Could the Content of Soluble Carbohydrates in the Young Shoots of Selected Willow Cultivars Be a Determinant of the Plants’ Attractiveness to Cervids (Cervidae, Mammalia)?

 , ,
, ,  ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
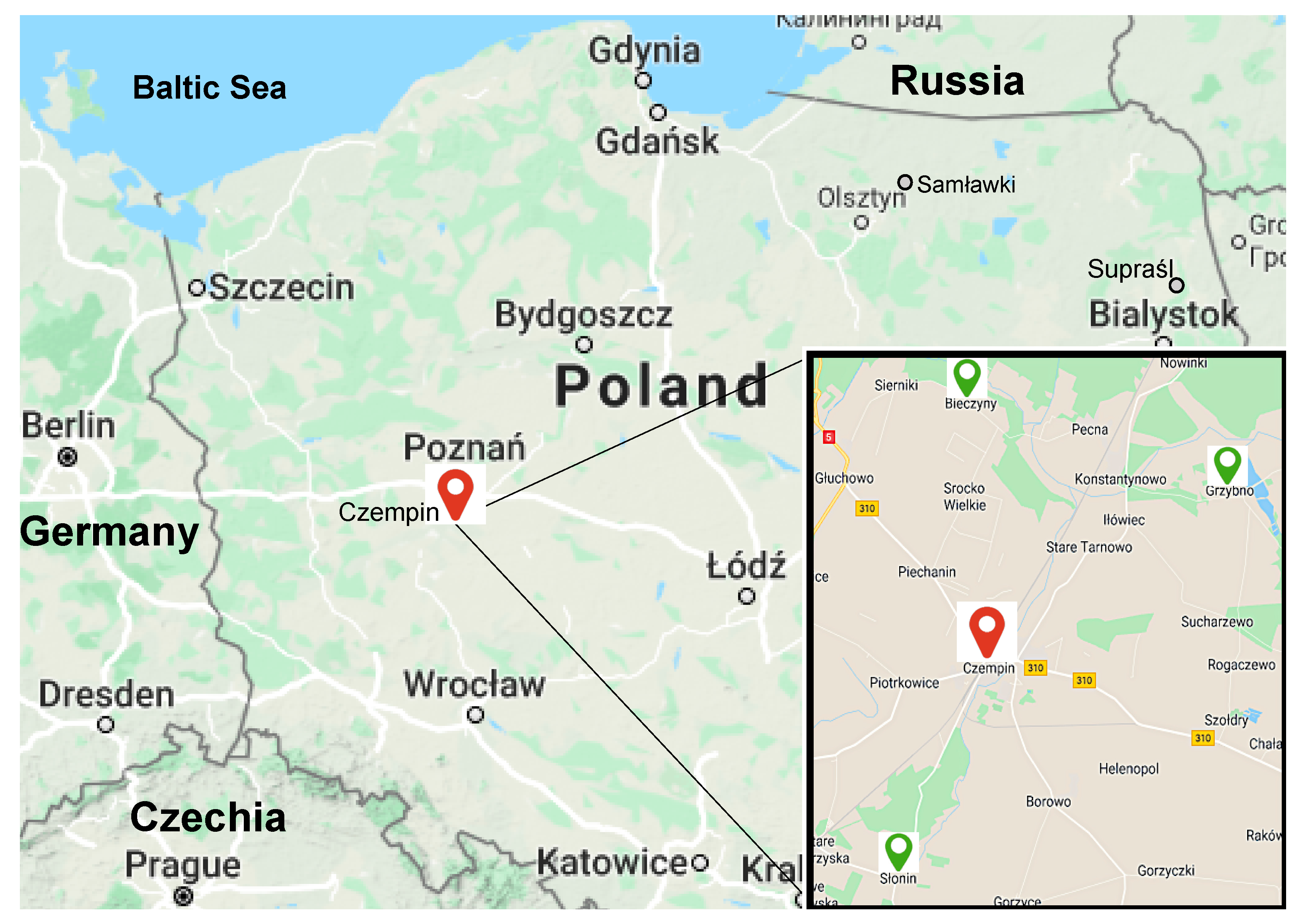
2.1. Experimental Plots
2.2. Browsing Damage Assessment
2.3. Analysis of the Content of Soluble Carbohydrates
