Muscle Cell Morphogenesis, Structure, Development and Differentiation Processes Are Significantly Regulated during Human Ovarian Granulosa Cells In Vitro Cultivation

 , , , , , , , , ,
, , , , , , , , ,  ,
,  and
and 
Abstract
1. Introduction
2. Experimental Section
2.1. Patients Characteristics
2.2. Long-Term In Vitro Culture of hGCs
2.3. RNA Extraction
2.4. Microarray Expression Analysis
2.5. RT-qPCR Analysis
2.6. Electron Microscopy
2.7. Statistical Analysis
3. Results
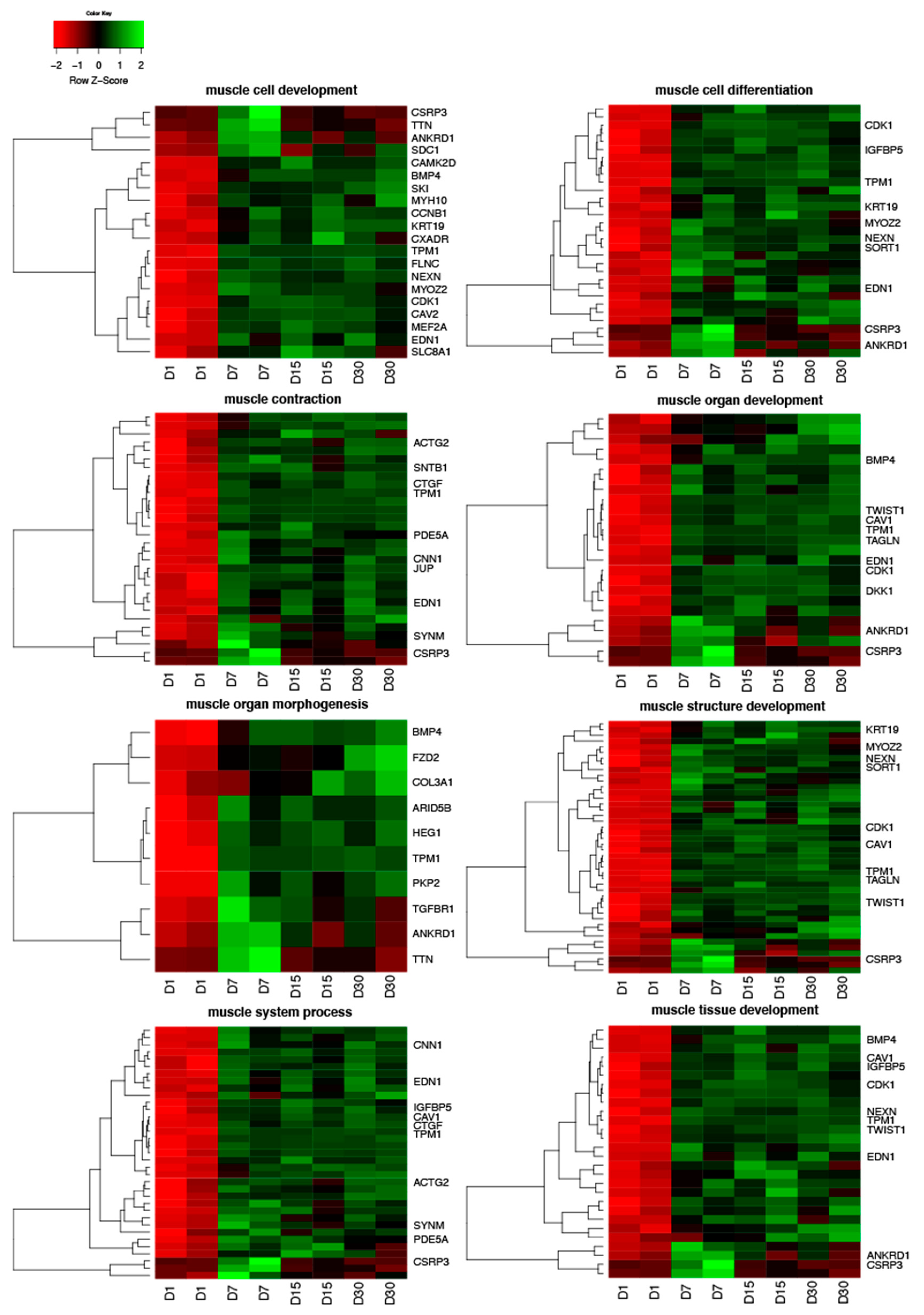
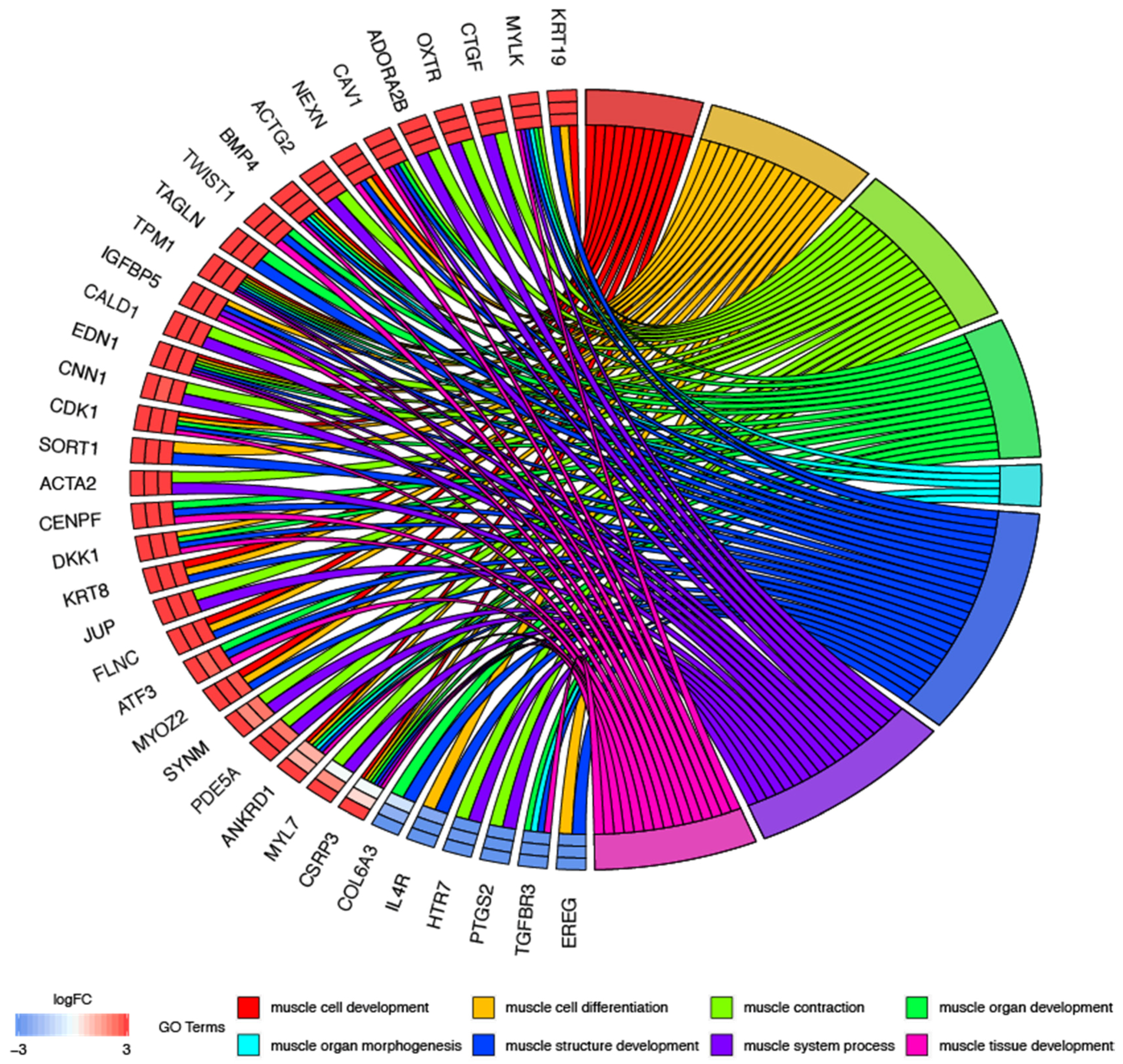
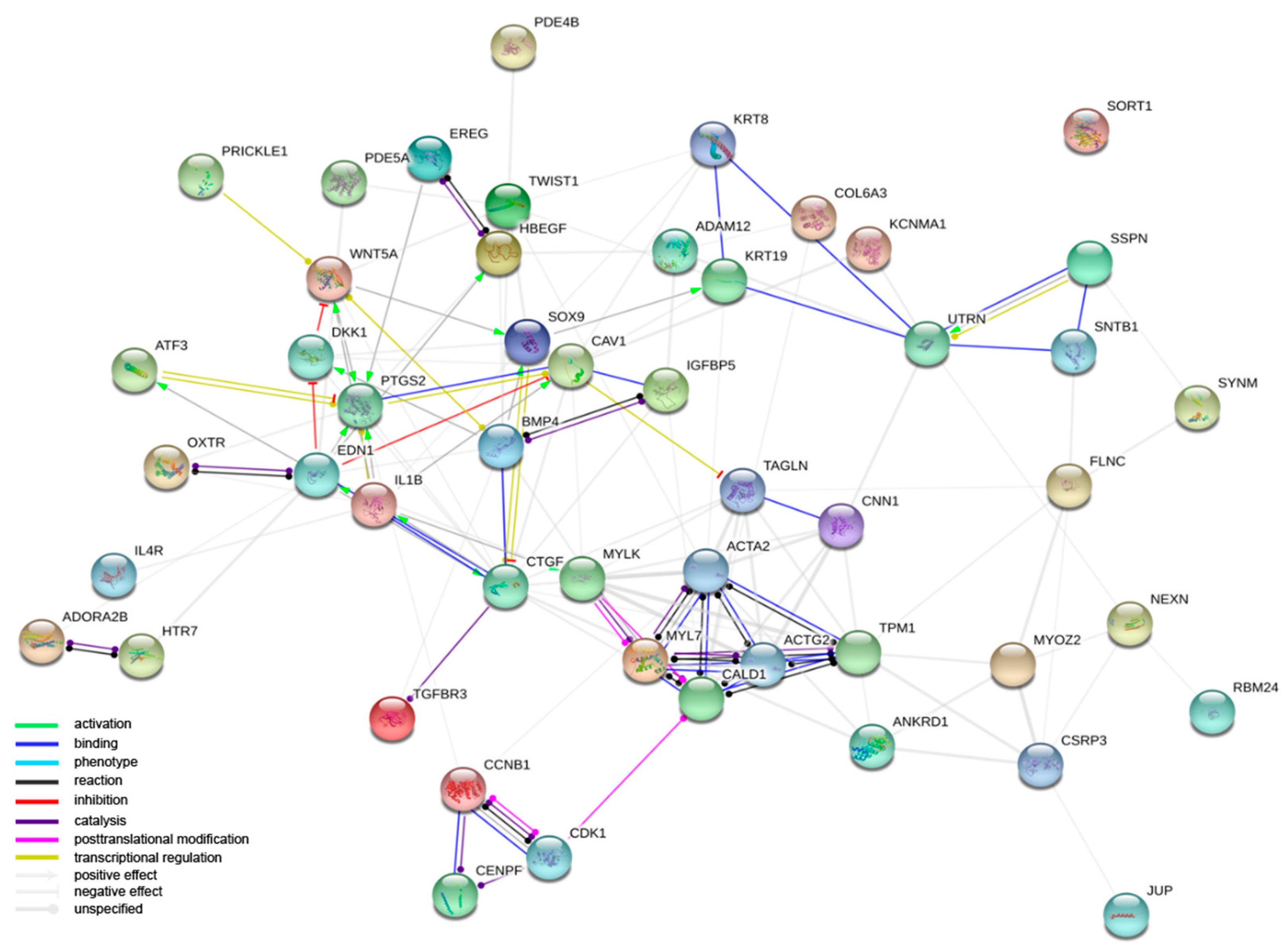
3.1. Microarray Results
3.2. Validation of Gene Expression Direction
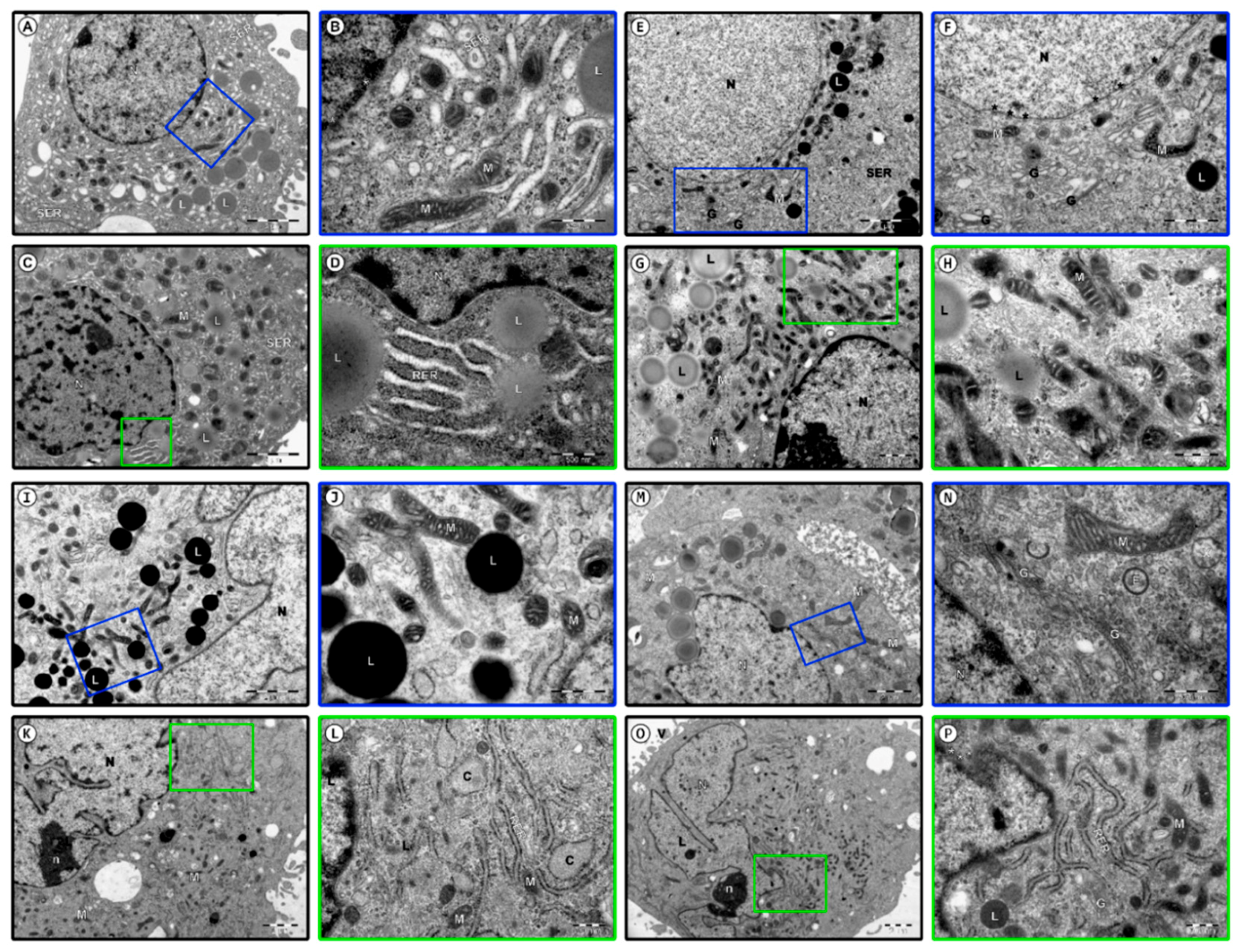
3.3. Ultrastructural Observation of Human Granulosa Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Rybska, M.; Knap, S.; Jankowski, M.; Jeseta, M.; Bukowska, D.; Antosik, P.; Nowicki, M.; Zabel, M.; Kempisty, B.; Jaśkowski, J.M. Characteristic of factors influencing the proper course of folliculogenesis in mammals. Med. J. Cell Biol. 2018, 6, 33–38. [Google Scholar] [CrossRef]
- Rybska, M.; Knap, S.; Jankowski, M.; Jeseta, M.; Bukowska, D. Cytoplasmic and nuclear maturation of oocytes in mammals—Living in the shadow of cells developmental capability. Med. J. Cell Biol. 2018, 1, 13–17. [Google Scholar] [CrossRef]
- Žáková, J.; Crha, I.; Jelínková, L.; Mardešic, T.; Pastor, Z.; Trávník, P.; Kempisty, B.; Ventruba, P.; Ješeta, M. Current topics in assisted reproduction in the Czech Republic. Med. J. Cell Biol. 2018, 6, 61–65. [Google Scholar] [CrossRef]
- Rolf, H.J.; Kierdorf, U.; Kierdorf, H.; Schulz, J.; Seymour, N.; Schliephake, H.; Napp, J.; Niebert, S.; Wölfel, H.; Wiese, K.G. Localization and characterization of STRO-1 cells in the deer pedicle and regenerating antler. PLoS ONE 2008, 3, e2064. [Google Scholar] [CrossRef] [PubMed]
- Mora, J.M.; Fenwick, M.A.; Castle, L.; Baithun, M.; Ryder, T.A.; Mobberley, M.; Carzaniga, R.; Franks, S.; Hardy, K. Characterization and Significance of Adhesion and Junction-Related Proteins in Mouse Ovarian Follicles1. Biol. Reprod. 2012, 86, 153, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Saeed-Zidane, M.; Linden, L.; Salilew-Wondim, D.; Held, E.; Neuhoff, C.; Tholen, E.; Hoelker, M.; Schellander, K.; Tesfaye, D. Cellular and exosome mediated molecular defense mechanism in bovine granulosa cells exposed to oxidative stress. PLoS ONE 2017, 12, e0187569. [Google Scholar] [CrossRef] [PubMed]
- Khamsi, F.; Roberge, S. Granulosa cells of the cumulus oophorus are different from mural granulosa cells in their response to gonadotrophins and IGF-I. J. Endocrinol. 2001, 170, 565–573. [Google Scholar] [CrossRef] [PubMed]
- Kossowska-Tomaszczuk, K.; De Geyter, C.; De Geyter, M.; Martin, I.; Holzgreve, W.; Scherberich, A.; Zhang, H. The Multipotency of Luteinizing Granulosa Cells Collected from Mature Ovarian Follicles. Stem Cells 2009, 27, 210–219. [Google Scholar] [CrossRef]
