Sugar Beet Shoot and Root Phenotypic Plasticity to Nitrogen, Phosphorus, Potassium and Lime Omission

 ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Site
2.2. Experimental Design and Fertilizer Management
2.3. Crop Management and Field Data Collection
2.4. Field Data Collection
2.4.1. Leaf Area Index
2.4.2. Shoot and Root Fresh and Dry Weight and Nutrient Concentrations
2.4.3. Root Sampling, Preparation and Scanning
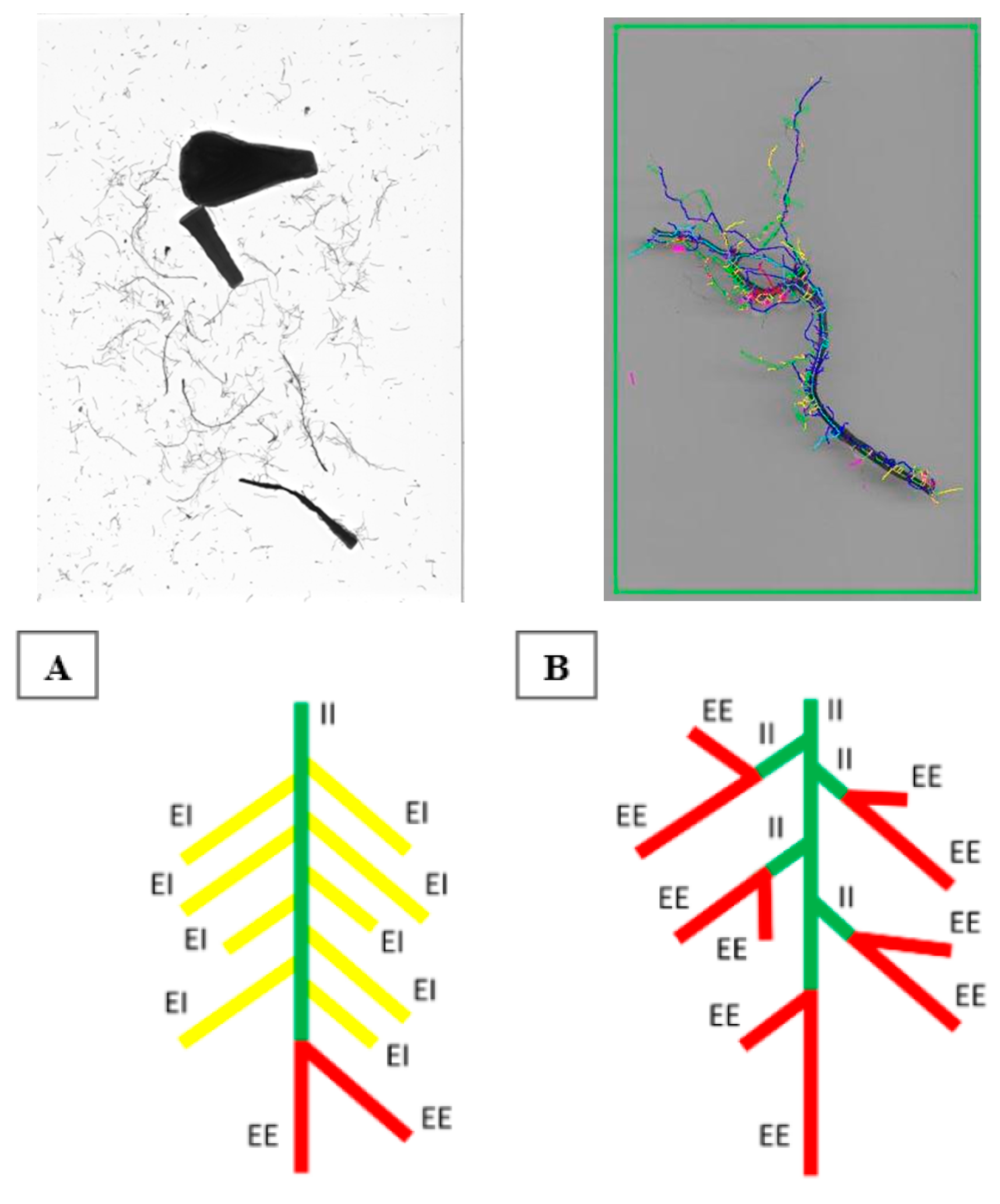
2.4.4. Image Analysis of Root Length, Root Diameter, and Analysis of Basic Link Connectivity
2.4.5. Soil Observations and Analysis
2.5. Data Analyses
3. Results
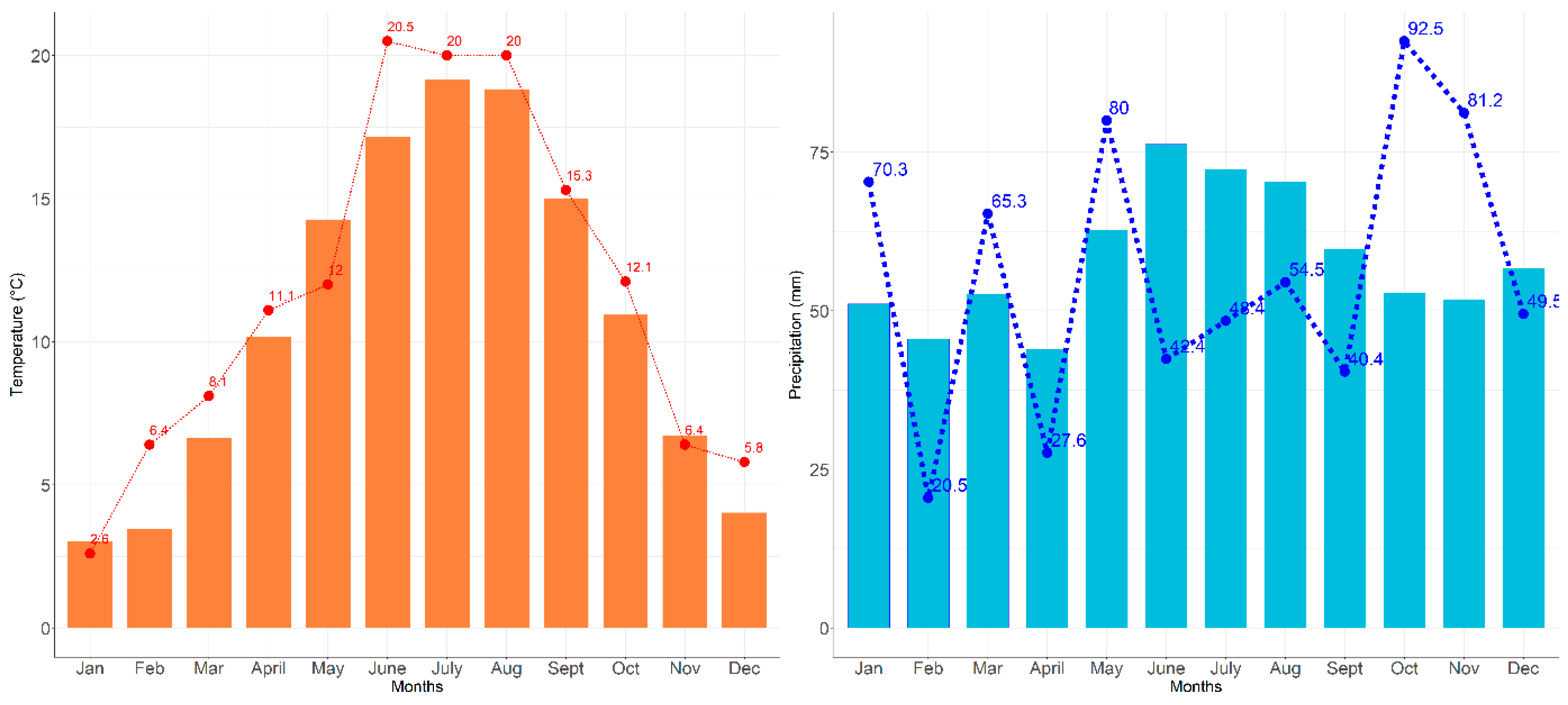
3.1. Growing Conditions during the Growth Period
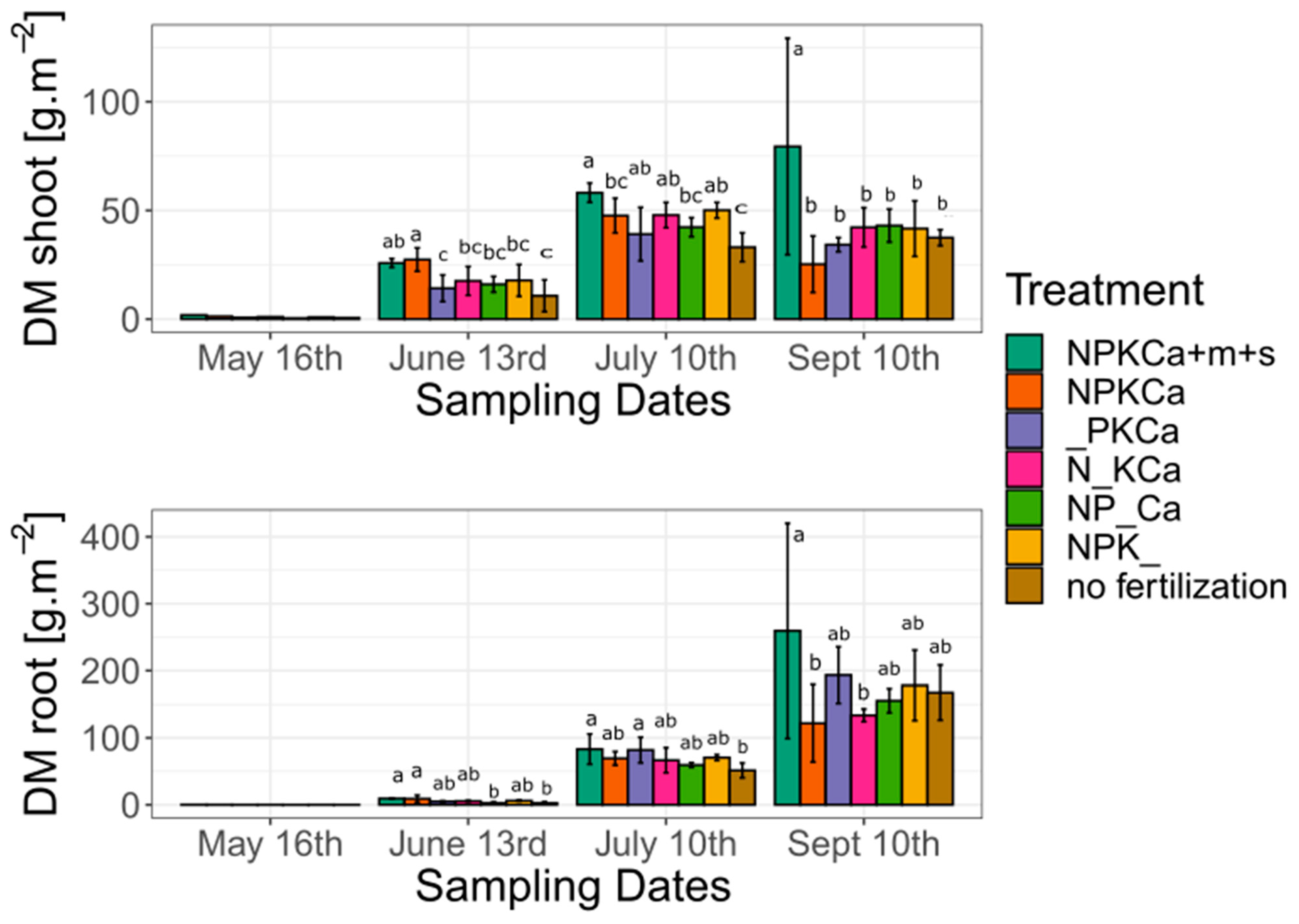
3.2. Shoot Growth
3.3. Root Dry Matter Weight
3.4. Root Shoot Ratio
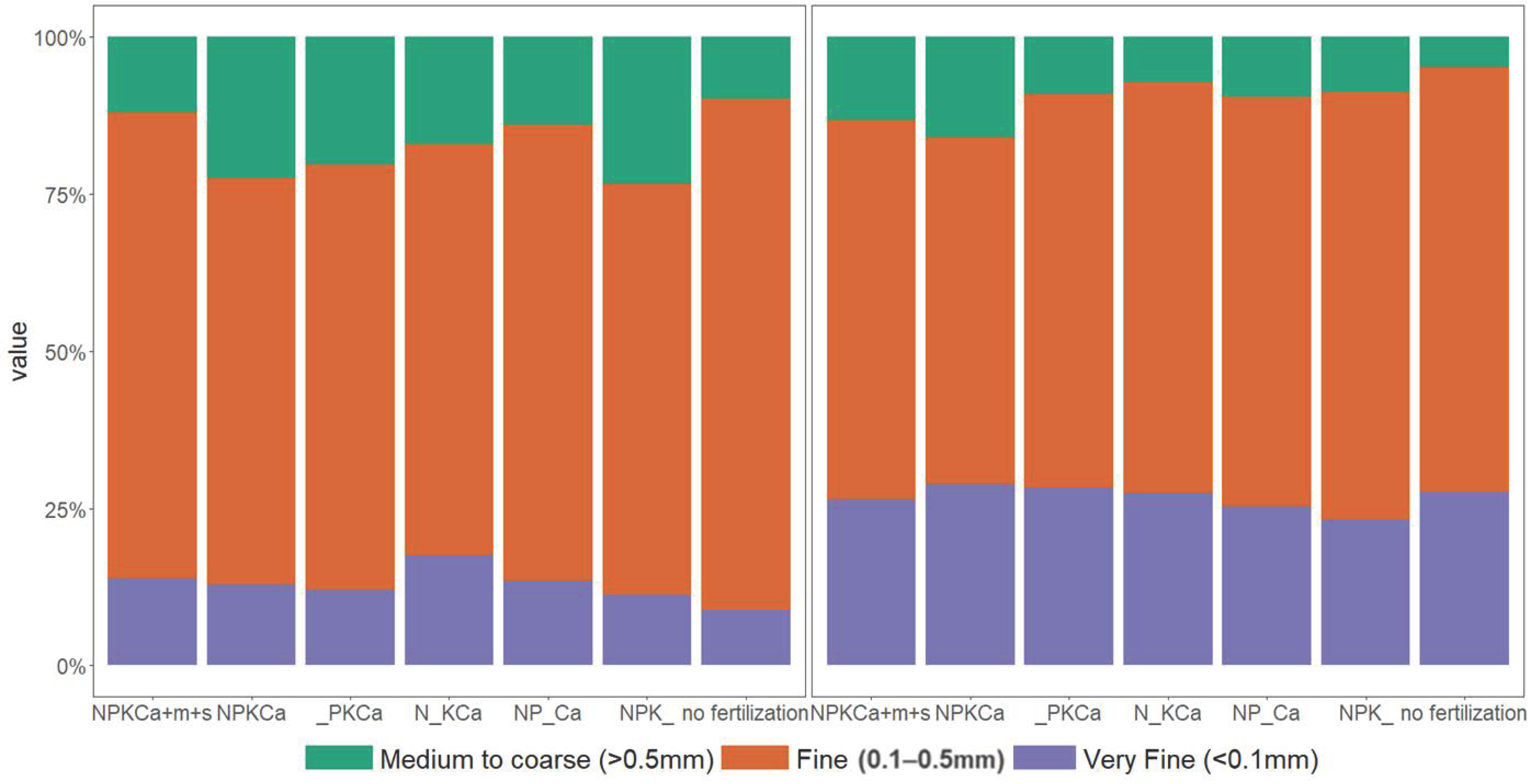
3.5. Root Morphology
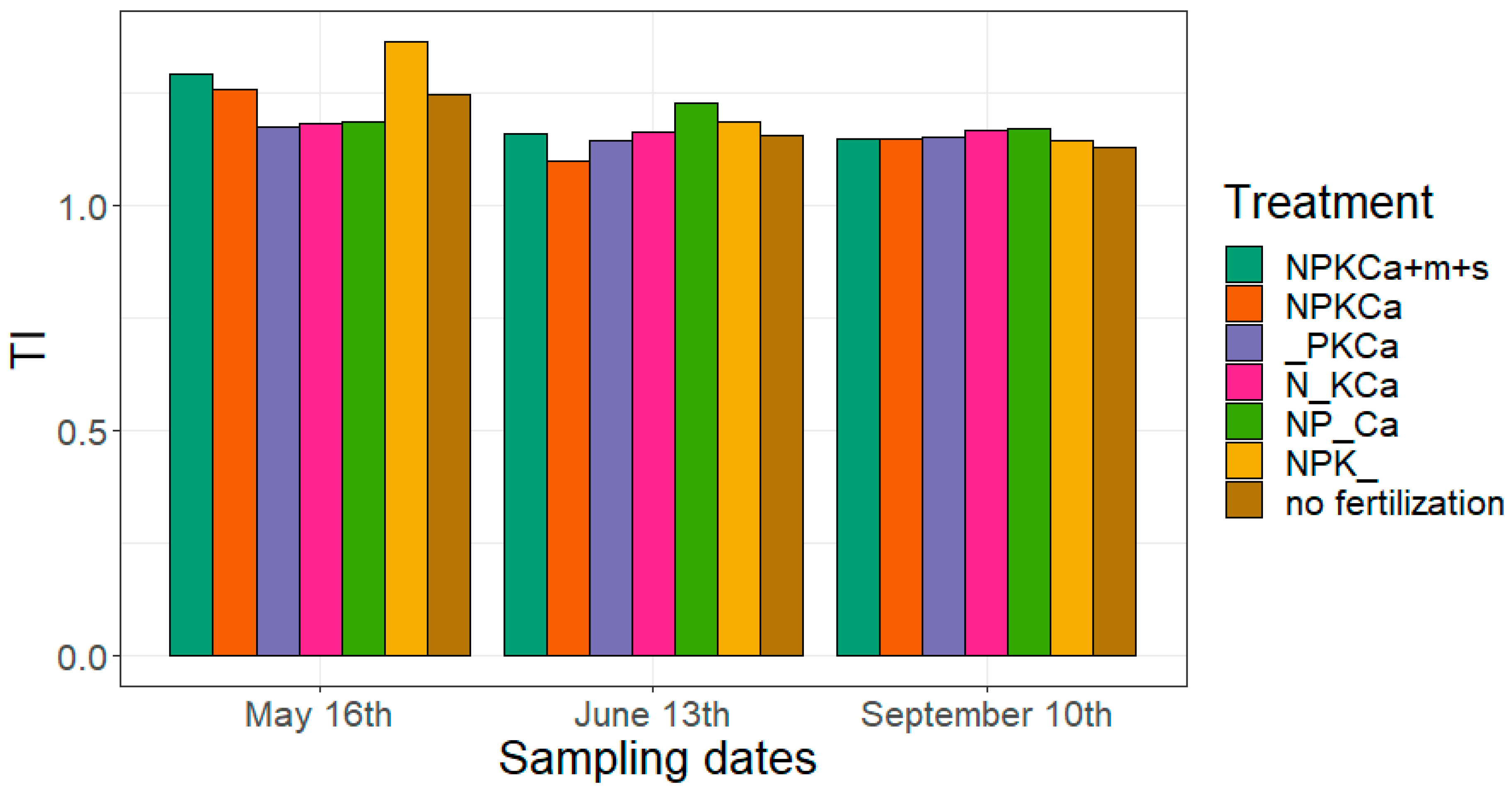
3.6. Root Link Analysis
4. Discussion
4.1. Effect of N, P and K Omission and Omission of Liming on Shoot Growth, Root Shoot Ratio and Yield
4.2. Impact of Nutrient or Liming Omission on Root Morphology and Link Basic Connectivity
4.3. Other Factors Influencing Root Growth and Observations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Goulding, K.; Jarvis, S.; Whitmore, A. Optimizing Nutrient Management for Farm Systems. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 667–680. [Google Scholar] [CrossRef] [PubMed]
- Craufurd, P.Q.; Vadez, V.; Jagadish, S.K.; Prasad, P.V.; Zaman-Allah, M. Crop Science Experiments Designed to Inform Crop Modeling. Agric. For. Meteorol. 2013, 170, 8–18. [Google Scholar] [CrossRef]
- Warren, J.M.; Hanson, P.J.; Iversen, C.M.; Kumar, J.; Walker, A.P.; Wullschleger, S.D. Root Structural and Functional Dynamics in Terrestrial Biosphere Models–Evaluation and Recommendations. New Phytol. 2015, 205, 59–78. [Google Scholar] [CrossRef] [PubMed]
- McCormack, M.L.; Guo, D.; Iversen, C.M.; Chen, W.; Eissenstat, D.M.; Fernandez, C.W.; Li, L.; Ma, C.; Ma, Z.; Poorter, H.; et al. Building a Better Foundation: Improving Root-Trait Measurements to Understand and Model Plant and Ecosystem Processes. New Phytol. 2017, 215, 27–37. [Google Scholar] [CrossRef]
- Bingham, I. Soil-root-canopy Interactions. Ann. Appl. Biol. 2001, 138, 243–251. [Google Scholar] [CrossRef]
- Monteith, J.L. Climate and the Efficiency of Crop Production in Britain. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1977, 281, 277–294. [Google Scholar]
