Functional Heterogeneity of Mouse and Human Brain OPCs: Relevance for Preclinical Studies in Multiple Sclerosis

 , , , ,
, , , ,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Human Biopsies
2.3. HOG Cells
2.4. Oligodendrocyte Culture
2.5. Preparation of Extracts for Anosmin-1
2.6. Chemotaxis
2.7. Time-Lapse Imaging
2.8. Differentiation Assay
2.9. Pharmacological Treatments
2.10. Differential FGFR Expression
2.11. Proliferation Assays
2.12. 1H HR-MAS NMR Spectroscopy of OPCs
2.13. Imaging Analysis
2.14. Statistical Analysis
3. Results
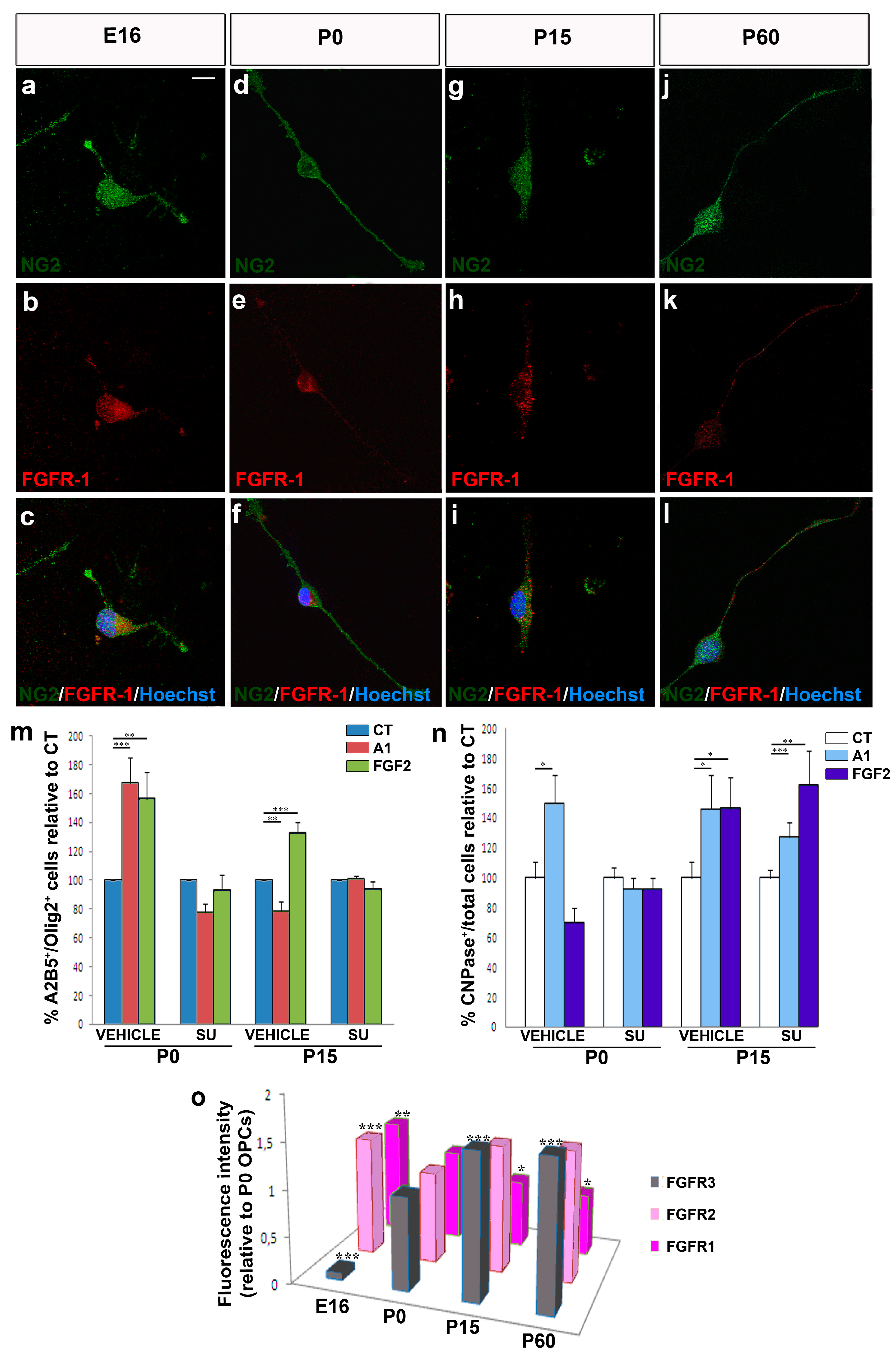
3.1. Changes in the Intrinsic Properties of OPCs with Aging
3.2. Migrating OPCs Respond Dynamically to FGF2 and Anosmin-1 in an Age-Dependent Manner
3.3. Migration of Human OPCs in the Presence of FGF2 and Anosmin-1
3.4. The Biological Effects of FGF2 and Anosmin-1 on OPC Proliferation and Differentiation
3.5. The effects of FGF2 and Anosmin-1 on OPCs are FGFR-Dependent
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CNS | Central Nervous System |
| DMEM | Dulbecco’s Modified Eagle’s Medium |
| ECM | Extracellular Matrix |
| FBS | Fetal Bovine Serum |
| HRMAS | High-Resolution Magic Angle Spinning |
| MS | Multiple Sclerosis |
| OPC | Oligodendrocyte Precursor Cell |
References
- Dawson, M.R.; Polito, A.; Levine, J.M.; Reynolds, R. NG2-expressing glial progenitor cells: An abundant and widespread population of cycling cells in the adult rat CNS. Mol. Cell Neurosci. 2003, 24, 476–488. [Google Scholar] [CrossRef]
- De Castro, F.; Zalc, B. Migration of Myelin-Forming Cells in the CNS. In Cellular Migration and Formation of Neuronal Connections; Rubenstein, J.L.R., Rakic, P., Eds.; Academic Press: Oxford, UK, 2013; Chapter 23; pp. 417–429. [Google Scholar]
- McKenzie, I.A.; Ohayon, D.; Li, H.; de Faria, J.P.; Emery, B.; Tohyama, K.; Richardson, W.D. Motor skill learning requires active central myelination. Science 2014, 346, 318–322. [Google Scholar] [CrossRef] [PubMed]
- Richardson, W.D.; Kessaris, N.; Pringle, N. Oligodendrocyte wars. Nat. Rev. Neurosci. 2006, 7, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Young, K.M.; Psachoulia, K.; Tripathi, R.B.; Dunn, S.J.; Cossell, L.; Attwell, D.; Tohyama, K.; Richardson, W.D. Oligodendrocyte dynamics in the healthy adult CNS: Evidence for myelin remodeling. Neuron 2013, 77, 873–885. [Google Scholar] [CrossRef] [PubMed]
- Dimou, L.; Simon, C.; Kirchhoff, F.; Takebayashi, H.; Götz, M. Progeny of Olig2-expressing progenitors in the gray and white matter of the adult mouse cerebral cortex. J. Neurosci. 2008, 28, 10434–10442. [Google Scholar] [CrossRef]
- Dimou, L.; Gallo, V. NG2-glia and their functions in the central nervous system. Glia 2015, 63, 1429–1451. [Google Scholar] [CrossRef]
- Kang, S.H.; Fukaya, M.; Yang, J.K.; Rothstein, J.D.; Bergles, D.E. NG2+ CNS glial progenitors remain committed to the oligodendrocyte lineage in postnatal life and following neurodegeneration. Neuron 2010, 68, 668–681. [Google Scholar] [CrossRef]
- Psachoulia, K.; Jamen, F.; Young, K.M.; Richardson, W.D. Cell cycle dynamics of NG2 cells in the postnatal and ageing brain. Neuron Glia Biol. 2009, 5, 57–67. [Google Scholar] [CrossRef]