- Rojewska, M.; Popis, M.; Jankowski, M.; Bukowska, D.; Antosik, P.; Kempisty, B. Stemness specificity of epithelial cells—Application of cell and tissue technology in regenerative medicine. Med. J. Cell Biol. 2018, 6, 114–119. [Google Scholar] [CrossRef]
- Kranc, W.; Brązert, M.; Celichowski, P.; Bryja, A.; Nawrocki, M.J.; Ożegowska, K.; Jankowski, M.; Jeseta, M.; Pawelczyk, L.; Bręborowicz, A.; et al. ‘Heart development and morphogenesis’ is a novel pathway for human ovarian granulosa cell differentiation during long-term in vitro cultivation-a microarray approach. Mol. Med. Rep. 2019, 19, 1705–1715. [Google Scholar] [CrossRef]
- Brevini, T.A.L.; Pennarossa, G.; Rahman, M.M.; Paffoni, A.; Antonini, S.; Ragni, G.; deEguileor, M.; Tettamanti, G.; Gandolfi, F. Morphological and Molecular Changes of Human Granulosa Cells Exposed to 5-Azacytidine and Addressed Toward Muscular Differentiation. Stem Cell Rev. Rep. 2014, 10, 633–642. [Google Scholar] [CrossRef] [PubMed]
- Bruckova, L.; Soukup, T.; Visek, B.; Moos, J.; Moosova, M.; Pavelkova, J.; Rezabek, K.; Kucerova, L.; Micuda, S.; Brcakova, E.; et al. Proliferative potential and phenotypic analysis of long-term cultivated human granulosa cells initiated by addition of follicular fluid. J. Assist. Reprod. Genet. 2011, 28, 939–950. [Google Scholar] [CrossRef] [PubMed]
- Ai, A.; Tang, Z.; Liu, Y.; Yu, S.; Li, B.; Huang, H.; Wang, X.; Cao, Y.; Zhang, W. Characterization and identification of human immortalized granulosa cells derived from ovarian follicular fluid. Exp. Ther. Med. 2019, 18, 2167–2177. [Google Scholar] [CrossRef] [PubMed]
- Brązert, M.; Kranc, W.; Celichowski, P.; Ożegowska, K.; Budna-Tukan, J.; Jeseta, M.; Pawelczyk, L.; Bruska, M.; Zabel, M.; Nowicki, M.; et al. Novel markers of human ovarian granulosa cell differentiation toward osteoblast lineage: A microarray approach. Mol. Med. Rep. 2019, 20, 4403–4414. [Google Scholar] [CrossRef] [PubMed]
- Chamier-Gliszczyńska, A.; Brązert, M.; Sujka-Kordowska, P.; Popis, M.; Ożegowska, K.; Stefańska, K.; Kocherova, I.; Celichowski, P.; Kulus, M.; Bukowska, D.; et al. Genes involved in angiogenesis and circulatory system development are differentially expressed in porcine epithelial oviductal cells during long-term primary in vitro culture—A transcriptomic study. Med. J. Cell Biol. 2018, 6, 163–173. [Google Scholar] [CrossRef]
- Budna, J.; Celichowski, P.; Bryja, A.; Jeseta, M.; Jankowski, M.; Bukowska, D.; Antosik, P.; Nowicki, A.; Brüssow, K.P.; Bruska, M.; et al. Expression Changes in Fatty acid Metabolic Processrelated Genes in Porcine Oocytes During in Vitro Maturation. Med. J. Cell Biol. 2018, 6, 48–54. [Google Scholar] [CrossRef]
- Kranc, W.; Brązert, M.; Ożegowska, K.; Budna-Tukan, J.; Celichowski, P.; Jankowski, M.; Bryja, A.; Nawrocki, M.J.; Popis, M.; Jeseta, M.; et al. Response to abiotic and organic substances stimulation belongs to ontologic groups significantly up-regulated in porcine immature oocytes. Med. J. Cell Biol. 2018. [Google Scholar] [CrossRef]