- Allmaras, R.R.; Linden, D.R.; Clapp, C. Corn-Residue Transformations into Root and Soil Carbon as Related to Nitrogen, Tillage, and Stover Management. Soil Sci. Soc. Am. J. 2004, 68, 1366–1375. [Google Scholar] [CrossRef]
- Benjamin, J.G.; Halvorson, A.D.; Nielsen, D.C.; Mikha, M.M. Crop Management Effects on Crop Residue Production and Changes in Soil Organic Carbon in the Central Great Plains. Agron. J. 2010, 102, 990–997. [Google Scholar] [CrossRef]
- Fageria, N.K. The Role of Plant. Roots in Crop. Production; CRC Press: Boca Raton, FL, USA, 2012; ISBN 1-4398-6737-2. [Google Scholar]
- Smit, A.; George, E.; Groenwold, J. Root observations and measurements at (transparent) interfaces with soil. In Root Methods; Springer: Berlin, Heidelberg, 2000; pp. 235–271. [Google Scholar]
- Van Noordwijk, M. Roots: Length, Biomass, Production and Mortality. In Tropical Soil Biology and Fertility: A Handbook of Methods; CAB International Press: Wallingford, UK, 1993; pp. 132–144. [Google Scholar]
- Otto, R.; Trivelin, P.C.O.; Franco, H.C.J.; Faroni, C.E.; Vitti, A.C. Root System Distribution of Sugar Cane as Related to Nitrogen Fertilization, Evaluated by Two Methods: Monolith and Probes. Rev. Bras. Ciência Solo 2009, 33, 601–611. [Google Scholar] [CrossRef]
- Hoffmann, C.M. Changes in Root Morphology with Yield Level of Sugar Beet. Sugar Ind. 2017, 142, 420–425. [Google Scholar] [CrossRef]
- Klimešová, J.; Martínková, J.; Ottaviani, G. Belowground Plant Functional Ecology: Towards an Integrated Perspective. Funct. Ecol. 2018, 32, 2115–2126. [Google Scholar] [CrossRef]
- Li, X.; Zeng, R.; Liao, H. Improving Crop Nutrient Efficiency through Root Architecture Modifications. J. Integr. Plant. Biol. 2016, 58, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, C.M. Importance of Canopy Closure and Dry Matter Partitioning for Yield Formation of Sugar Beet Varieties. Field Crop. Res. 2019, 236, 75–84. [Google Scholar] [CrossRef]
- Fitter, A.; Stickland, T. Architectural Analysis of Plant Root Systems 2. Influence of Nutrient Supply on Architecture in Contrasting Plant Species. New Phytol. 1991, 118, 383–389. [Google Scholar] [CrossRef]
- Carvalho, P.; Foulkes, M.J. Roots and Uptake of Water and Nutrients. In Encyclopedia of Sustainability Science and Technology; Meyers, R.A., Ed.; Springer: New York, NY, USA, 2018; pp. 1–24. ISBN 978-1-4939-2493-6. [Google Scholar]
- Ostonen, I.; Püttsepp, Ü.; Biel, C.; Alberton, O.; Bakker, M.; Lõhmus, K.; Majdi, H.; Metcalfe, D.; Olsthoorn, A.; Pronk, A. Specific Root Length as an Indicator of Environmental Change. Plant. Biosyst. 2007, 141, 426–442. [Google Scholar] [CrossRef]