- Rivers, L.E.; Young, K.M.; Rizzi, M.; Jamen, F.; Psachoulia, K.; Wade, A.; Kessaris, N.; Richardson, W.D. PDGFRA/NG2 glia generate myelinating oligodendrocytes and piriform projection neurons in adult mice. Nat. Neurosci. 2008, 11, 1392–1401. [Google Scholar] [CrossRef]
- Zhu, Q.; Whittemore, S.R.; Devries, W.H.; Zhao, X.; Kuypers, N.J.; Qiu, M. Dorsally-derived oligodendrocytes in the spinal cord contribute to axonal myelination during development and remyelination following focal demyelination. Glia 2011, 59, 1612–1621. [Google Scholar] [CrossRef]
- Murcia-Belmonte, V.; Esteban, P.F.; Martínez-Hernández, J.; Gruart, A.; Luján, R.; Delgado-García, J.M.; de Castro, F. Anosmin-1 over-expression regulates oligodendrocyte precursor cell proliferation, migration and myelin sheath thickness. Brain Struct. Funct. 2016, 221, 1365–1385. [Google Scholar] [CrossRef] [PubMed]
- Hill, R.A.; Nishiyama, A. NG2 cells (polydendrocytes): Listeners to the neural network with diverse properties. Glia 2014, 62, 1195–1210. [Google Scholar] [CrossRef] [PubMed]
- Richardson, W.D.; Young, K.M.; Tripathi, R.B.; McKenzie, I. NG2-glia as multipotent neural stem cells: Fact or fantasy? Neuron 2011, 70, 661–673. [Google Scholar] [CrossRef] [PubMed]
- Calver, A.R.; Hall, A.C.; Yu, W.P.; Walsh, F.S.; Heath, J.K.; Betsholtz, C.; Richardson, W.D. Oligodendrocyte population dynamics and the role of PDGF in vivo. Neuron 1998, 20, 869–882. [Google Scholar] [CrossRef]
- Simon, C.; Götz, M.; Dimou, L. Progenitors in the adult cerebral cortex: Cell cycle properties and regulation by physiological stimuli and injury. Glia 2011, 59, 869–881. [Google Scholar] [CrossRef]
- Nishiyama, A.; Komitova, M.; Suzuki, R.; Zhu, X. Polydendrocytes (NG2 cells): Multifunctional cells with lineage plasticity. Nat. Rev. Neurosci. 2009, 10, 9–22. [Google Scholar] [CrossRef]
- Seo, J.H.; Miyamoto, N.; Hayakawa, K.; Pham, L.D.; Maki, T.; Ayata, C.; Kim, K.W.; Lo, E.H.; Arai, K. Oligodendrocyte precursors induce early blood-brain barrier opening after white matter injury. J. Clin. Invest. 2013, 123, 782–786. [Google Scholar] [CrossRef]
- Pepper, R.; Cullen, L.C.; Pitman, K.A.; Young, K.M. How do cells of the oligodendrocyte lineage affect neuronal circuits to influence motor function, memory and mood? Front. Cell Neurosci. 2018, 12, 399. [Google Scholar] [CrossRef]
- Boyd, A.; Zhang, H.; Williams, A. Insufficient OPC migration into demyelinated lesions is a cause of poor remyelination in MS and mouse models. Acta Neuropathol. 2013, 125, 841–859. [Google Scholar] [CrossRef]
- Clemente, D.; Ortega, M.C.; Arenzana, F.J.; de Castro, F. FGF-2 and Anosmin-1 are selectively expressed in different types of multiple sclerosis lesions. J. Neurosci. 2011, 31, 14899–14909. [Google Scholar] [CrossRef]
- Ferent, J.; Zimmer, C.; Durbec, P.; Ruat, M.; Traiffort, E. Sonic Hedgehog signaling is a positive oligodendrocyte regulator during demyelination. J. Neurosci. 2013, 33, 1759–1772. [Google Scholar] [CrossRef] [PubMed]
- Moyon, S.; Dubessy, A.L.; Aigrot, M.S.; Trotter, M.; Huang, J.K.; Dauphinot, L.; Potier, M.C.; Kerninon, C.; Melik Parsadaniantz, S.; Franklin, R.J.; et al. Demyelination causes adult CNS progenitors to revert to an immature state and express immune cues that support their migration. J. Neurosci. 2015, 35, 4–20. [Google Scholar] [CrossRef]