- Kulus, M.; Józkowiak, M.; Kulus, J.; Popis, M.; Borowiec, B.; Stefańska, K.; Celichowski, P.; Nawrocki, M.J.; Bukowska, D.; Brüssow, K.P.; et al. “Cell Cycle Process”, “Cell Division” and “Cell Proliferation” Belong To Ontology Groups Highly Regulated During Long-Term Culture of Porcine Oviductal Epithelial Cells. Med. J. Cell Biol. 2019, 7, 15–24. [Google Scholar] [CrossRef]
- Stefańska, K.; Kocherova, I.; Knap, S.; Kulus, M.; Celichowski, P.; Jeseta, M. The genes regulating maintenance of cellular protein location are differentially expressed in porcine epithelial oviductal cells during longterm in vitro cultivation. Med. J. Cell Biol. 2019, 7, 77–85. [Google Scholar] [CrossRef]
- Chomczynski, P.; Sacchi, N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef]
- Nawrocki, M.J.; Celichowski, P.; Jankowski, M.; Kranc, W.; Bryja, A.; Borys-Wójcik, S.; Jeseta, M.; Antosik, P.; Bukowska, D.; Bruska, M.; et al. Ontology groups representing angiogenesis and blood vessels development are highly up-regulated during porcine oviductal epithelial cells long-term real-time proliferation—A primary cell culture approach. Med. J. Cell Biol. 2018, 6, 186–194. [Google Scholar] [CrossRef]
- Bryja, A.; Dyszkiewicz-Konwińska, M.; Jankowski, M.; Celichowski, P.; Stefańska, K.; Chamier-Gliszczyńska, A.; Popis, M.; Mehr, K.; Bukowska, D.; Antosik, P.; et al. Ion homeostasis and transport are regulated by genes differentially expressed in porcine buccal pouch mucosal cells during long-term culture in vitro-a microarray approach. Bryja al. Med. J. Cell Biol. 2018, 6, 75–82. [Google Scholar] [CrossRef]
- Walter, W.; Sánchez-Cabo, F.; Ricote, M. GOplot: An R package for visually combining expression data with functional analysis. Bioinformatics 2015, 31, 2912–2914. [Google Scholar] [CrossRef] [PubMed]
- Virant-Klun, I.; Zech, N.; Rožman, P.; Vogler, A.; Cvjetičanin, B.; Klemenc, P.; Maličev, E.; Meden-Vrtovec, H. Putative stem cells with an embryonic character isolated from the ovarian surface epithelium of women with no naturally present follicles and oocytes. Differentiation 2008, 76, 843–856. [Google Scholar] [CrossRef]
- Virant-Klun, I.; Rožman, P.; Cvjeticanin, B.; Vrtacnik-Bokal, E.; Novakovic, S.; Rülicke, T.; Dovc, P.; Meden-Vrtovec, H. Parthenogenetic Embryo-Like Structures in the Human Ovarian Surface Epithelium Cell Culture in Postmenopausal Women with No Naturally Present Follicles and Oocytes. Stem Cells Dev. 2009, 18, 137–150. [Google Scholar] [CrossRef] [PubMed]