2.4. Experiment with Farmed Red Deer and Fallow Deer
2.5. Statistical Analysis
3. Results
3.1. Soil Analysis
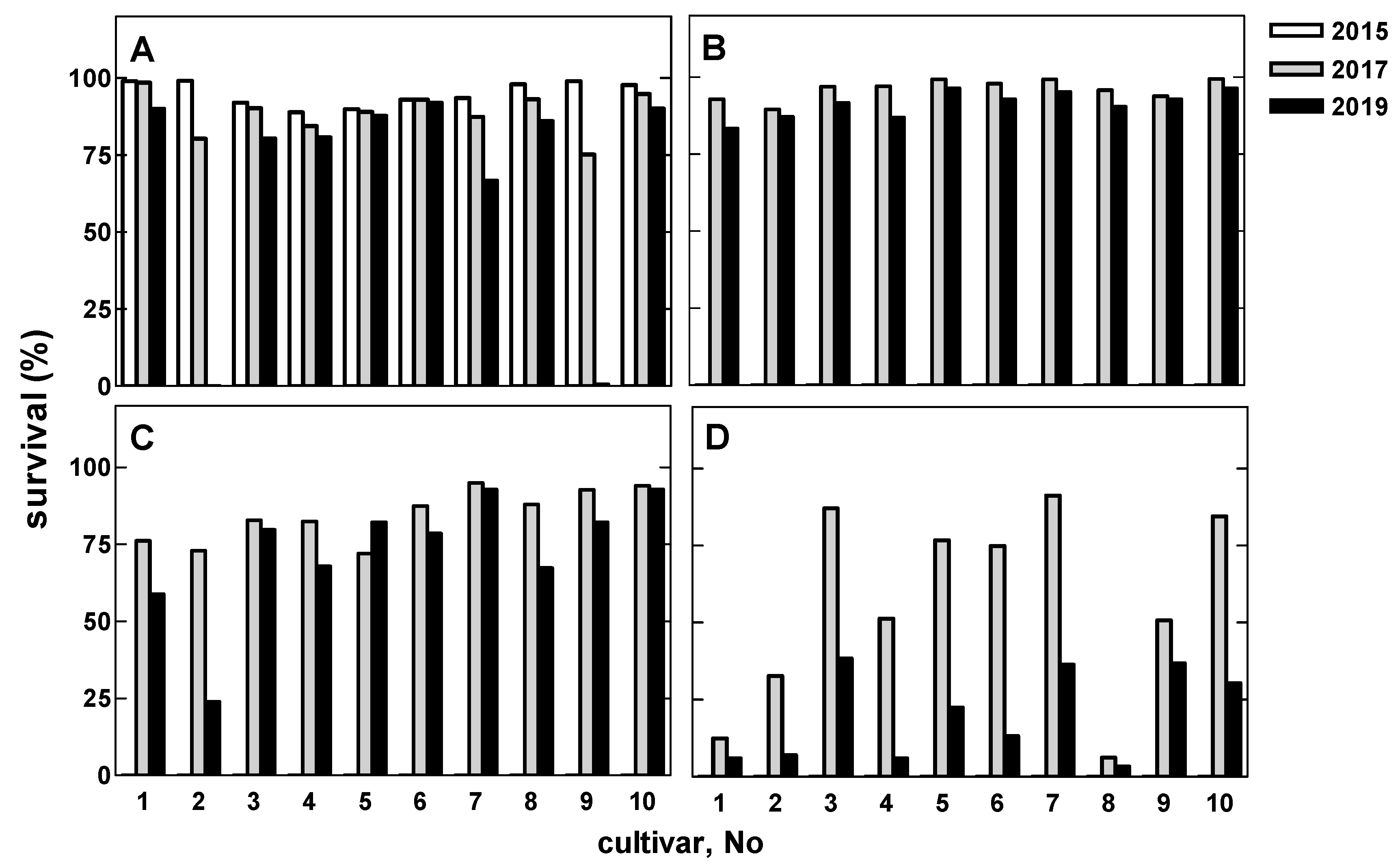
3.2. Willow Performance (Survival)
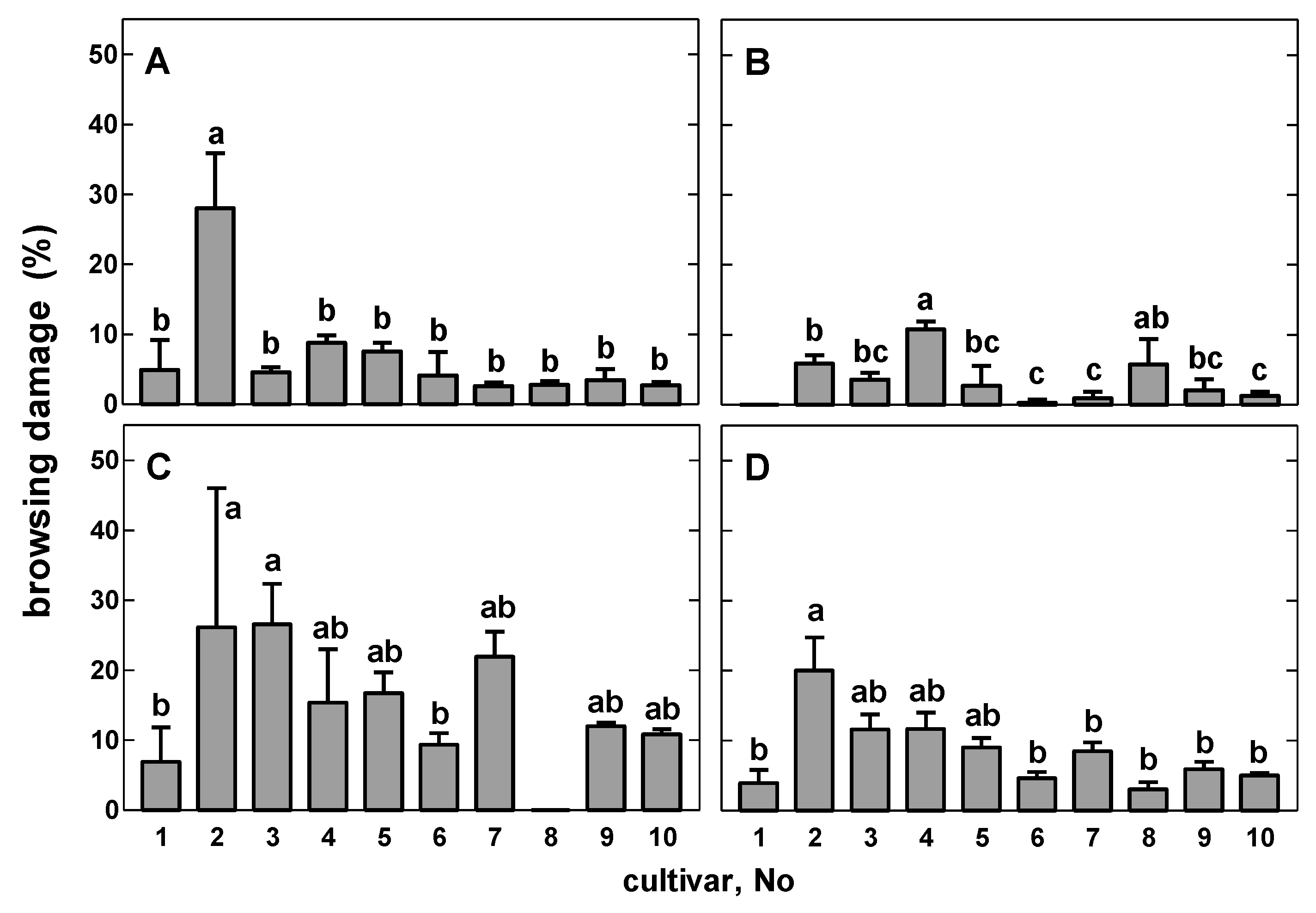
3.3. Analysis of Shoot Browsing
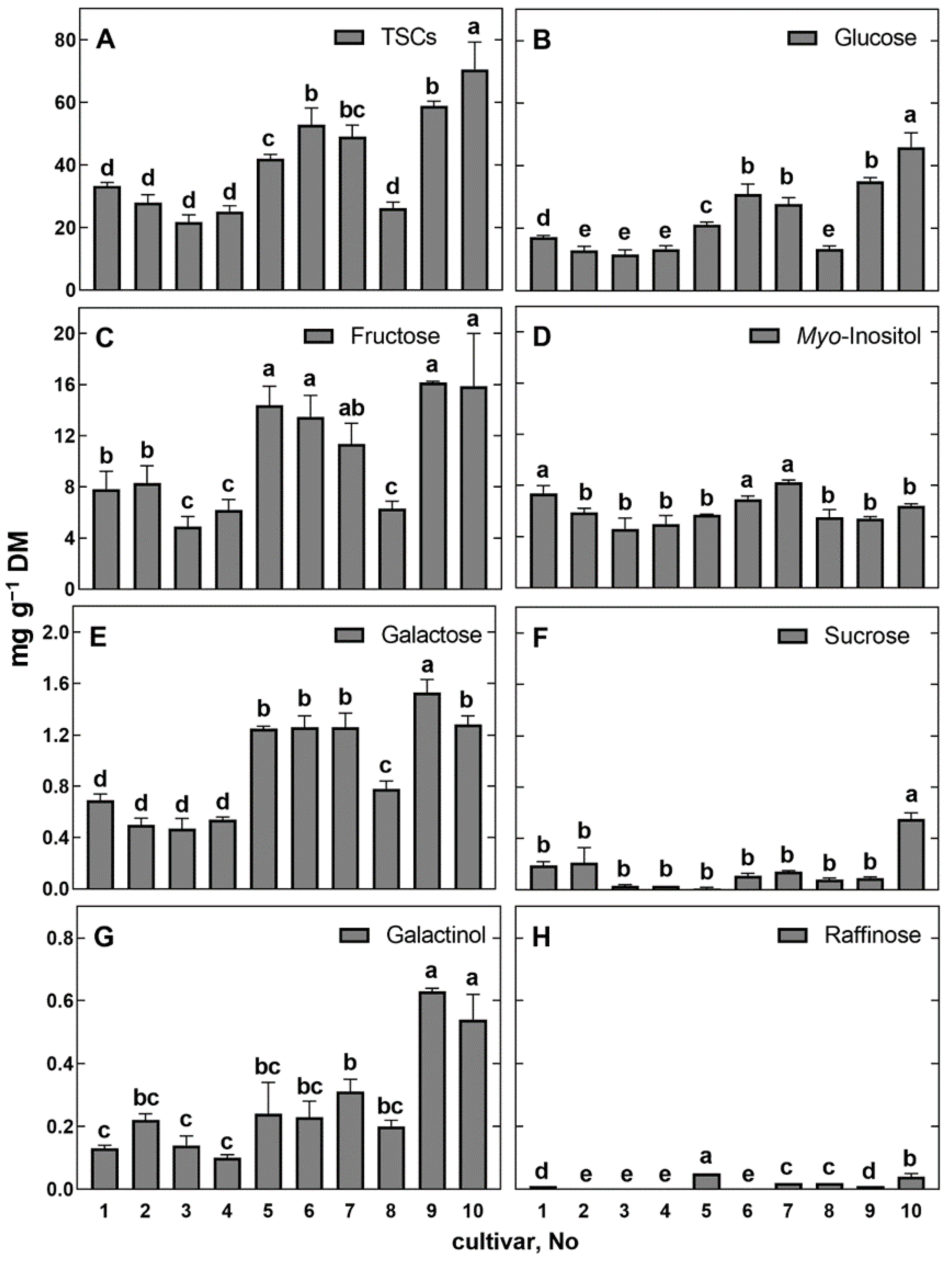
3.4. Content of Soluble Carbohydrates