- Ortega, J.A.; Radonjic, N.V.; Zecevic, N. Sonic hedgehog promotes generation and maintenance of human forebrain Olig2 progenitors. Front. Cell Neurosci. 2013, 7, 254. [Google Scholar] [CrossRef] [PubMed]
- Piaton, G.; Williams, A.; Seilhean, D.; Lubetzki, C. Remyelination in multiple sclerosis. Prog. Brain Res. 2009, 175, 453–464. [Google Scholar] [PubMed]
- Williams, A.; Piaton, G.; Aigrot, M.S.; Belhadi, A.; Theaudin, M.; Petermann, F.; Thomas, J.L.; Zalc, B.; Lubetzki, C. Semaphorin 3A and 3F: Key players in myelin repair in multiple sclerosis? Brain 2007, 130 Pt 10, 2554–2565. [Google Scholar] [CrossRef]
- Diaz-Balzac, C.A.; Lazaro-Pena, M.I.; Ramos-Ortiz, G.A.; Bulow, H.E. The Adhesion Molecule KAL-1/anosmin-1 Regulates Neurite Branching through a SAX-7/L1CAM-EGL-15/FGFR Receptor Complex. Cell Rep. 2015, 11, 1377–1384. [Google Scholar] [CrossRef]
- Azin, M.; Mirnajafi-Zadeh, J.; Javan, M. Fibroblast Growth Factor-2 Enhanced The Recruitment of Progenitor Cells and Myelin Repair in Experimental Demyelination of Rat Hippocampal Formations. Cell J. 2015, 17, 540. [Google Scholar] [CrossRef]
- Bribián, A.; Barallobre, M.J.; Soussi-Yanicostas, N.; de Castro, F. Anosmin-1 modulates the FGF-2-dependent migration of oligodendrocyte precursors in the developing optic nerve. Mol. Cell Neurosci. 2006, 33, 2–14. [Google Scholar] [CrossRef]
- Bribián, A.; Esteban, P.F.; Clemente, D.; Soussi-Yanicostas, N.; Thomas, J.L.; Zalc, B.; de Castro, F. A novel role for anosmin-1 in the adhesion and migration of oligodendrocyte precursors. Dev. Neurobiol. 2008, 68, 1503–1516. [Google Scholar] [CrossRef]
- Endo, Y.; Ishiwata-Endo, H.; Yamada, K.M. Extracellular matrix protein anosmin promotes neural crest formation and regulates FGF, BMP, and WNT activities. Dev. Cell 2012, 23, 305–316. [Google Scholar] [CrossRef]
- Hu, Y.; Poopalasundaram, S.; Graham, A.; Bouloux, P.M. GnRH neuronal migration and olfactory bulb neurite outgrowth are dependent on FGF receptor 1 signaling, specifically via the PI3K p110alpha isoform in chick embryo. Endocrinology 2013, 154, 388–399. [Google Scholar] [CrossRef] [PubMed]
- Mierzwa, A.J.; Zhou, Y.X.; Hibbits, N.; Vana, A.C.; Armstrong, R.C. FGF2 and FGFR1 signaling regulate functional recovery following cuprizone demyelination. Neurosci. Lett. 2013, 548, 280–285. [Google Scholar] [CrossRef] [PubMed]
- Murcia-Belmonte, V.; Medina-Rodríguez, E.M.; Bribián, A.; de Castro, F.; Esteban, P.F. ERK1/2 signaling is essential for the chemoattraction exerted by human FGF2 and human anosmin-1 on newborn rat and mouse OPCs via FGFR1. Glia 2014, 62, 374–386. [Google Scholar] [CrossRef]
- Mohan, H.; Friese, A.; Albrecht, S.; Krumbholz, M.; Elliott, C.L.; Arthur, A.; Menon, R.; Farina, C.; Junker, A.; Stadelmann, C.; et al. Transcript profiling of different types of multiple sclerosis lesions yields FGF1 as a promoter of remyelination. Acta Neuropathol. Commun. 2014, 2, 168. [Google Scholar] [CrossRef]
- Furusho, M.; Roulois, A.J.; Franklin, R.J.; Bansal, R. Fibroblast growth factor signaling in oligodendrocyte-lineage cells facilitates recovery of chronically demyelinated lesions but is redundant in acute lesions. Glia 2015, 63, 1714–1728. [Google Scholar] [CrossRef]