- Parte, S.; Bhartiya, D.; Telang, J.; Daithankar, V.; Salvi, V.; Zaveri, K.; Hinduja, I. Detection, Characterization, and Spontaneous Differentiation In Vitro of Very Small Embryonic-Like Putative Stem Cells in Adult Mammalian Ovary. Stem Cells Dev. 2011, 20, 1451–1464. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.; Canning, J.; Kaneko, T.; Pru, J.K.; Tilly, J.L. Germline stem cells and follicular renewal in the postnatal mammalian ovary. Nature 2004, 428, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Dzafic, E.; Stimpfel, M.; Virant-Klun, I. Plasticity of granulosa cells: On the crossroad of stemness and transdifferentiation potential. J. Assist. Reprod. Genet. 2013, 30, 1255–1261. [Google Scholar] [CrossRef] [PubMed]
- Wagner, M.; Yoshihara, M.; Douagi, I.; Damdimopoulos, A.; Panula, S.; Petropoulos, S.; Lu, H.; Pettersson, K.; Palm, K.; Katayama, S.; et al. Single-cell analysis of human ovarian cortex identifies distinct cell populations but no oogonial stem cells. Nat. Commun. 2020, 11. [Google Scholar] [CrossRef]
- Ignatiadis, M.; Xenidis, N.; Perraki, M.; Apostolaki, S.; Politaki, E.; Kafousi, M.; Stathopoulos, E.N.; Stathopoulou, A.; Lianidou, E.; Chlouverakis, G.; et al. Different prognostic value of cytokeratin-19 mRNA-positive circulating tumor cells according to estrogen receptor and HER2 status in early-stage breast cancer. J. Clin. Oncol. 2007, 25, 5194–5202. [Google Scholar] [CrossRef]
- Bozionellou, V.; Mavroudis, D.; Perraki, M.; Papadopoulos, S.; Apostolaki, S.; Stathopoulos, E.; Stathopoulou, A.; Lianidou, E.; Georgoulias, V. Trastuzumab administration can effectively target chemotherapy-resistant cytokeratin-19 messenger RNA-positive tumor cells in the peripheral blood and bone marrow of patients with breast cancer. Clin. Cancer Res. 2004, 10, 8185–8194. [Google Scholar] [CrossRef] [PubMed]
- Bártek, J.; Bártková, J.; Taylor-Papadimitriou, J.; Rejthar, A.; Kovařík, J.; Lukáš, Z.; Vojtěšek, B. Differential expression of keratin 19 in normal human epithelial tissues revealed by monospecific monoclonal antibodies. Histochem. J. 1986, 18, 565–575. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Yasuchika, K.; Ishii, T.; Katayama, H.; Yoshitoshi, E.Y.; Ogiso, S.; Kita, S.; Yasuda, K.; Fukumitsu, K.; Mizumoto, M.; et al. Keratin 19, a cancer stem cell marker in human hepatocellular carcinoma. Clin. Cancer Res. 2015, 21, 3081–3091. [Google Scholar] [CrossRef] [PubMed]
- Coulombe, P.A.; Wong, P. Cytoplasmic intermediate filaments revealed as dynamic and multipurpose scaffolds. Nat. Cell Biol. 2004, 6, 699–706. [Google Scholar] [CrossRef] [PubMed]
- Sanborn, B.M.; Dodge, K.; Monga, M.; Qian, A.; Wang, W.; Yue, C. Molecular mechanisms regulating the effects of oxytocin on myometrialintracellular calcium. Adv. Exp. Med. Biol. 1998, 449, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Zingg, H.H.; Laporte, S.A. The oxytocin receptor. Trends Endocrinol. Metab. 2003, 14, 222–227. [Google Scholar] [CrossRef]
- Elabd, C.; Cousin, W.; Upadhyayula, P.; Chen, R.Y.; Chooljian, M.S.; Li, J.; Kung, S.; Jiang, K.P.; Conboy, I.M. Oxytocin is an age-specific circulating hormone that is necessary for muscle maintenance and regeneration. Nat. Commun. 2014, 5, 4082. [Google Scholar] [CrossRef]
