–336. [Google Scholar]
- Roussos, P.; Denaxa, N.-K.; Damvakaris, T. Strawberry fruit quality attributes after application of plant growth stimulating compounds. Sci. Hort. 2009, 119, 138–146. [Google Scholar] [CrossRef]
- Denaxa, N.-K.; Roussos, P.A.; Vemmos, S.N. Assigning a role to the endogenous phenolic compounds on adventitious root formation of olive stem cuttings. J. Plant. Growth Regul. 2020, 39, 411–421. [Google Scholar] [CrossRef]
- Biswas, M. Girdling, wounding, stem maturity and IBA effects on rooting of jackfruit stem cuttings. J. Haw. Pac. Agrl. 1995, 6, 19–25. [Google Scholar]
- Delargy, J.A.; Wright, C.E. Root formation in cuttings of apple (cv. Bramley’s seedling) in relation to ringbarking and to etiolation. New Phytol. 1978, 81, 117–127. [Google Scholar] [CrossRef]
- Das, P.; Basak, U.C.; Das, A.B. Metabolic changes during rooting in pre-girdled stem cuttings and air-layers of Heritiera. Bot. Bull. Acad. Sin. 1997, 38, 91–95. [Google Scholar]
- Evert, D.R.; Smittle, D.A. Limb girdling influences rooting survival, total sugar and starch of dormant hardwood peach cuttings. HortScience 1990, 25, 1224–1226. [Google Scholar] [CrossRef]
- Wiesman, Z.; Lavee, S. Relationship of carbohydrate sources and indole-3-butyric acid in olive cuttings. Aust. J. Plant. Physiol. 1995, 22, 811–816. [Google Scholar] [CrossRef]
- Yoo, Y.K.; Kim, K.S. Seasonal variation in rooting ability, plant hormones, carbohydrate, nitrogen, starch and soluble sugar contents in cuttings of White Forsythia (Abeliophyllum distichum Nakai). J. Kor. Soc. Hort. Sci. 1996, 37, 554–560. [Google Scholar]
- Kuokkanen, K.; Julkunen-Tiitto, R.; Keinanen, M.; Niemela, P.; Tahvanainen, J. The effect of elevated CO2 and temperature on the secondary chemistry of Betula pendula seedlings. Trees-Struct. Funct. 2001, 15, 378–384. [Google Scholar] [CrossRef]
- Roussos, P.A.; Tassis, A. Effects of girdling, nitrogen, zinc and auxin foliar spray applications on mandarin fruit “Nova” quality characteristics. Emir. J. Food Agric. 2011, 23, 431–439. [Google Scholar]
- Kubota, N.; Nishiyama, N.; Shimamura, K. Effect of girdling lateral bearing branches on astringency and phenolic contents of peach fruits. J. Jpn. Soc. Hort. Sci. 1993, 62, 69–73. [Google Scholar] [CrossRef][Green Version]
- Khandaker, M.M.; Hossain, A.S.; Osman, N.; Boyce, A.N. Application of girdling for improved fruit retention, yield and fruit quality in Syzygium samarangense under field conditions. Int. J. Agric. Biol. 2011, 13, 18–24. [Google Scholar]
- Curir, P.; Sulis, S.; Mariani, F.; van Sumere, C.F.; Marchesini, A.; Dolci, M. Influence of endogenous phenols on rootability of Chamaelaucium uncinatum Schauer stem cuttings. Sci. Hort. 1993, 55, 103–314. [Google Scholar] [CrossRef]
- Osterc, G.; Štefančič, M.; Solar, A.; Štampar, F. Phenolic content in cuttings of two clones of hybrid chestnut (Castanea crenata x Castanea sativa) in the first days after cutting severance. Acta Agric. Scand. B 2008, 58, 162–168. [Google Scholar]
- Kevers, C.; Hausman, J.F.; Gaspar, T. Involvement of putrescine and of its catabolic pathway in the induction of rooting of walnut shoots in vitro. In Biology of Root Formation and Development; Altman, A., Waisel, Y., Eds.; Plenum Press: New York, NY, USA, 1997; pp. 161–162. [Google Scholar]