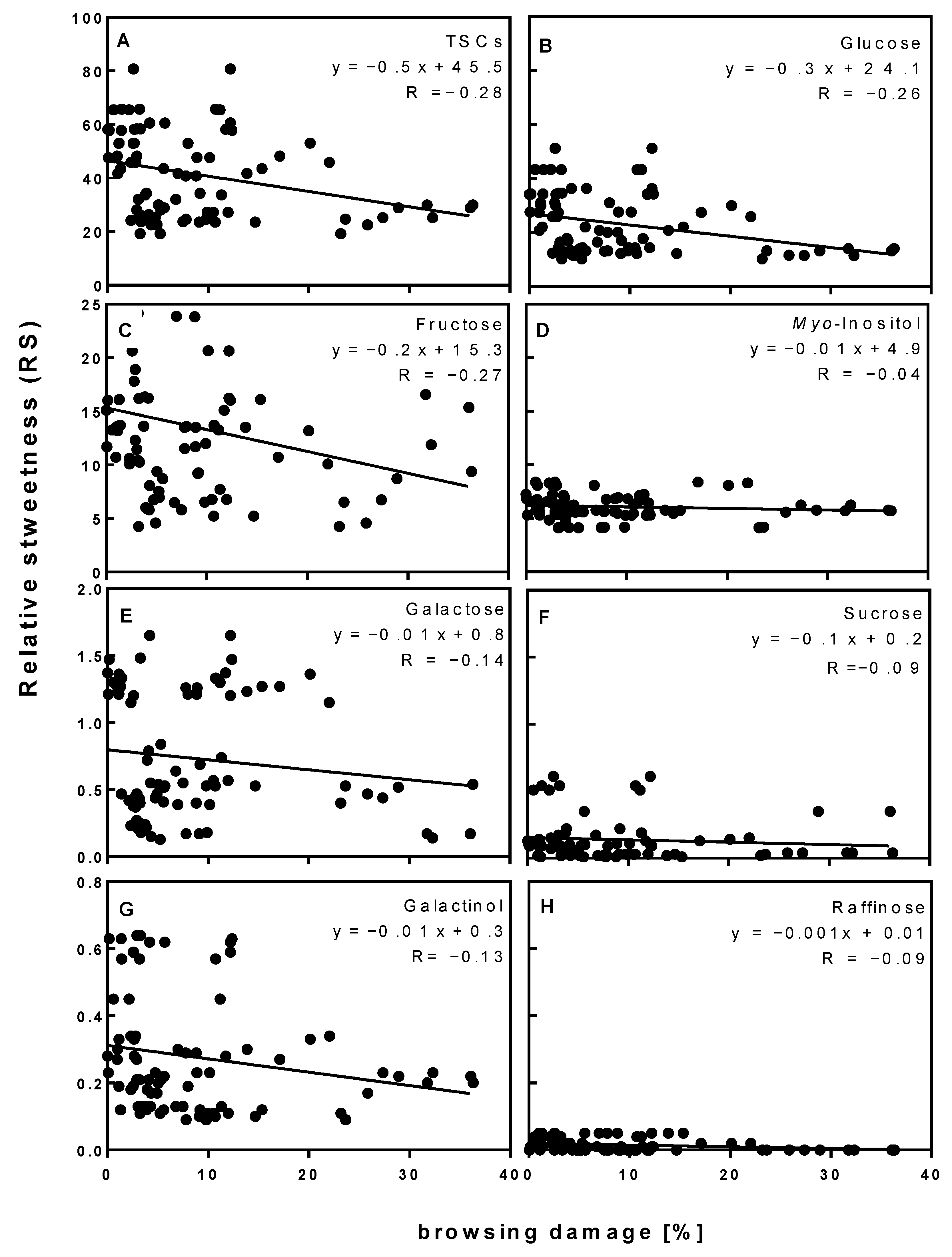
3.5. Shoot Sweetness and Browsing Damage
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Scott, K. Taste Recognition: Food for Thought. Neuron 2005, 48, 455–464. [Google Scholar] [CrossRef] [PubMed]
- State Forests Poland Forests in Poland 2018; Milewski, W., Ed.; The State Forests Information Centre: Warsaw, Poland, 2018; ISBN 978-83-65659-40-8.
- Kokocki, C.; Kowalski, A. Growth and Survival of Some Species of “Forage” Willows in Forest and Non-Forest Habitats; [Wzrost i przeżywalność niektórych gatunków wierzb “paszowych” na siedliskach leśnych i nieleśnych.]; University of Life Sciences: Poznań, Poland, 2012. [Google Scholar]
- Klocek, A.; Hycza, T.; Jodłowski, K.; Kalinowski, M.; Kołakowski, B.; Szmidla, H.; Zachara, T. Forest Research Institute Bulletin No 1/2018. From the Forest World [Z leśnego świata] 2018, 102, 1–4. [Google Scholar]
- Willow—Cultivation, Requirements, Varieties [Wierzba—Uprawa, Wymagania, Odmiany]. Available online: https://www.sadowniczy.pl/Wierzba-uprawa-wymagania-pielegnacja-cinfo-pol-624.html (accessed on 10 July 2020).