- Bouma, T.; Nielsen, K.; Koutstaal, B. Sample Preparation and Scanning Protocol for Computerised Analysis of Root Length and Diameter. Plant. Soil 2000, 218, 185–196. [Google Scholar] [CrossRef]
- Berntson, G. A Computer Program for Characterizing Root System Branching Patterns. Plant. Soil 1992, 140, 145–149. [Google Scholar] [CrossRef]
- Fitter, A.H. An Architectural Approach to the Comparative Ecology of Plant Root Systems. New Phytol. 1987, 106, 61–77. [Google Scholar] [CrossRef]
- Kutschera, L. Wurzelatlas Mitteleuropäischer Ackerunkäuter Und Kulturpflanzen; DLG-Verlags-GmbH: Frankfurt, Germany, 1960; p. 249. [Google Scholar]
- Vamerali, T.; Guarise, M.; Ganis, A.; Mosca, G. Effects of Water and Nitrogen Management on Fibrous Root Distribution and Turnover in Sugar Beet. Eur. J. Agron. 2009, 31, 69–76. [Google Scholar] [CrossRef]
- Ulrich, A.; Hills, F.J. Sugar Beet Nutrient Deficiency Symptoms: A Color. Atlas and Chemical Guide; UCANR Publications: Oakland, CA, USA, 1969; Volume 4051. [Google Scholar]
- Terry, N.; Ulrich, A. Effects of Potassium Deficiency on the Photosynthesis and Respiration of Leaves of Sugar Beet. Plant. Physiol. 1973, 51, 783–786. [Google Scholar] [CrossRef]
- Christenson, D.R.; Draycott, A.P. Nutrition-Phosphorus, Sulphur, Potassium, Sodium, Calcium, Magnesium and Micronutrients-Liming and Nutrient Deficiencies. Sugar Beet 2006, 185–220. [Google Scholar] [CrossRef]
- Hergert, G.W. Sugar Beet Fertilization. Sugar Tech. 2010, 12, 256–266. [Google Scholar] [CrossRef]
- Shaw, B.; Thomas, T.; Cooke, D. Responses of Sugar Beet (Beta Vulgaris L.) to Drought and Nutrient Deficiency Stress. Plant. Growth Regul. 2002, 37, 77–83. [Google Scholar] [CrossRef]
- Abdel-Motagally, F.M.; Attia, K.K. Response of Sugar Beet Plants to Nitrogen and Potassium Fertilization in Sandy Calcareous Soil. Int. J. Agric. Biol. 2009, 11, 695–700. [Google Scholar]
- Hodge, A. The Plastic Plant: Root Responses to Heterogeneous Supplies of Nutrients. New Phytol. 2004, 162, 9–24. [Google Scholar] [CrossRef]
- Hetrick, B. Mycorrhizas and Root Architecture. Experientia 1991, 47, 355–362. [Google Scholar] [CrossRef]
- Theurer, J.C. Fibrous Root Growth and Partitioning in Smooth Root Sugarbeet versus Standard Root Type. J. Sugar Beet Res. 1993, 30, 143–150. [Google Scholar] [CrossRef]
- Körschens, M. Die Wihtigsten Dauerfeldversuche Der Welt Übersicht, Bedeutung, Ergebnisse. Arch. Agron. Soil Sci. 1997, 42, 157–168. [Google Scholar] [CrossRef]