- Uribe, M.E.; Canal, M.J.; Fraga, M.; Ferrando, A.; Rios, D.; Noceda, C.; Altabella, T.; Tiburcio, A.F.; Rodrıguez, R. Polyamines in herbaceous and woody plants. Curr. Top. Plant Biol. 2004, 5, 53–62. [Google Scholar]
- Biondi, S.; Diaz, T.; Iglesias, I.; Gamberini, G.; Bagni, N. Polyamines and ethylene in relation to adventitious root for- mation in Prunus avium. Physiol. Plant. 1990, 78, 474–483. [Google Scholar] [CrossRef]
- Neves, C.; Santos, H.; Vilas-Boas, L.; Amancio, S. Involvement of free and conjugated polyamines and free amino acids in the adventitious rooting of micropropagated cork oak and grapevine shoots. Plant. Physiol. Biochem. 2002, 40, 1071–1080. [Google Scholar] [CrossRef]
- Tang, W.; Newton, R.J. Polyamines promote root elongation and growth by increasing root cell division in regenerated Virginia pine (Pinus virginiana Mill.) plantlets. Plant. Cell Rep. 2005, 24, 581–589. [Google Scholar] [CrossRef] [PubMed]
- Mendes, A.F.S.; Cidade, L.C.; Otoni, W.C.; Soares-Filho, W.S.; Costa, M.G.C. Role of auxins, polyamines and ethylene in root formation and growth in sweet orange. Biol. Plant. 2011, 55, 375–378. [Google Scholar] [CrossRef]
- Rugini, E.; Jacoboni, A.; Luppino, M. Role of basal shoot darkening and exogenous putrescine treatments on in vitro rooting and on endogenous polyamine changes in difficult-to-root woody species. Sci. Hort. 1993, 53, 63–72. [Google Scholar] [CrossRef]
- Rugini, E.; Di Francesco, G.; Muganu, M.; Astolfi, S.; Caricato, G. The effects of polyamines and hydrogen peroxide on root formation in olive and the role of polyamines as an early marker for rooting ability. In Biology of Root Formation and Development; Altman, A., Waisel, Y., Eds.; Plenum Press: New York, NY, USA, 1997; pp. 65–73. [Google Scholar]
- Tsafouros, A.; Frantzeskaki, A.; Assimakopoulou, A.; Roussos, P.A. Spatial and temporal changes of mineral nutrients and carbohydrates in cuttings of four stone fruit rootstocks and their contribution to rooting potential. Sci. Hort. 2019, 253, 227–240. [Google Scholar] [CrossRef]
- Loreti, F.; Morini, S. Propagation techniques. In The Peach: Botany, Production and Uses; Layne, D.R., Bassi, D., Eds.; CAB International: Cambridge, UK, 2008; pp. 221–243. [Google Scholar]
- Swarts, A.; Matsiliza-Mlathi, B.; Kleynhans, R. Rooting and survival of Lobostemon fruticosus (L) H. Buek stem cuttings as affected by season, media and cutting position. S. Afr. J. Bot. 2018, 119, 80–85. [Google Scholar] [CrossRef]
- Del Rio, C.; Rallo, L.; Caballero, J.M. Effects of carbohydrate content on the seasonal rooting of vegetative and reproductive cuttings of olive. J. Hort. Sci. 1991, 66, 301–309. [Google Scholar] [CrossRef]
- Tate, H.; Page, T. Cutting propagation of Santalum austrocaledonicum: The effect of genotype, cutting source, cutting size, propagation medium, IBA and irradiance. New For. 2018, 49, 551–570. [Google Scholar] [CrossRef]
- Tombesi, S.; Palliotti, A.; Poni, S.; Farinelli, D. Influence of light and shoot development stage on leaf photosynthesis and carbohydrate status during the adventitious root formation in cuttings of Corylus avellana L. Front. Plant. Sci. 2015, 6, 973. [Google Scholar] [CrossRef] [PubMed]