- Medina-Rodríguez, E.M.; Arenzana, F.J.; Bribián, A.; de Castro, F. Protocol to isolate a large amount of functional oligodendrocyte precursor cells from the cerebral cortex of adult mice and humans. PLoS ONE 2013, 8, e81620. [Google Scholar] [CrossRef]
- Bin, J.M.; Rajasekharan, S.; Kuhlmann, T.; Hanes, I.; Marcal, N.; Han, D.; Rodrigues, S.P.; Leong, S.Y.; Newcombe, J.; Antel, J.P.; et al. Full-length and fragmented netrin-1 in multiple sclerosis plaques are inhibitors of oligodendrocyte precursor cell migration. Am. J. Pathol. 2013, 183, 673–680. [Google Scholar] [CrossRef]
- Cui, Q.L.; Kuhlmann, T.; Miron, V.E.; Leong, S.Y.; Fang, J.; Gris, P.; Kennedy, T.E.; Almazan, G.; Antel, J. Oligodendrocyte progenitor cell susceptibility to injury in multiple sclerosis. Am. J. Pathol. 2013, 183, 516–525. [Google Scholar] [CrossRef]
- John, G.R. Investigation of astrocyte—Oligodendrocyte interactions in human cultures. Methods Mol. Biol. 2012, 814, 401–414. [Google Scholar]
- Medina-Rodriguez, E.M.; Arenzana, F.J.; Pastor, J.; Redondo, M.; Palomo, V.; Garcia de Sola, R.; Gil, C.; Martinez, A.; Bribian, A.; de Castro, F. Inhibition of endogenous phosphodiesterase 7 promotes oligodendrocyte precursor differentiation and survival. Cell Mol. Life Sci. 2013, 70, 3449–3462. [Google Scholar] [CrossRef]
- Soussi-Yanicostas, N.; Hardelin, J.P.; Arroyo-Jimenez, M.M.; Ardouin, O.; Legouis, R.; Levilliers, J.; Traincard, F.; Betton, J.M.; Cabanie, L.; Petit, C. Initial characterization of anosmin-1, a putative extracellular matrix protein synthesized by definite neuronal cell populations in the central nervous system. J. Cell Sci. 1996, 109 Pt 7, 1749–1757. [Google Scholar]
- García-González, D.; Clemente, D.; Coelho, M.; Esteban, P.F.; Soussi-Yanicostas, N.; de Castro, F. Dynamic roles of FGF-2 and Anosmin-1 in the migration of neuronal precursors from the subventricular zone during pre- and postnatal development. Exp. Neurol. 2010, 222, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Casaccia-Bonnefil, P.; Tikoo, R.; Kiyokawa, H.; Friedrich, V., Jr.; Chao, M.V.; Koff, A. Oligodendrocyte precursor differentiation is perturbed in the absence of the cyclin-dependent kinase inhibitor p27Kip1. Genes Dev. 1997, 11, 2335–2346. [Google Scholar] [CrossRef] [PubMed]
- Esteban, P.F.; Murcia-Belmonte, V.; García-González, D.; de Castro, F. The cysteine-rich region and the whey acidic protein domain are essential for anosmin-1 biological functions. J. Neurochem. 2013, 124, 708–720. [Google Scholar] [CrossRef]
- Murcia-Belmonte, V.; Esteban, P.F.; García-González, D.; De Castro, F. Biochemical dissection of Anosmin-1 interaction with FGFR1 and components of the extracellular matrix. J. Neurochem. 2010, 115, 1256–1265. [Google Scholar] [CrossRef]
- García-González, D.; Murcia-Belmonte, V.; Esteban, P.F.; Ortega, F.; Díaz, D.; Sánchez-Vera, I.; Lebron-Galán, R.; Escobar-Castanondo, L.; Martínez-Millán, L.; Weruaga, E.; et al. Anosmin-1 over-expression increases adult neurogenesis in the subventricular zone and neuroblast migration to the olfactory bulb. Brain Struct. Funct. 2016, 221, 239–260. [Google Scholar] [CrossRef]