- Holthusen, D.; Jänicke, M.; Peth, S.; Horn, R. Physical Properties of a Luvisol for Different Long-term Fertilization Treatments: I. Mesoscale Capacity and Intensity Parameters. J. Plant. Nutr. Soil Sci. 2012, 175, 4–13. [Google Scholar] [CrossRef]
- Rueda-Ayala, V.; Ahrends, H.E.; Siebert, S.; Gaiser, T.; Hüging, H.; Ewert, F. Impact of Nutrient Supply on the Expression of Genetic Improvements of Cereals and Row Crops–A Case Study Using Data from a Long-Term Fertilization Experiment in Germany. Eur. J. Agron. 2018, 96, 34–46. [Google Scholar] [CrossRef]
- Schüller, H. Die CAL-Methode, Eine Neue Methode Zur Bestimmung Des Pflanzenverfügbaren Phosphates in Böden. Z. Für Pflanz. Und Bodenkd. 1969, 123, 48–63. [Google Scholar]
- Fitter, A.H. Functional Significance of Root Morphology and Root System Architecture. Spec. Publ. Ser. Br. Ecol. Soc. 1985, 4, 87–106. [Google Scholar]
- Tsialtas, J.; Maslaris, N. Effect of N Fertilization Rate on Sugar Yield and Non-Sugar Impurities of Sugar Beets (Beta Vulgaris) Grown Under Mediterranean Conditions. J. Agron. Crop. Sci. 2005, 191, 330–339. [Google Scholar] [CrossRef]
- Davidson, R. Effects of Soil Nutrients and Moisture on Root/Shoot Ratios in Lolium Perenne L. and Trifolium Repens L. Ann. Bot. 1969, 33, 571–577. [Google Scholar] [CrossRef]
- Yan, Z.; Eziz, A.; Tian, D.; Li, X.; Hou, X.; Peng, H.; Han, W.; Guo, Y.; Fang, J. Biomass Allocation in Response to Nitrogen and Phosphorus Availability: Insight from Experimental Manipulations of Arabidopsis Thaliana. Front. Plant. Sci. 2019, 10, 598. [Google Scholar] [CrossRef]
- Hermans, C.; Hammond, J.P.; White, P.J.; Verbruggen, N. How Do Plants Respond to Nutrient Shortage by Biomass Allocation? Trends Plant. Sci. 2006, 11, 610–617. [Google Scholar] [CrossRef]
- Claassen, N.; Meyer, D.; Jungk, A. Phosphorus Acquisition of Field Grown Sugar Beet and Its Evaluation with a Simulation Model. In Proceedings of the First Congress of the European Society of Agronomy, Paris, France, 5–7 December 1990. [Google Scholar]
- Yi, J.; Krusenbaum, L.; Unger, P.; Hüging, H.; Seidel, S.J.; Schaaf, G.; Gall, J. Deep Learning for Non-Invasive Diagnosis of Nutrient Deficiencies in Sugar Beet Using RGB Images. Sensors 2020, 20, 5893. [Google Scholar] [CrossRef]
- Kumar, A.; Shahbaz, M.; Koirala, M.; Blagodatskaya, E.; Seidel, S.J.; Kuzyakov, Y.; Pausch, J. Root Trait Plasticity and Plant Nutrient Acquisition in Phosphorus Limited Soil. J. Plant. Nutr. Soil Sci. 2019, 182, 945–952. [Google Scholar] [CrossRef]