- Moussad, E.E.D.A.; Brigstock, D.R. Connective tissue growth factor: What’s in a name? Mol. Genet. Metab. 2000, 71, 276–292. [Google Scholar] [CrossRef]
- Hishikawa, K.; Nakaki, T.; Fujii, T. Connective tissue growth factor induces apoptosis via caspase 3 in cultured human aortic smooth muscle cells. Eur. J. Pharmacol. 2000, 392, 19–22. [Google Scholar] [CrossRef]
- Gao, R.; Ball, D.A.K.; Perbal, B.; Brigstock, D.R. Connective tissue growth factor induces c-fos gene activation and cell proliferation through p44/42 MAP kinase in primary rat hepatic stellate cells. J. Hepatol. 2004, 40, 431–438. [Google Scholar] [CrossRef]
- Shimo, T.; Nakanishi, T.; Nishida, T.; Asano, M.; Kanyama, M.; Kuboki, T.; Tamatani, T.; Tezuka, K.; Takemura, M.; Matsumura, T.; et al. Connective Tissue Growth Factor Induces the Proliferation, Migration, and Tube Formation of Vascular Endothelial Cells In Vitro, and Angiogenesis In Vivo. J. Biochem. 1999, 126, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.; Haginoya, K.; Wu, Y.; Chiba, Y.; Nakanishi, T.; Onuma, A.; Sato, Y.; Takigawa, M.; Iinuma, K.; Tsuchiya, S. Connective tissue growth factor is overexpressed in muscles of human muscular dystrophy. J. Neurol. Sci. 2008, 267, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Hishikawa, K.; Oemar, B.S.; Tanner, F.C.; Nakaki, T.; Fujii, T.; Lüscher, T.F. Overexpression of connective tissue growth factor gene induces apoptosis in human aortic smooth muscle cells. Circulation 1999, 100, 2108–2112. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.H.; Pech, M.; Karnovsky, M.J. Connective tissue growth factor (CTGF) stimulates vascular smooth muscle cell growth and migration in vitro. Eur. J. Cell Biol. 2000, 79, 915–923. [Google Scholar] [CrossRef] [PubMed]
- Hassel, D.; Dahme, T.; Erdmann, J.; Meder, B.; Huge, A.; Stoll, M.; Just, S.; Hess, A.; Ehlermann, P.; Weichenhan, D.; et al. Nexilin mutations destabilize cardiac Z-disks and lead to dilated cardiomyopathy. Nat. Med. 2009, 15, 1281–1288. [Google Scholar] [CrossRef] [PubMed]
- Ohtsuka, T.; Nakanishi, H.; Ikeda, W.; Satoh, A.; Momose, Y.; Nishioka, H.; Takai, Y. Nexilin: A novel actin filament-binding protein localized at cell- matrix adherens junction. J. Cell Biol. 1998, 143, 1227–1238. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.; Hakuno, F.; Northcott, P.; Pessin, J.E.; Adcock, M.R. Nexilin, a Cardiomyopathy-Associated F-Actin Binding Protein, Binds and Regulates IRS1 Signaling in Skeletal Muscle Cells. PLoS ONE 2013, 8, e55634. [Google Scholar] [CrossRef]
- Arber, S.; Halder, G.; Caroni, P. Muscle LIM protein, a novel essential regulator of myogenesis, promotes myogenic differentiation. Cell 1994, 79, 221–231. [Google Scholar] [CrossRef]
- Vafiadaki, E.; Arvanitis, D.A.; Papalouka, V.; Terzis, G.; Roumeliotis, T.I.; Spengos, K.; Garbis, S.D.; Manta, P.; Kranias, E.G.; Sanoudou, D. Muscle lim protein isoform negatively regulates striated muscle actin dynamics and differentiation. FEBS J. 2014, 281, 3261–3279. [Google Scholar] [CrossRef]