- Rocha, C.M.; Barros, A.S.; Gil, A.M.; Goodfellow, B.J.; Humpfer, E.; Spraul, M.; Carreira, I.M.; Melo, J.B.; Bernardo, J.; Gomes, A.; et al. Metabolic profiling of human lung cancer tissue by 1H high resolution magic angle spinning (HRMAS) NMR spectroscopy. J. Proteome Res. 2010, 9, 319–332. [Google Scholar] [CrossRef]
- Moestue, S.; Sitter, B.; Bathen, T.F.; Tessem, M.B.; Gribbestad, I.S. HR MAS MR spectroscopy in metabolic characterization of cancer. Curr. Top. Med. Chem. 2011, 11, 2–26. [Google Scholar] [CrossRef]
- Govindaraju, V.; Young, K.; Maudsley, A.A. Proton NMR chemical shifts and coupling constants for brain metabolites. NMR Biomed. 2000, 13, 129–153. [Google Scholar] [CrossRef]
- Flores, A.I.; Narayanan, S.P.; Morse, E.N.; Shick, H.E.; Yin, X.; Kidd, G.; Avila, R.L.; Kirschner, D.A.; Macklin, W.B. Constitutively active Akt induces enhanced myelination in the CNS. J. Neurosci. 2008, 28, 7174–7183. [Google Scholar] [CrossRef]
- Lu, Z.; Ku, L.; Chen, Y.; Feng, Y. Developmental abnormalities of myelin basic protein expression in fyn knock-out brain reveal a role of Fyn in posttranscriptional regulation. J. Biol. Chem. 2005, 280, 389–395. [Google Scholar] [CrossRef]
- González-Martínez, D.; Kim, S.H.; Hu, Y.; Guimond, S.; Schofield, J.; Winyard, P.; Vannelli, G.B.; Turnbull, J.; Bouloux, P.M. Anosmin-1 modulates fibroblast growth factor receptor 1 signaling in human gonadotropin-releasing hormone olfactory neuroblasts through a heparan sulfate-dependent mechanism. J. Neurosci. 2004, 24, 10384–10392. [Google Scholar] [CrossRef]
- Hu, Y.; Guimond, S.E.; Travers, P.; Cadman, S.; Hohenester, E.; Turnbull, J.E.; Kim, S.H.; Bouloux, P.M. Novel mechanisms of fibroblast growth factor receptor 1 regulation by extracellular matrix protein anosmin-1. J. Biol. Chem. 2009, 284, 29905–29920. [Google Scholar] [CrossRef]
- Bansal, R.; Pfeiffer, S.E. FGF-2 converts mature oligodendrocytes to a novel phenotype. J. Neurosci. Res. 1997, 50, 215–228. [Google Scholar] [CrossRef]
- Bansal, R. Fibroblast growth factors and their receptors in oligodendrocyte development: Implications for demyelination and remyelination. Dev. Neurosci. 2002, 24, 35–46. [Google Scholar] [CrossRef]
- Oh, L.Y.; Yong, V.W. Astrocytes promote process outgrowth by adult human oligodendrocytes in vitro through interaction between bFGF and astrocyte extracellular matrix. Glia 1996, 17, 237–253. [Google Scholar] [CrossRef]
- Bansal, R.; Kumar, M.; Murray, K.; Morrison, R.S.; Pfeiffer, S.E. Regulation of FGF receptors in the oligodendrocyte lineage. Mol. Cell Neurosci. 1996, 7, 263–275. [Google Scholar] [CrossRef]
- Fortin, D.; Rom, E.; Sun, H.; Yayon, A.; Bansal, R. Distinct fibroblast growth factor (FGF)/FGF receptor signaling pairs initiate diverse cellular responses in the oligodendrocyte lineage. J. Neurosci. 2005, 25, 7470–7479. [Google Scholar] [CrossRef]
- Furusho, M.; Dupree, J.L.; Nave, K.A.; Bansal, R. Fibroblast growth factor receptor signaling in oligodendrocytes regulates myelin sheath thickness. J. Neurosci. 2012, 32, 6631–6641. [Google Scholar] [CrossRef]
- Franklin, R.J.; Gallo, V. The translational biology of remyelination: Past, present, and future. Glia 2014, 62, 1905–1915. [Google Scholar] [CrossRef]