- Zhang, Y.; Peng, Y.; Yun-Feng, P.; Xue-Xian, L.; Fan-Jun, C.; Chun-Jian, L. Fine Root Patterning and Balanced Inorganic Phosphorus Distribution in the Soil Indicate Distinctive Adaptation of Maize Plants to Phosphorus Deficiency. Pedosphere 2012, 22, 870–877. [Google Scholar] [CrossRef]
- Shen, Q.; Wen, Z.; Dong, Y.; Li, H.; Miao, Y.; Shen, J. The Responses of Root Morphology and Phosphorus-Mobilizing Exudations in Wheat to Increasing Shoot Phosphorus Concentration. AoB Plants 2018, 10, ply054. [Google Scholar] [CrossRef]
- Steingrobe, B. Root Renewal of Sugar Beet as a Mechanism of P Uptake Efficiency. J. Plant. Nutr. Soil Sci. 2001, 164, 533–539. [Google Scholar] [CrossRef]
- Dunbabin, V.; Diggle, A.; Rengel, Z. Is There an Optimal Root Architecture for Nitrate Capture in Leaching Environments? Plant. Cell Environ. 2003, 26, 835–844. [Google Scholar] [CrossRef]
- Song, X.; Wan, F.; Chang, X.; Zhang, J.; Sun, M.; Liu, Y. Effects of Nutrient Deficiency on Root Morphology and Nutrient Allocation in Pistacia Chinensis Bunge Seedlings. Forests 2019, 10, 1035. [Google Scholar] [CrossRef]
- Mollier, A.; Pellerin, S. Maize Root System Growth and Development as Influenced by Phosphorus Deficiency. J. Exp. Bot. 1999, 50, 487–497. [Google Scholar] [CrossRef]
- Ryser, P. The Mysterious Root Length. Plant. Soil 2006, 286, 1–6. [Google Scholar] [CrossRef]
- Ryser, P. Intra-and Interspecific Variation in Root Length, Root Turnover and the Underlying Parameters. In Variation in Plant Growth; Backhuys Publishers: Leiden, The Netherlands, 1998; pp. 441–465. [Google Scholar]
- Maurel, C.; Nacry, P. Root Architecture and Hydraulics Converge for Acclimation to Changing Water Availability. Nat. Plants 2020, 6, 744–749. [Google Scholar] [CrossRef]
- Fitters, T.F.J.; Mooney, S.J.; Sparkes, D.L. Impact of Water Availability on Root Growth of Sugar Beet Varieties. Soil Use Manag. 2020. [Google Scholar] [CrossRef]
- Rogers, E.D.; Monaenkova, D.; Mijar, M.; Nori, A.; Goldman, D.I.; Benfey, P.N. X-Ray Computed Tomography Reveals the Response of Root System Architecture to Soil Texture. Plant. Physiol. 2016, 171, 2028. [Google Scholar] [CrossRef]
- Watt, M.; McCully, M.E.; Kirkegaard, J.A. Soil Strength and Rate of Root Elongation Alter the Accumulation of Pseudomonas Spp. and Other Bacteria in the Rhizosphere of Wheat. Funct. Plant. Biol. 2003, 30, 483–491. [Google Scholar] [CrossRef]
- Nagel, K.A.; Kastenholz, B.; Jahnke, S.; van Dusschoten, D.; Aach, T.; Mühlich, M.; Truhn, D.; Scharr, H.; Terjung, S.; Walter, A.; et al. Temperature Responses of Roots: Impact on Growth, Root System Architecture and Implications for Phenotyping. Funct. Plant. Biol. 2009, 36, 947–959. [Google Scholar] [CrossRef]