- Geeves, M.A. 4.13 Thin Filament Regulation. In Comprehensive Biophysics; Elsevier: Amsterdam, The Netherlands, 2012; pp. 251–267. [Google Scholar]
- Lehrer, S.S.; Geeves, M.A. The myosin-activated thin filament regulatory state, M-open: A link to hypertrophic cardiomyopathy (HCM). J. Muscle Res. Cell Motil. 2014, 35, 153–160. [Google Scholar] [CrossRef]
- Gordon, A.M.; Homsher, E.; Regnier, M. Regulation of contraction in striated muscle. Physiol. Rev. 2000, 80, 853–924. [Google Scholar] [CrossRef] [PubMed]
- Kamm, K.E.; Stull, J.T. Dedicated Myosin Light Chain Kinases with Diverse Cellular Functions. J. Biol. Chem. 2001, 276, 4527–4530. [Google Scholar] [CrossRef] [PubMed]
- Khromov, A.S.; Wang, H.; Choudhury, N.; McDuffie, M.; Herring, B.P.; Nakamoto, R.; Owens, G.K.; Somlyo, A.P.; Somlyo, A.V. Smooth muscle of telokin-deficient mice exhibits increased sensitivity to Ca2+ and decreased cGMP-induced relaxation. Proc. Natl. Acad. Sci. USA 2006, 103, 2440–2445. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, T.; Schlegel, A.; Scherer, P.E.; Lisanti, M.P. Caveolins, a family of scaffolding proteins for organizing “preassembled signaling complexes” at the plasma membrane. J. Biol. Chem. 1998, 273, 5419–5422. [Google Scholar] [CrossRef] [PubMed]
- Volonte, D.; Liu, Y.; Galbiati, F. The modulation of caveolin-1 expression controls satellite cell activation during muscle repair. FASEB J. 2005, 19, 237–239. [Google Scholar] [CrossRef] [PubMed]
- Taylor, D.S.; Cheng, X.; Pawlowski, J.E.; Wallace, A.R.; Ferrer, P.; Molloy, C.J. Epiregulin is a potent vascular smooth muscle cell-derived mitogen induced by angiotensin II, endothelin-1, and thrombin. Proc. Natl. Acad. Sci. USA 1999, 96, 1633–1638. [Google Scholar] [CrossRef]
- Takahashi, M.; Hayashi, K.; Yoshida, K.; Ohkawa, Y.; Komurasaki, T.; Kitabatake, A.; Ogawa, A.; Nishida, W.; Yano, M.; Monden, M.; et al. Epiregulin as a major autocrine/paracrine factor released from ERK- and p38MAPK-activated vascular smooth muscle cells. Circulation 2003, 108, 2524–2529. [Google Scholar] [CrossRef]
- Sekiguchi, T.; Yamada, K.; Yazawa, T.; Miyamoto, K. Expression of Epiregulin and Amphiregulin in the Rat Ovary. J. Mol. Endocrinol. 2004, 33, 281–291. [Google Scholar]
- López-Casillas, F.; Riquelme, C.; Pérez-Kato, Y.; Veronica Ponce-Castaneda, M.; Osses, N.; Esparza-Lopez, J.; Gonzalez-Nunez, G.; Cabello-Verrugio, C.; Mendoza, V.; Troncoso, V.; et al. Betaglycan expression is transcriptionally up-regulated during skeletal muscle differentiation: Cloning of murine betaglycan gene promoter and its modulation by MyoD, retinoic acid, and transforming growth factor-β. J. Biol. Chem. 2003, 278, 382–390. [Google Scholar] [CrossRef]