- Maki, T.; Liang, A.C.; Miyamoto, N.; Lo, E.H.; Arai, K. Mechanisms of oligodendrocyte regeneration from ventricular-subventricular zone-derived progenitor cells in white matter diseases. Front. Cell Neurosci. 2013, 7, 275. [Google Scholar] [CrossRef]
- Nishiyama, A. Polydendrocytes: NG2 cells with many roles in development and repair of the CNS. Neuroscientist 2007, 13, 62–76. [Google Scholar] [CrossRef]
- Tepavcevic, V.; Kerninon, C.; Aigrot, M.S.; Meppiel, E.; Mozafari, S.; Arnould-Laurent, R.; Ravassard, P.; Kennedy, T.E.; Nait-Oumesmar, B.; Lubetzki, C. Early netrin-1 expression impairs central nervous system remyelination. Ann. Neurol. 2014, 76, 252–268. [Google Scholar] [CrossRef]
- Syed, Y.A.; Baer, A.; Hofer, M.P.; González, G.A.; Rundle, J.; Myrta, S.; Huang, J.K.; Zhao, C.; Rossner, M.J.; Trotter, M.W.; et al. Inhibition of phosphodiesterase-4 promotes oligodendrocyte precursor cell differentiation and enhances CNS remyelination. EMBO Mol. Med. 2013, 5, 1918–1934. [Google Scholar] [CrossRef]
- De Castro, F.; Bribián, A.; Ortega, M.C. Regulation of oligodendrocyte precursor migration during development, in adulthood and in pathology. Cell Mol. Life Sci. 2013, 70, 4355–4368. [Google Scholar] [CrossRef]
- Huang, J.K.; Franklin, R.J. Regenerative medicine in multiple sclerosis: Identifying pharmacological targets of adult neural stem cell differentiation. Neurochem. Int. 2011, 59, 329–332. [Google Scholar] [CrossRef]
- Azim, K.; Raineteau, O.; Butt, A.M. Intraventricular injection of FGF-2 promotes generation of oligodendrocyte-lineage cells in the postnatal and adult forebrain. Glia 2012, 60, 1977–1990. [Google Scholar] [CrossRef]
- Murtie, J.C.; Zhou, Y.X.; Le, T.Q.; Vana, A.C.; Armstrong, R.C. PDGF and FGF2 pathways regulate distinct oligodendrocyte lineage responses in experimental demyelination with spontaneous remyelination. Neurobiol. Dis. 2005, 19, 171–182. [Google Scholar] [CrossRef] [PubMed]
- McKinnon, R.D.; Smith, C.; Behar, T.; Smith, T.; Dubois-Dalcq, M. Distinct effects of bFGF and PDGF on oligodendrocyte progenitor cells. Glia 1993, 7, 245–254. [Google Scholar] [CrossRef]
- Jansen, J.F.; Shamblott, M.J.; van Zijl, P.C.; Lehtimaki, K.K.; Bulte, J.W.; Gearhart, J.D.; Hakumaki, J.M. Stem cell profiling by nuclear magnetic resonance spectroscopy. Magn. Reson. Med. 2006, 56, 666–670. [Google Scholar] [CrossRef]
- Urenjak, J.; Williams, S.R.; Gadian, D.G.; Noble, M. Proton nuclear magnetic resonance spectroscopy unambiguously identifies different neural cell types. J. Neurosci. 1993, 13, 981–989. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Fodor, W.L.; Machaty, Z. Dynamics of lamin A/C in porcine embryos produced by nuclear transfer. Mol. Reprod. Dev. 2007, 74, 1221–1227. [Google Scholar] [CrossRef] [PubMed]
- Maniotis, A.J.; Chen, C.S.; Ingber, D.E. Demonstration of mechanical connections between integrins, cytoskeletal filaments, and nucleoplasm that stabilize nuclear structure. Proc. Natl. Acad. Sci. USA 1997, 94, 849–854. [Google Scholar] [CrossRef] [PubMed]
- Pollard, T.D.; Borisy, G.G. Cellular motility driven by assembly and disassembly of actin filaments. Cell 2003, 112, 453–465. [Google Scholar] [CrossRef]