- Szczukowski, S.; Tworkowski, J.; Wiwart, M.; Przyborowski, J. Wicker (Salix sp.). Cultivation and Possibilities of Use; [Wiklina (Salix sp.). Uprawa i możliwości wykorzystania.]; Szczukowski, S., Ed.; Wydawnictwo ART: Olsztyn, Poland, 2002; ISBN 8387443409. [Google Scholar]
- Rahmonov, O.; Szczypek, T.; Wach, J. The Blendow desert (Pustynia Błędowska) a unique phenomenon of the Polish landscape. Ann. Geogr. 2006, 39, 34–36. [Google Scholar]
- Wiaderek, I.; Waliszewska, B. Selected mechanical properties of one-year old twigs of Salix acutifolia. Ann. Warsaw Univ. Life Sci. SGGW. For. Wood Technol. 2010, 72, 427–432. [Google Scholar]
- Drogoszewski, B.; Wlazełko, M. The Trial of Determining the Food Preference of Roe Deer (Capreolus capreolus L.) in Relation of Some Willow Varieties; [Próba określania wybiórczości żerowej sarny (Caprcolus capreolus L.) w odniesieniu do nickiórych odmian wierzby]; PTPN: Poznań, Poland, 1980. [Google Scholar]
- Krauze-Baranowska, M.; Szumowicz, E. Willow—The source of antiinflammatory and analgesic medicinal plants. Postępy Fitoter. 2004, 2, 77–86. [Google Scholar]
- Niemiec, P.; Dubas, J.W. Forest Self-Service Pharmacies for Animals. [Leśne samoobsługowe apteki dla zwierzyny]. Brać Low. 2015, 10, 50–51. [Google Scholar]
- Bukiewicz, H. Willow species useful for occlusal plots in forest fisheries. [Gatunki wierzb przydatne na poletka ogryzowo-zgryzowe w łowiskach leśnych.]. Zach. Porad. Low. 1963, 3, 7–8. [Google Scholar]
- Kenstaviciene, P.; Nenortiene, P.; Kiliuviene, G.; Zevzikovas, A.; Lukosius, A.; Kazlauskiene, D. Application of high-performance liquid chromatography for research of salicin in bark of different varieties of Salix. Medicina 2009, 45, 644–651. [Google Scholar] [CrossRef]
- Förster, N.; Ulrichs, C.; Zander, M.; Kätzel, R.; Mewis, I. Factors Influencing the Variability of Antioxidative Phenolic Glycosides in Salix Species. J. Agric. Food Chem. 2010, 58, 8205–8210. [Google Scholar] [CrossRef]
- Sulima, P.; Krauze-Baranowska, M.; Przyborowski, J.A. Variations in the chemical composition and content of salicylic glycosides in the bark of Salix purpurea from natural locations and their significance for breeding. Fitoterapia 2017, 118, 118–125. [Google Scholar] [CrossRef]
- Zalewski, K.; Lahuta, L.B.; Martysiak-Żurowska, D.; Okorski, A.; Nitkiewicz, B.; Zielonka, Ł. Effect of Exogenous Application of Methyl Jasmonate on the Lipid and Carbohydrate Content and Composition of Winter Triticale (Triticosecale Wittm.) Grain and the Severity of Fungal Infections in Triticale Plants and Grain. J. Agric. Food Chem. 2019, 67, 5932–5939. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Villaluenga, C.; Frias, J.; Vidal-Valverde, C. Alpha-Galactosides: Antinutritional Factors or Functional Ingredients? Crit. Rev. Food Sci. Nutr. 2008, 48, 301–316. [Google Scholar] [CrossRef] [PubMed]
- Crittenden, R.G.; Playne, M.J. Production, properties and applications of food-grade oligosaccharides. Trends Food Sci. Technol. 1996, 7, 353–361. [Google Scholar] [CrossRef]
- The Importance of Myo-Inositol in Plants. Available online: https://www.lebanonturf.com/education-center/plant-biostimulants/the-importance-of-myo-inositol-in-plants (accessed on 10 June 2020).
- Cichon, R.; Wądołowska, L. Carbohydrates. In Human Nutrition. Introduction to Nutrition Science; [Żywienie człowieka. Podstawy nauki o żywieniu]; Gawęcki, J., Ed.; Polish Scientific Publishers PWN: Warsaw, Poland, 2010; pp. 149–150. [Google Scholar]
- Sweet Life of a Chemist [Słodkie Życie Chemika]. Available online: https://mlodytechnik.pl./eksperymenty-i-zadania-szkolne/chemia/29068-slodkie-zyciechemika?highlight=WyJzXHUwMTQyb2R5Y3oiXQ (accessed on 10 July 2020).