- da Luz, C.M.; da Broi, M.G.; Donabela, F.C.; Paro de Paz, C.C.; Meola, J.; Navarro, P.A. PTGS2 down-regulation in cumulus cells of infertile women with endometriosis. Reprod. Biomed. Online 2017, 35, 379–386. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Abbreviation | Primer Sequences F | Primer Sequences R | Ensembl Accession Number | Product Size (bp) |
|---|---|---|---|---|---|
| Oxytocin receptor | OXTR | TTCTTCGTGCAGATGTGGAG | GGACGAGTTGCTCTTTTTGC | ENSG00000111640 | 234 |
| Keratin 19 | KRT19 | TTTGAGACGGAACAGGCTCT | AATCCACCTCCACACTGACC | ENSG00000171345 | 211 |
| Epiregulin | EREG | CCAAGGACGGAAAATGCTTA | AAAATTAGCTGGGCATGGTG | ENSG00000124882 | 237 |
| Wingless-type MMTV integration site family, member 5A | WNT5A | TGGCTTTGGCCATATTTTTC | CCGATGTACTGCATGTGGTC | ENSG00000114251 | 199 |
| Transforming growth factor, beta receptor III | TGFBR3 | CCAAGATGAATGGCACACAC | CCATCTGGCCAACCACTACT | ENSG00000069702 | 151 |
| Prostaglandin-enderoperoxide synthase 2 | PTGS2 | TGAGCATCTACGGTTTGCTG | TGCTTGTCTGGAACAACTGC | ENSG00000073756 | 158 |
| Caveolin 1 | CAV1 | TCTCTACACCGTTCCCATCC | CAATCTTGACCACGTCATCG | ENSG00000105974 | 164 |
| Nexilin | NEXN | AAAAGAAGGCGTTTGCTGAA | CCTCTTCCTCTCCCATTTCC | ENSG00000162614 | 240 |
| Myosin light chain kinase | MYLK | TTGCTGAGGAAAAGCCTCAT | TTCCCGTCCTCATCGTAGTC | ENSG00000065534 | 193 |
| Cysteine and glycine-rich protein 3 | CSRP3 | CCTTGGCACAAGACCTGTTT | TTGTGTAAGGCCTCCAAACC | ENSG00000129170 | 150 |
| Actin, gamma 2, smooth muscle, enteric | ACTG2 | ACCCACAATGTCCCCATCTA | CTCCTTGATGTCTCGCACAA | ENSG00000163017 | 165 |
| Activating transcription factor 3 | ATF3 | CGCTGGAATCAGTCACTGTC | AGGCACTCCGTCTTCTCCTT | ENSG00000162772 | 160 |
| Tropomyosin 1 | TPM1 | GCTGGTTGAGGAAGAGTTGG | TCGCTCTCAATGATGACCAG | ENSG00000140416 | 246 |
| SRY (sex determining region Y) -box 9 | SOX9 | TTGAGCCTTAAAACGGTGCT | CTGGTGTTCTGAGAGGCACA | ENSG00000125398 | 244 |
| Phosphodiesterase 4b, cAMP-specific | PDE4B | GGAAAAATCCCAGGTTGGTT | AGTGGTGGTGAGGGACTTTG | ENSG00000184588 | 159 |
| Interleukin 4 receptor | IL4R | CAAGCTCTTGCCCTGTTTTC | TGCACAGAAGCTCCCTTTTT | ENSG00000077238 | 238 |
| 5-hydroxytryptamine (serotonin) receptor 7, adenylate cyclase-coupled | HTR7 | GAAGAGTGCTGCCAAACACA | GGTGGCTGCTTTCTGTTCTC | ENSG00000148680 | 181 |
| Collagen, type VI, alpha 3 | COL6A3 | ATCTCCTTCATCCCGGACTT | GGACCCATCGATGAGAAAGA | ENSG00000163359 | 192 |
| Interleukin 1, beta | IL1B | GGGCCTCAAGGAAAAGAATC | TTCTGCTTGAGAGGTGCTGA | ENSG00000125538 | 205 |
| Beta-actin | ACTB | AAAGACCTGTACGCCAACAC | CTCAGGAGGAGCAATGATCTTG | ENSG00000075624 | 132 |
| Hypoxanthine-guanine phosphoribosyltransferase | HPRT1 | TGGCGTCGTGATTAGTGATG | ACATCTCGAGCAAGACGTTC | ENSG00000165704 | 141 |
| Glyceraldehyde 3-phosphate dehydrogenase | GAPDH | TCAGCCGCATCTTCTTTTGC | ACGACCAAATCCGTTGACTC | ENSG00000111640 | 90 |
| Gene Symbol | Gene Name | Fold Change | Adj. p. val |
|---|---|---|---|
| KRT19 | Keratin 19 | 47.96 | 0.024 |
| OXTR | Oxytocin receptor | 38.39 | 0.001 |
| CTGF | Connective tissue growth factor | 36.07 | 0.003 |