- Trobec, M.; Štampar, F.; Veberič, R.; Osterc, G. Fluctuations of different endogenous phenolic compounds and cinnamic acid in the first days of the rooting process of cherry rootstock ‘GiSelA 5′ leafy cuttings. J. Plant. Physiol. 2005, 162, 589–597. [Google Scholar] [CrossRef] [PubMed]
- Hausman, J.F.; Keversa, C.; Gaspara, T. Auxin-polyamine interaction in the control of the rooting inductive phase of poplar shoots in vitro. Plant. Sci. 1995, 110, 63–71. [Google Scholar] [CrossRef]
- Bartolini, G.; Toponi, M.A.; Di Monte, G. Variations in free polyamines and their effect on adventitious rooting of peach cuttings. J. Hort. Sci. Biotechnol. 2008, 83, 120–124. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factors | Rotten Cuttings (%) | Callusing (%) | Rooted Cuttings (%) |
|---|---|---|---|
| Treatment | |||
| Control | 84 ± 8 a | 13 ± 5 b | 3 ± 2 b |
| Girdling | 65 ± 7 b | 25 ± 7 a | 11 ± 4 a |
| Season | |||
| Autumn | 67 ± 8 b | 24 ± 7 a | 9 ± 4 a |
| Spring | 82 ± 9 a | 13 ± 5 b | 5 ± 3 b |
| Shoot part | |||
| Basal | 80 ± 7 a | 16 ± 6 a | 5 ± 3 b |
| Middle | 71 ± 7 b | 21 ± 6 a | 9 ± 4 a |
| Factors | Rotten Cuttings (%) | Callusing (%) | Rooted Cuttings (%) |
|---|---|---|---|
| Season × Treatment | |||
| Autumn × Control | 79 ± 8 b | 17 ± 5 b | 4 ± 3 c |
| Autumn × Girdling | 55 ± 6 c | 32 ± 8 a | 13 ± 5 a |
| Spring × Control | 87 ± 9 a | 9 ± 4 c | 2 ± 2 c |
| Spring × Girdling | 74 ± 9 b | 18 ± 6 b | 8 ± 4 b |
| Season × Shoot part | |||
| Autumn × Basal cuttings | 71 ± 8 a | 22 ± 6 a | 7 ± 3 a |
| Autumn × Middle cuttings | 63 ± 7 a | 26 ± 8 a | 11 ± 5 a |
| Spring × Basal cuttings | 84 ± 8 a | 12 ± 5 a | 4 ± 3 a |
| Spring × Middle cuttings | 79 ± 10 a | 14 ± 6 a | 7 ± 3 a |
| Treatment × Shoot part | |||
| Control × Basal cuttings | 85 ± 8 a | 12 ± 5 a | 3 ± 2 c |
| Girdling × Basal cuttings | 70 ± 7 a | 22 ± 6 a | 8 ± 4 b |
| Control × Middle cuttings | 82 ± 9 a | 14 ± 4 a | 4 ± 3 c |
| Girdling × Middle cuttings | 59 ± 8 a | 27 ± 8 a | 14 ± 5 a |
| Season × Treatment × Shoot part | |||
| Spring × Control × Basal cuttings | 90 ± 8 a | 8 ± 5 c | 2 ± 2 c |
| Spring × Control × Middle cuttings | 87 ± 11 a | 10 ± 4 c | 3 ± 3 c |
| Spring × Girdling × Basal cuttings | 78 ± 9 ab | 17 ± 5 bc | 5 ± 4 c |
| Spring × Girdling × Middle cuttings | 70 ± 9 bc | 19 ± 7 bc | 11 ± 4 b |
| Autumn × Control × Basal cuttings | 80 ± 10 ab | 16 ± 6 bc | 4 ± 3 c |
| Autumn × Control × Middle cuttings | 77 ± 8 ab | 18 ± 5 bc | 5 ± 3 c |
| Autumn × Girdling × Basal cuttings | 62 ± 6 cd | 28 ± 8 ab | 10 ± 4 b |
| Autumn × Girdling × Middle cuttings | 49 ± 7 d | 35 ± 10 a | 16 ± 7 a |
| Factors | Suc | Gluc | Fruc | Man | Total Sugars | Starch | FPut | FSpd | FTotal | Put | Spd | Spm | Total |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Treatment (T) | *** | *** | ** | *** | *** | *** | *** | ** | *** | *** | *** | *** | *** |
| Shoot part (P) | * | *** | ns | ** | *** | *** | *** | ** | *** | *** | *** | *** | *** |
| Season (S) | *** | *** | ** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** |
| T × P | ns | *** | ns | ns | ns | ns | *** | ns | ** | *** | ** | *** | *** |
| T × S | ns | *** | ns | ** | *** | * | *** | ** | ** | *** | ** | *** | *** |
| P × S | ns | ns | * | ns | ns | ns | *** | ns | *** | ** | ns | ** | ** |
| T × P × S | ns | ns | ** | ns | ns | ns | *** | ns | * | * | ** | ** | *** |
| Factors | Total Phenols | Total o-Diphenols | Total Flavonoids | Total Flavanols |
|---|---|---|---|---|