- Aliferis, K.A.; Chamoun, R.; Jabaji, S. Metabolic responses of willow (Salix purpurea L.) leaves to mycorrhization as revealed by mass spectrometry and 1H NMR spectroscopy metabolite profiling. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef]
- Gąsecka, M.; Mleczek, M.; Drzewiceka, K.; Magdziak, Z.; Rissmann, I.; Chadzinikolau, T.; Golinski, P. Physiological and morphological changes in Salix viminalis L. as a result of plant exposure to copper. J. Environ. Sci. Health Part A 2012, 47, 548–557. [Google Scholar] [CrossRef]
- Corol, D.; Harflett, C.; Beale, M.; Ward, J. An Efficient High Throughput Metabotyping Platform for Screening of Biomass Willows. Metabolites 2014, 4, 946–976. [Google Scholar] [CrossRef]
- Regier, N.; Streb, S.; Zeeman, S.C.; Frey, B. Seasonal changes in starch and sugar content of poplar (Populus deltoides x nigra cv. Dorskamp) and the impact of stem girdling on carbohydrate allocation to roots. Tree Physiol. 2010, 30, 979–987. [Google Scholar] [CrossRef]
- ElSayed, A.I.; Rafudeen, M.S.; Golldack, D. Physiological aspects of raffinose family oligosaccharides in plants: Protection against abiotic stress. Plant Biol. 2014, 16, 1–8. [Google Scholar] [CrossRef]
- Peterbauer, T.; Richter, A. Biochemistry and physiology of raffinose family oligosaccharides and galactosyl cyclitols in seeds. Seed Sci. Res. 2001, 11, 185–197. [Google Scholar] [CrossRef]
- Slewinski, T.L.; Braun, D.M. Current perspectives on the regulation of whole-plant carbohydrate partitioning. Plant Sci. 2010, 178, 341–349. [Google Scholar] [CrossRef]
- Kozlowski, T.T. Carbohydrate sources and sinks in woody plants. Bot. Rev. 1992, 58, 107–222. [Google Scholar] [CrossRef]
- Yarmolinsky, D.A.; Zuker, C.S.; Ryba, N.J.P. Common Sense about Taste: From Mammals to Insects. Cell 2009, 139, 234–244. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.; Owyang, C. Sugars, Sweet Taste Receptors, and Brain Responses. Nutrients 2017, 9, 653. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of the Cultivar | Willow Species | Cultivar No. Used in Tables and Figures |
|---|---|---|
| Resko Forest Inspektorate | S. amygdalina cv. Dunajec | 1 |
| Resko Forest Inspektorate | S. amygdalina cv. Krakowianka | 2 |
| Experimental Station of University of Warmia and Mazury in Obory | S. amygdalina 1045 | 3 |
| Experimental Station of University of Warmia and Mazury in Obory | S. amygdalina 1102 | 4 |
| Experimental Station of University of Warmia and Mazury in Obory | S. amygdalina 1036 | 5 |
| Experimental Station of University of Warmia and Mazury in Obory | S. fragilis cv. Kamon | 6 |
| Resko Forest Inspektorate | S. fragilis cv. Kamon | 7 |
| Experimental Station of University of Warmia and Mazury in Bałdy | S. laurina 220/225 | 8 |
| Experimental Station of University of Warmia and Mazury in Bałdy | S. pentederana | 9 |
| Experimental Station of University of Warmia and Mazury in Obory | S. purpurea 1126 | 10 |
| Sugar | [20] | [21] | Average |
|---|---|---|---|
| Glucose–fructose syrup/relative sweetness | 100 | 100 | 100 |