- Cariboni, A.; Pimpinelli, F.; Colamarino, S.; Zaninetti, R.; Piccolella, M.; Rumio, C.; Piva, F.; Rugarli, E.I.; Maggi, R. The product of X-linked Kallmann’s syndrome gene (KAL1) affects the migratory activity of gonadotropin-releasing hormone (GnRH)-producing neurons. Hum. Mol. Genet. 2004, 13, 2781–2791. [Google Scholar] [CrossRef]
- Choy, C.T.; Kim, H.; Lee, J.Y.; Williams, D.M.; Palethorpe, D.; Fellows, G.; Wright, A.J.; Laing, K.; Bridges, L.R.; Howe, F.A.; et al. Anosmin-1 contributes to brain tumor malignancy through integrin signal pathways. Endocr. Relat. Cancer 2014, 21, 85–99. [Google Scholar] [CrossRef]
- Jian, B.; Nagineni, C.N.; Meleth, S.; Grizzle, W.; Bland, K.; Chaudry, I.; Raju, R. Anosmin-1 involved in neuronal cell migration is hypoxia inducible and cancer regulated. Cell Cycle. 2009, 8, 3770–3776. [Google Scholar] [CrossRef]
- Kanda, M.; Shimizu, D.; Fujii, T.; Sueoka, S.; Tanaka, Y.; Ezaka, K.; Takami, H.; Tanaka, H.; Hashimoto, R.; Iwata, N.; et al. Function and diagnostic value of Anosmin-1 in gastric cancer progression. Int. J. Cancer 2016, 138, 721–730. [Google Scholar] [CrossRef]
- De Castro, F.; Seal, R.; Maggi, R.; Group of HcfKALn. ANOS1: A unified nomenclature for Kallmann syndrome 1 gene (KAL1) and anosmin-1. Brief. Funct. Genom. 2017, 16, 205–210. [Google Scholar] [CrossRef]
- Liu, J.; Cao, W.; Chen, W.; Xu, L.; Zhang, C. Decreased expression of Kallmann syndrome 1 sequence gene (KAL1) contributes to oral squamous cell carcinoma progression and significantly correlates with poorly differentiated grade. J. Oral Pathol. Med. 2015, 44, 109–114. [Google Scholar] [CrossRef]
- Nishiyama, A.; Suzuki, R.; Zhu, X. NG2 cells (polydendrocytes) in brain physiology and repair. Front. Neurosci. 2014, 8, 133. [Google Scholar] [CrossRef] [PubMed]
- Spitzer, S.O.; Sitnikov, S.; Kamen, Y.; Evans, K.A.; Kronenberg-Versteeg, D.; Dietmann, S.; de Faria, O., Jr.; Agathou, S.; Karadottir, R.T. Oligodendrocyte Progenitor Cells Become Regionally Diverse and Heterogeneous with Age. Neuron 2019, 101, 459–471.e455. [Google Scholar] [CrossRef]
- Dubois-Dalcq, M.; Ffrench-Constant, C.; Franklin, R.J. Enhancing central nervous system remyelination in multiple sclerosis. Neuron 2005, 48, 9–12. [Google Scholar] [CrossRef]
- Leong, S.Y.; Rao, V.T.; Bin, J.M.; Gris, P.; Sangaralingam, M.; Kennedy, T.E.; Antel, J.P. Heterogeneity of oligodendrocyte progenitor cells in adult human brain. Ann. Clin. Transl. Neurol. 2014, 1, 272–283. [Google Scholar] [CrossRef]
- Mitew, S.; Hay, C.M.; Peckham, H.; Xiao, J.; Koenning, M.; Emery, B. Mechanisms regulating the development of oligodendrocytes and central nervous system myelin. Neuroscience 2014, 276, 29–47. [Google Scholar] [CrossRef]
- Medina-Rodríguez, E.M.; Bribián, A.; Boyd, A.; Palomo, V.; Pastor, J.; Lagares, A.; Gil, C.; Martínez, A.; Williams, A.; de Castro, F. Promoting in vivo remyelination with small molecules: A neuroreparative pharmacological treatment for Multiple Sclerosis. Sci. Rep. 2017, 7, 43545. [Google Scholar] [CrossRef]
- De Castro, F.; Josa-Prado, F. CHAPTER 10 Regulation of Oligodendrocyte Differentiation: New Targets for Drug Discovery in Remyelination. In Emerging Drugs and Targets for Multiple Sclerosis; The Royal Society of Chemistry: London, UK, 2019; pp. 222–240. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient | Age (Years)/Sex | Diagnostic | Lobe | Sample Weight (gr) |