| Treatment (T) | *** | *** | *** | *** |
| Shoot part (P) | *** | *** | ** | *** |
| Season (S) | *** | *** | *** | *** |
| T × P | *** | ns | ns | *** |
| T × S | ns | ** | ns | *** |
| P × S | *** | ns | *** | *** |
| T × P × S | ** | ** | ns | *** |
| Factors | HT | Tyr | CGA | Ver | Luteolin-7 | Rutin | OLE | Quer | Luteolin |
|---|---|---|---|---|---|---|---|---|---|
| Treatment (T) | *** | *** | *** | *** | *** | *** | *** | *** | ** |
| Shoot part (P) | ns | ns | ns | ns | ** | ** | ns | ns | ns |
| Season (S) | *** | *** | *** | *** | ** | *** | *** | *** | *** |
| T × P | *** | *** | *** | ns | ns | ns | ns | ** | ns |
| T × S | * | ns | *** | ns | ns | *** | ** | *** | ns |
| P × S | ns | ns | ns | ns | ns | ns | ns | ns | ns |
| T × P x S | ns | ns | ns | ns | ns | ns | * | ns | ns |
| Factors | Principal Components | |||
|---|---|---|---|---|
| 1 | 2 | 3 | 4 | |
| Rooting | 0.089 | 0.308 | −0.202 | −0.014 |
| Hydroxytyrosol | 0.227 | 0.142 | 0.002 | −0.082 |
| Tyrosol | 0.161 | 0.220 | −0.118 | −0.144 |
| Chlorogenic acid | −0.199 | 0.193 | −0.158 | 0.109 |
| Verbascoside | 0.221 | 0.149 | 0.063 | −0.034 |
| Luteolin-7-glucoside | −0.069 | 0.354 | −0.245 | 0.011 |
| Rutin | −0.158 | 0.295 | −0.175 | −0.262 |
| Oleuropein | 0.182 | 0.254 | 0.083 | 0.041 |
| Quercetin | 0.233 | 0.105 | 0.068 | −0.189 |
| Luteolin | 0.232 | 0.055 | 0.086 | −0.214 |
| Free Put | 0.209 | 0.145 | −0.058 | 0.172 |
| Free Spd | 0.246 | 0.029 | 0.058 | −0.026 |
| Total free PAs | 0.245 | 0.041 | 0.044 | 0.026 |
| Total Put | −0.212 | 0.152 | 0.143 | −0.262 |
| Total Spd | −0.174 | 0.169 | 0.394 | −0.273 |
| Total Spm | −0.190 | 0.168 | 0.353 | −0.218 |
| Total PAs | −0.196 | 0.167 | 0.305 | −0.262 |
| Sucrose | −0.127 | 0.241 | 0.230 | 0.429 |
| Glucose | 0.271 | 0.159 | 0.201 | 0.172 |
| Fructose | −0.099 | 0.191 | 0.001 | 0.267 |
| Mannitol | 0.229 | 0.085 | 0.134 | 0.083 |
| Total Sugars | 0.184 | 0.233 | 0.222 | 0.211 |
| Starch | −0.207 | 0.070 | 0.223 | 0.335 |
| Total phenols | −0.199 | 0.205 | −0.264 | 0.008 |
| Total o-diphenols | −0.163 | 0.262 | −0.117 | 0.166 |
| Total flavonoids | −0.243 | 0.059 | −0.088 | 0.051 |
| Total flavanols | 0.144 | 0.281 | −0.255 | −0.140 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Denaxa, N.-K.; Vemmos, S.N.; Roussos, P.A. Shoot Girdling Improves Rooting Performance of Kalamata Olive Cuttings by Upregulating Carbohydrates, Polyamines and Phenolic Compounds. Agriculture 2021, 11, 71. https://doi.org/10.3390/agriculture11010071
Denaxa N-K, Vemmos SN, Roussos PA. Shoot Girdling Improves Rooting Performance of Kalamata Olive Cuttings by Upregulating Carbohydrates, Polyamines and Phenolic Compounds. Agriculture. 2021; 11(1):71. https://doi.org/10.3390/agriculture11010071
Chicago/Turabian StyleDenaxa, Nikoleta-Kleio, Stavros N. Vemmos, and Peter A. Roussos. 2021. "Shoot Girdling Improves Rooting Performance of Kalamata Olive Cuttings by Upregulating Carbohydrates, Polyamines and Phenolic Compounds" Agriculture 11, no. 1: 71. https://doi.org/10.3390/agriculture11010071
APA StyleDenaxa, N.-K., Vemmos, S. N., & Roussos, P. A. (2021). Shoot Girdling Improves Rooting Performance of Kalamata Olive Cuttings by Upregulating Carbohydrates, Polyamines and Phenolic Compounds. Agriculture, 11(1), 71. https://doi.org/10.3390/agriculture11010071