| Sucrose | 100 | 100 | 100 |
| Fructose | 173 | 180 | 176.50 |
| Glucose | 74 | 75 | 74.50 |
| Galactose | 32.1 | 32 | 32.05 |
| Maltose | 32 | 30 | 31 |
| Mannose | - | 30 | 30 |
| Lactose | 16 | 25 | 20.5 |
| Raffinose | 10 | - | 10 |
| Myo-inositol | 50 | - | 50 |
| Galactinol | 60 | - | 60 |
| Plot | Soil Type | pH (in KCl) | Soil Content (Average of Three Replicates) | ||||
|---|---|---|---|---|---|---|---|
| P205 (mg/100 g of Soil) | K20 (mg/100 g of Soil) | Mg (mg/100 g of Soil) | N Total (%) | C Organic (%) | |||
| Czempin | Loamy sand | 4.98 | 14.8 | 14.0 | 3.6 | 0.086 | 1.98 |
| Bieczyny | Sandy loam | 7.60 | 7.3 | 7.44 | 4.6 | 0.35 | 1.62 |
| Grzybno | Slightly loamy sand | 6.42 | 12.0 | 3.1 | 1.9 | 0.07 | 1.02 |
| Słońsk | Slightly loamy sand | 7.31 | 13.8 | 8.2 | 2.1 | 0.098 | 0.75 |
| Cultivar No. | Deer Czempiń | Deer Kosewo G. | Fallow Deer Kosewo G. |
|---|---|---|---|
| 1 | 10.76 | 0 | 5.94 |
| 2 | 3.91 | 0 | 6.44 |
| 3 | 4.48 | 0 | 3.00 |
| 4 | 18.50 | 1.02 | 4.25 |
| 5 | 14.49 | 13.54 | 8.90 |
| 6 | 8.58 | 0 | 6.39 |
| 7 | 8.99 | 0 | 2.18 |
| 8 | 18.32 | 6.92 | 4.80 |
| 9 | 16.76 | 0 | 0.07 |
| 10 | 4.49 | 0 | 4.72 |
| Cultivar | ||
|---|---|---|
| S. cordata UWM 1036 (July 2019) | S. cordata UWM 1036 (January 2020) | |
| Fructose | 14.37 ± 1.50 | 2.92 ± 0.03 |
| Galactose | 1.25 ± 0.02 | 0.00 |
| Glucose | 21.00 ± 1.02 | 2.04 ± 0.01 |
| Myo-inositol | 5.70 ± 0.07 | 0.43 ± 0.01 |
| Sucrose | 0.01 ± 0.01 | 37.85 ± 0.48 |
| Galactionol | 0.24 ± 0.10 | 0.00 |
| Raffinose | 0.05 ± 0.00 | 14.18 ± 0.68 |
| Stachyose | 0.00 | 5.28 ± 0.20 |
| Verbascose | 0.00 | 0.34 ± 0.02 |
| Total sugars | 41.37 ± 2.40 | 63.03 ± 1.25 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Budny, M.; Zalewski, K.; Lahuta, L.B.; Stolarski, M.J.; Stryiński, R.; Okorski, A. Could the Content of Soluble Carbohydrates in the Young Shoots of Selected Willow Cultivars Be a Determinant of the Plants’ Attractiveness to Cervids (Cervidae, Mammalia)? Agriculture 2021, 11, 67. https://doi.org/10.3390/agriculture11010067
Budny M, Zalewski K, Lahuta LB, Stolarski MJ, Stryiński R, Okorski A. Could the Content of Soluble Carbohydrates in the Young Shoots of Selected Willow Cultivars Be a Determinant of the Plants’ Attractiveness to Cervids (Cervidae, Mammalia)? Agriculture. 2021; 11(1):67. https://doi.org/10.3390/agriculture11010067
Chicago/Turabian StyleBudny, Maciej, Kazimierz Zalewski, Lesław Bernard Lahuta, Mariusz Jerzy Stolarski, Robert Stryiński, and Adam Okorski. 2021. "Could the Content of Soluble Carbohydrates in the Young Shoots of Selected Willow Cultivars Be a Determinant of the Plants’ Attractiveness to Cervids (Cervidae, Mammalia)?" Agriculture 11, no. 1: 67. https://doi.org/10.3390/agriculture11010067
APA StyleBudny, M., Zalewski, K., Lahuta, L. B., Stolarski, M. J., Stryiński, R., & Okorski, A. (2021). Could the Content of Soluble Carbohydrates in the Young Shoots of Selected Willow Cultivars Be a Determinant of the Plants’ Attractiveness to Cervids (Cervidae, Mammalia)? Agriculture, 11(1), 67. https://doi.org/10.3390/agriculture11010067