- Rich, S.M.; Watt, M. Soil Conditions and Cereal Root System Architecture: Review and Considerations for Linking Darwin and Weaver. J. Exp. Bot. 2013, 64, 1193–1208. [Google Scholar] [CrossRef]
- Gleeson, S.K.; Good, R.E. Root Growth Response to Water and Nutrients in the New Jersey Pinelands. Can. J. For. Res. 2010, 40, 167–172. [Google Scholar] [CrossRef]
- Perkons, U.; Kautz, T.; Uteau, D.; Peth, S.; Geier, V.; Thomas, K.; Holz, K.L.; Athmann, M.; Pude, R.; Köpke, U. Root-Length Densities of Various Annual Crops Following Crops with Contrasting Root Systems. Soil Tillage Res. 2014, 137, 50–57. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Application Rate per Element | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Sugar Beet (kg ha−1 a−1) | Rotation (kg ha−1) | ||||||||
| Treatment | Type of Fertilizer | N | P | K | Ca | N | P | K | Ca |
| NPKCa + m + s | Synthetic | 120 | 53 | 199 | 0 | 530 | 287 | 1078 | 1293 |
| Manure | 40 | 22 | 83 | 50 | |||||
| NPKCa | Synthetic | 80 | 31 | 116 | 0 | 230 | 155 | 580 | 1143 |
| _PKCa | Synthetic | 0 | 31 | 116 | 0 | 0 | 155 | 580 | 1143 |
| N_KCa | Synthetic | 80 | 0 | 116 | 0 | 230 | 0 | 580 | 1143 |
| NP_Ca | Synthetic | 80 | 31 | 0 | 0 | 230 | 155 | 0 | 1143 |
| NPK_ | Synthetic | 80 | 31 | 116 | 0 | 230 | 155 | 580 | 0 |
| unfertilized | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Parameter | Definition | Unit |
|---|---|---|
| Root length | Total length of all roots present per plant | cm |
| Average root diameter | Average diameter of all roots of one sample | mm |
| L1 | Root length (cm) of very fine roots (diameter < 0.1 mm) | cm |
| L2 | Root length (cm) of fine roots (diameter 0.1–0.5 mm) | cm |
| L3 | Root length (cm) of medium fine roots (diameter > 0.5 mm) | cm |
| EE | Length in cm of exterior-exterior links per cm of total root length | cm |
| EI | Length in cm of exterior-interior links per cm of total root length | cm |
| II | Length in cm of interior-interior links per cm of total root length | cm |
| Shoot Dry Matter Weight (g·plant−1) | |||||||
|---|---|---|---|---|---|---|---|
| NPKCa + m + s | NPKCa | _PKCa | N_KCa | NP_Ca | NPK_ | No Fertilization | |
| 16 May | 1.86 | 1.28 | 0.91 | 1.03 | 0.71 | 0.39 | 0.54 |
| 13 June | 25.80 ± 1.27 ab | 27.39 ± 5.13 a | 14.18 ± 4.93 c | 17.56 ± 5.34 bc | 16.01 ± 2.27 bc | 17.82 ± 5.91 bc | 10.73 ± 5.90 c |
| 10 July | 58.15 ± 1.79 a | 47.62 ± 6.43 ab | 39.09 ± 9.91 bc | 47.85 ± 5.54 ab | 42.28 ± 2.75 bc | 50.08 ± 1.46 ab | 33.06 ± 6.32 c |