| NEXN | Nexilin (F actin binding protein) | 35.21 | 0.001 |
| MYLK | Myosin light chain kinase | 27.45 | 0.003 |
| CSRP3 | Cysteine and glycine-rich protein 3 (cardiac LIM protein) | 27.41 | 0.019 |
| CAV1 | Caveolin 1, caveolae protein, 22kDa | 22.41 | 0.002 |
| ACTG2 | Actin, gamma 2, smooth muscle, enteric | 21.30 | 0.024 |
| ATF3 | Activating transcription factor 3 | 19.20 | 0.019 |
| TMP1 | Tropomyosin 1 (alpha) | 17.88 | <0.001 |
| SOX9 | SRY (sex determining region Y)-box 9 | −5.74 | 0.020 |
| WNT5A | Wingless-type MMTV integration site family, member 5A | −6.31 | 0.018 |
| PDE4B | Phosphodiesterase 4B, cAMP-specific | −7.09 | <0.001 |
| IL4R | Interleukin 4 receptor | −8.22 | 0.003 |
| HTR7 | 5-hydroxytryptamine (serotonin) receptor 7, adenylate cyclase-coupled | −8.85 | 0.005 |
| COL6A3 | Collagen, type VI, alpha 3 | −11.20 | 0.018 |
| IL1B | Interleukin 1, beta | −14.49 | 0.027 |
| TGFBR3 | Transforming growth factor, beta receptor III | −20.43 | 0.021 |
| PTGS2 | Prostaglandin-endoperioxide synthase 2 (prostaglandin G/H synthase and cyclooxygenase) | −22.65 | 0.001 |
| EREG | Epiregulin | −98.98 | <0.001 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dompe, C.; Kranc, W.; Jopek, K.; Kowalska, K.; Ciesiółka, S.; Chermuła, B.; Bryja, A.; Jankowski, M.; Perek, J.; Józkowiak, M.; et al. Muscle Cell Morphogenesis, Structure, Development and Differentiation Processes Are Significantly Regulated during Human Ovarian Granulosa Cells In Vitro Cultivation. J. Clin. Med. 2020, 9, 2006. https://doi.org/10.3390/jcm9062006
Dompe C, Kranc W, Jopek K, Kowalska K, Ciesiółka S, Chermuła B, Bryja A, Jankowski M, Perek J, Józkowiak M, et al. Muscle Cell Morphogenesis, Structure, Development and Differentiation Processes Are Significantly Regulated during Human Ovarian Granulosa Cells In Vitro Cultivation. Journal of Clinical Medicine. 2020; 9(6):2006. https://doi.org/10.3390/jcm9062006
Chicago/Turabian StyleDompe, Claudia, Wiesława Kranc, Karol Jopek, Katarzyna Kowalska, Sylwia Ciesiółka, Błażej Chermuła, Artur Bryja, Maurycy Jankowski, Joanna Perek, Małgorzata Józkowiak, and et al. 2020. "Muscle Cell Morphogenesis, Structure, Development and Differentiation Processes Are Significantly Regulated during Human Ovarian Granulosa Cells In Vitro Cultivation" Journal of Clinical Medicine 9, no. 6: 2006. https://doi.org/10.3390/jcm9062006
APA StyleDompe, C., Kranc, W., Jopek, K., Kowalska, K., Ciesiółka, S., Chermuła, B., Bryja, A., Jankowski, M., Perek, J., Józkowiak, M., Moncrieff, L., Hutchings, G., Janowicz, K., Pawelczyk, L., Bruska, M., Petitte, J., Mozdziak, P., Kulus, M., Piotrowska-Kempisty, H., ... Kempisty, B. (2020). Muscle Cell Morphogenesis, Structure, Development and Differentiation Processes Are Significantly Regulated during Human Ovarian Granulosa Cells In Vitro Cultivation. Journal of Clinical Medicine, 9(6), 2006. https://doi.org/10.3390/jcm9062006