|---|---|---|---|---|
| 540 | 51/Female | Epilepsy | Temporal | 4, 9 |
| 573 | 24/Male | Epilepsy | Temporal | 2, 9 |
| T-590 | 47/Female | Tumoral | Temporal | 1, 9 |
| 554 | 50/Female | Epilepsy | Temporal | 2, 6 |
| 574 | 45/Female | Epilepsy | Temporal | 1, 4 |
| 559 | 45/Female | Epilepsy | Temporal | 1, 1 |
| 579 | 50/Male | Epilepsy | Temporal | 2, 6 |
| 603 | 53/Female | Epilepsy | Temporal | 1, 1 |
| 549 | 32/Female | Epilepsy | Temporal | 2 |
| 641 | 27/Female | Epilepsy | Temporal | 1, 3 |
| T-604 | 37/Female | Tumoral | Temporal | 0, 7 |
| 363 | 51/Female | Epilepsy | Temporal | 4 |
| 583 | 40/Male | Epilepsy | Temporal | 0, 8 |
| T-1-SC | 62/Female | Tumoral | Temporal | 0, 2 |
| T-2-SC | 68/Male | Tumoral | Temporal | 0, 6 |
| T-3-SC | 61/Female | Tumoral | Temporal | 1, 5 |
| T-5-SC | 51/Female | Tumoral | Temporal | 3, 3 |
| Metabolite | δ (ppm), Multiplicity, Number H |
|---|---|
| myo-inositol | 3.93, m |
| myo-inositol | 3.77, m |
| glycine | 3.54, s 2H |
| taurine | 3.42, t, 2H |
| hipotaurine | 3.30, t, 2H |
| total-choline | 3.23, s, 9H |
| glutamine | 2.44, m, 2H |
| glutamate | 2.34, m, 2H |
| glutamine/glutamate/N AA | 2.12 |
| acetate | 1.91, s, 3H |
| lipids | 1.70 |
| alanine | 1.47, d, 3H |
| lactate and lipids (methylene) | 1.31 |
| lipids and aminoacids (methyl) | 0.94, t, 3H |
| CONDITION Number of Stops/Number of Analyzed Cells | |||
|---|---|---|---|
| Species and Age | CT | FGF2 | Anosmin-1 |
| Mouse P0 | 6.2 | 3.5 | 0.85 |
| Mouse P15 | 11.4 | 1.52 | 20.7 |
| Mouse P60 | 3.76 | 2.63 | 8.42 |
| haOPCs, adult | 3.61 | 8.8 | 17.6 |
| malOPCs, adult | 23.7 | 19.7 | 23.1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bribián, A.; Medina-Rodríguez, E.M.; Josa-Prado, F.; García-Álvarez, I.; Machín-Díaz, I.; Esteban, P.F.; Murcia-Belmonte, V.; Vega-Zelaya, L.; Pastor, J.; Garrido, L.; et al. Functional Heterogeneity of Mouse and Human Brain OPCs: Relevance for Preclinical Studies in Multiple Sclerosis. J. Clin. Med. 2020, 9, 1681. https://doi.org/10.3390/jcm9061681
Bribián A, Medina-Rodríguez EM, Josa-Prado F, García-Álvarez I, Machín-Díaz I, Esteban PF, Murcia-Belmonte V, Vega-Zelaya L, Pastor J, Garrido L, et al. Functional Heterogeneity of Mouse and Human Brain OPCs: Relevance for Preclinical Studies in Multiple Sclerosis. Journal of Clinical Medicine. 2020; 9(6):1681. https://doi.org/10.3390/jcm9061681
Chicago/Turabian StyleBribián, Ana, Eva M. Medina-Rodríguez, Fernando Josa-Prado, Isabel García-Álvarez, Isabel Machín-Díaz, Pedro F. Esteban, Verónica Murcia-Belmonte, Lorena Vega-Zelaya, Jesús Pastor, Leoncio Garrido, and et al. 2020. "Functional Heterogeneity of Mouse and Human Brain OPCs: Relevance for Preclinical Studies in Multiple Sclerosis" Journal of Clinical Medicine 9, no. 6: 1681. https://doi.org/10.3390/jcm9061681
APA StyleBribián, A., Medina-Rodríguez, E. M., Josa-Prado, F., García-Álvarez, I., Machín-Díaz, I., Esteban, P. F., Murcia-Belmonte, V., Vega-Zelaya, L., Pastor, J., Garrido, L., & de Castro, F. (2020). Functional Heterogeneity of Mouse and Human Brain OPCs: Relevance for Preclinical Studies in Multiple Sclerosis. Journal of Clinical Medicine, 9(6), 1681. https://doi.org/10.3390/jcm9061681