| 10 September | 79.41 ± 31.30 a | 25.25 ± 8.16 b | 34.25 ± 1.30 b | 42.20 ± 3.64 b | 43.02 ± 6.12 b | 41.63 ± 8.03 b | 37.47 ± 1.48 b |
| Leaf area index (m−2 m−2) | |||||||
| 13 June | 2.28 ± 0.42 b | 2.89 ± 0.21 a | 1.28 ± 0.16 cd | 1.53 ± 0.19 c | 0.99 ± 0.33 de | 1.66 ± 0.07 c | 0.73 ± 0.24 e |
| 10 July | 3.96 ± 0.56 a | 3.38 ± 0.39 ab | 2.88 ± 0.77 bc | 2.93 ± 0.13 bc | 2.85 ± 0.50 bc | 3.06 ± 0.07 abc | 2.27 ± 0.40 c |
| 10 September | 4.31 ± 2.33 a | 0.91 ± 0.34 b | 1.55 ± 0.47 b | 1.84 ± 0.39 b | 1.35 ± 0.31 b | 1.83 ± 0.35 b | 1.59 ± 0.14 b |
| Root dry matter weight (g·plant−1) | |||||||
| 16 May | 0.3 | 0.22 | 0.17 | 0.18 | 0.14 | 0.07 | 0.1 |
| 13 June | 9.39 ± 0.31 a | 9.02 ± 5.05 a | 5.01 ± 0.69 ab | 5.50 ± 0.78 ab | 2.75 ± 1.25 b | 6.09 ± 0.76 ab | 2.72 ± 0.39 b |
| 10 July | 83.19 ± 14.17 a | 69.34 ± 6.42 ab | 81.82 ± 7.70 a | 66.51 ± 17.83 ab | 59.37 ± 2.22 ab | 0.54 ± 1.75 ab | 51.35 ± 10.55 b |
| 10 September | 259.55 ± 100.98 a | 121.79 ± 46.65 b | 193.48 ± 5.93 ab | 133.57 ± 5.93 b | 155.35 ± 11.28 ab | 178.18 ± 32.99 ab | 167.41 ± 25.80 ab |
| Root-shoot ratio | |||||||
| 16 May | 0.16 | 0.17 | 0.19 | 0.17 | 0.2 | 0.17 | 0.19 |
| 13 June | 0.42 ± 0.11 a | 0.36 ± 0.24 a | 0.39 ± 0.13 a | 0.29 ± 0.05 a | 0.21 ± 0.08 a | 0.37 ± 0.12 a | 0.30 ± 0.05 a |
| 10 July | 1.54 ± 0.16 b | 1.51 ± 0.13 b | 2.07 ± 0.27 a | 1.40 ± 0.38 b | 1.42 ± 0.07 b | 1.55 ± 0.20 b | 1.55 ± 0.14 b |
| 10 September | 3.29 ± 0.27 bc | 4.04 ± 0.65 abc | 5.00 ± 0.27 a | 3.22 ± 0.17 c | 3.27 ± 0.20 bc | 4.29 ± 0.12 ab | 3.83 ± 0.69 bc |
| Fresh matter tuber yield at harvest (t ha−1) | |||||||
| 93.4 | 63.6 | 66.96 | 56.65 | 51.11 | 66.09 | 50.42 | |
| NPKCa + m + s | NPKCa | _PKCa | N_KCa | NP_Ca | NPK_ | No Fertilization | |
|---|---|---|---|---|---|---|---|
| Total root length (cm) | |||||||
| 16 May | 731.00 ± 150.15 a | 559.71 ± 281.50 ab | 225.91 ± 42.63 b | 432.79 ± 233.54 ab | 194.57 ± 91.13 b | 358.86 ± 296.45 ab | 606.51 ± 107.24 ab |
| 13 June | 1096.59 ± 458.37 a | 904.53 ± 143.88 ab | 652.03 ± 90.55 bc | 810.76 ± 152.92 abc | 606.93 ± 41.35 bc | 578.67 ± 98.54 bc | 432.64 ± 158.07 c |
| 10 September | 1658.6 ± 1107.65 a | 1921.9 ± 156.65 a | 1851.31 ± 249.68 a | 2047.73 ± 394.91 a | 2476.75 ± 696.71 a | 1475.9 ± 103.33 * | 1305.11 ± 150.79 a |
| Average root diameter (mm) | |||||||
| 16 May | 0.41 ± 0.04 a | 0.44 ± 0.07 a | 0.46 ± 0.06 a | 0.44 ± 0.05 a | 0.50 ± 0.17 a | 0.50 ± 0.08 a | 0.34 ± 0.00 a |
| 13 June | 0.74 ± 0.19 ab | 0.80 ± 0.03 ab | 0.68 ± 0.04 ab | 0.72 ± 0.10 ab | 0.58 ± 0.11 b | 0.85 ± 0.17 a | 0.82 ± 0.00 ab |
| 10 September | 2.23 ± 1.23 a | 1.14 ± 0.13 a | 1.08 ± 0.36 a | 1.11 ± 0.22 a | 0.93 ± 0.26 a | 0.93 ± 0.44 a | 1.29 ± 0.24 a |
| Topological specific root length (m g−1) | |||||||
| 16 May | 23.73 | 43.14 | 15.89 | 31.84 | 29.62 | 16.87 | 62.55 |
| 13 June | 1.29 ± 0.21 a | 6.15 ± 10.23 a | 1.32 ± 0.28 a | 1.49 ± 0.38 a | 2.94 ± 1.56 a | 0.96 ± 0.21 * | 1.81 ± 0.69 a |
| 10 September | 0.06 ± 0.04 b | 0.22 ± 0.09 a | 0.09 ± 0.01 ab | 0.14 ± 0.00 * | 0.16 ± 0.03 ab | 0.08 ± 0.01 * | 0.08 ± 0.02 b |
| Full Fertilization | No N | No P | No K | No Liming | No Fertilization | |
|---|---|---|---|---|---|---|
| Shoot DM | high */high * | low */low | low */low | |||
| Leaf area index | high */high * | low */ | ||||
| Root DM | high */high * | /high | /low * | low */low | ||
| Root-shoot ratio | high */high * | /low | /low | /low | ||
| Tuber yield | high | low | low | |||
| Total root length | high */ | low * | low */high | low */low | ||
| Root diameter | /low * | /high | ||||
| Specific root length | /low * | high/high | low/ | high/low * | ||
| Share of very fine & fine roots | high/ | /high | low/ | high/high | ||
| Share of interior links | high/ | low/high | low/high | /low | ||
| Number of total links | low/ | low/high | /high | /low | ||
| Topological index | Low/ | /high | /high | high/ | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hadir, S.; Gaiser, T.; Hüging, H.; Athmann, M.; Pfarr, D.; Kemper, R.; Ewert, F.; Seidel, S. Sugar Beet Shoot and Root Phenotypic Plasticity to Nitrogen, Phosphorus, Potassium and Lime Omission. Agriculture 2021, 11, 21. https://doi.org/10.3390/agriculture11010021
Hadir S, Gaiser T, Hüging H, Athmann M, Pfarr D, Kemper R, Ewert F, Seidel S. Sugar Beet Shoot and Root Phenotypic Plasticity to Nitrogen, Phosphorus, Potassium and Lime Omission. Agriculture. 2021; 11(1):21. https://doi.org/10.3390/agriculture11010021
Chicago/Turabian StyleHadir, Sofia, Thomas Gaiser, Hubert Hüging, Miriam Athmann, Daniel Pfarr, Roman Kemper, Frank Ewert, and Sabine Seidel. 2021. "Sugar Beet Shoot and Root Phenotypic Plasticity to Nitrogen, Phosphorus, Potassium and Lime Omission" Agriculture 11, no. 1: 21. https://doi.org/10.3390/agriculture11010021
APA StyleHadir, S., Gaiser, T., Hüging, H., Athmann, M., Pfarr, D., Kemper, R., Ewert, F., & Seidel, S. (2021). Sugar Beet Shoot and Root Phenotypic Plasticity to Nitrogen, Phosphorus, Potassium and Lime Omission. Agriculture, 11(1), 21. https://doi.org/10.3390/agriculture11010021