Systematic Multiomic Analysis of Ly75 Gene Expression and Its Prognostic Value through the Infiltration of Natural Killer (NK) Cells in Skin Cutaneous Melanoma


Abstract
1. Introduction
2. Experimental Section
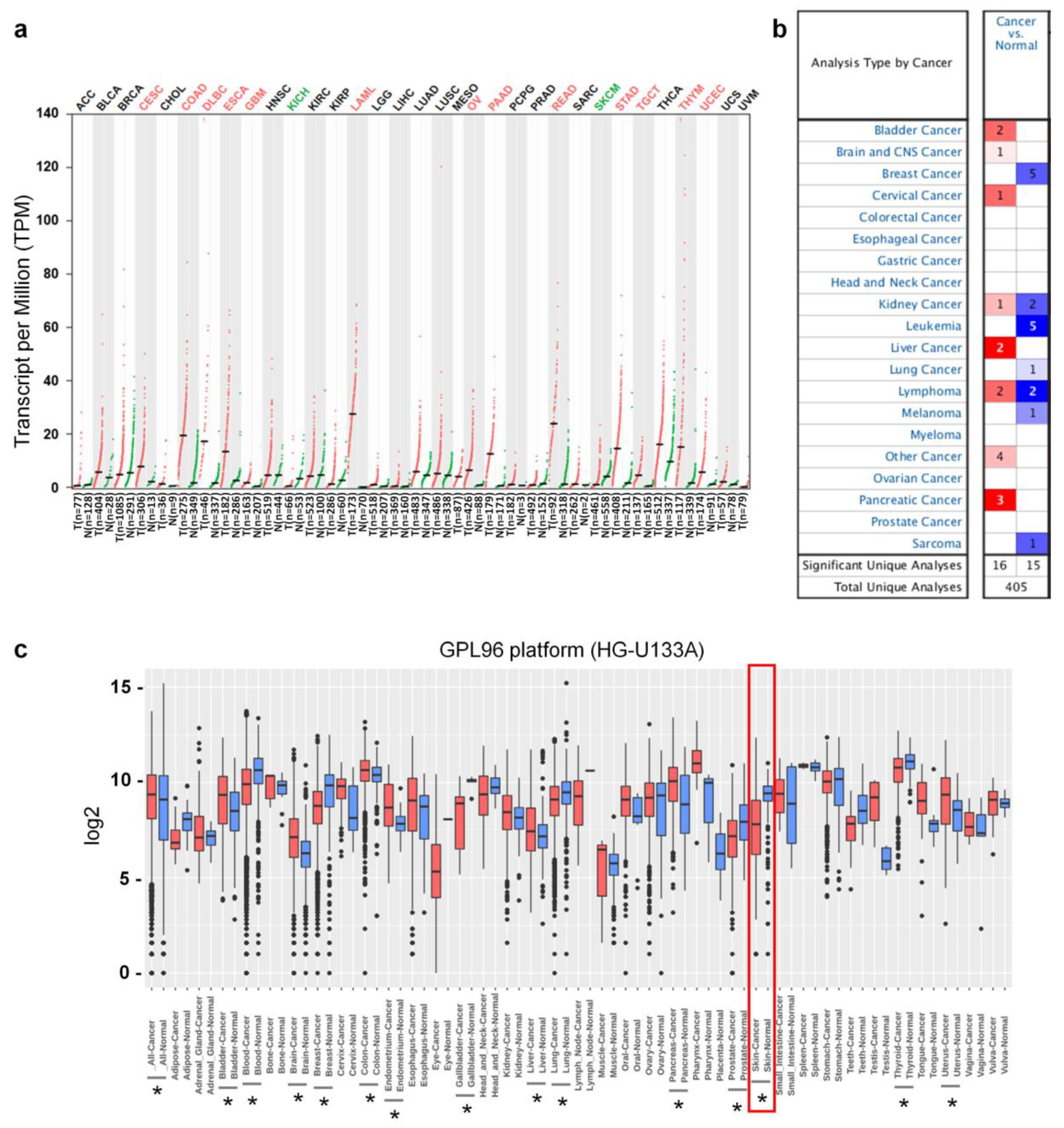
2.1. Ly75 mRNA Expression and Genome Alteration in Cancers
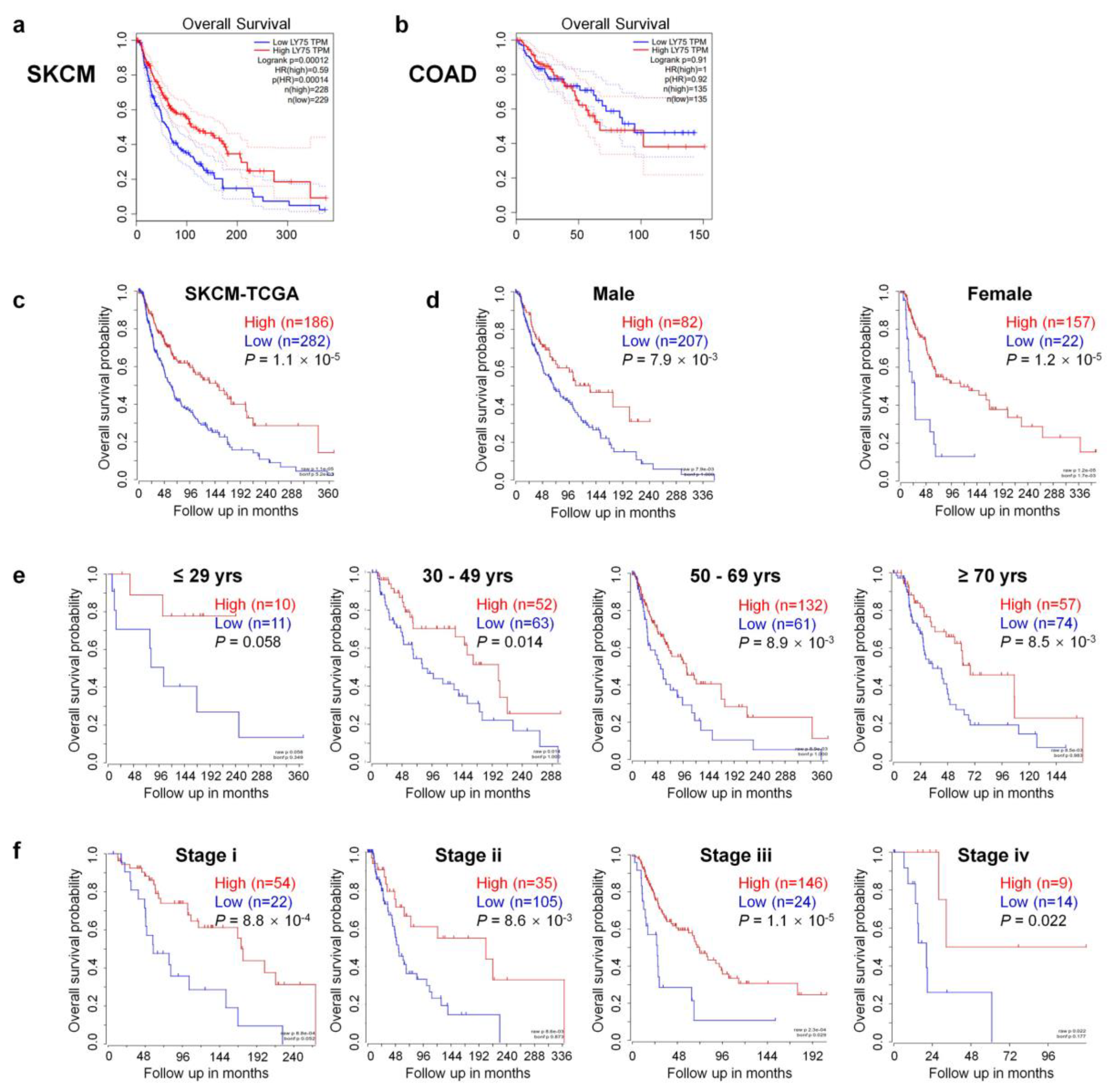
2.2. Prognostic Value of Ly75 Expression in Various Tumors
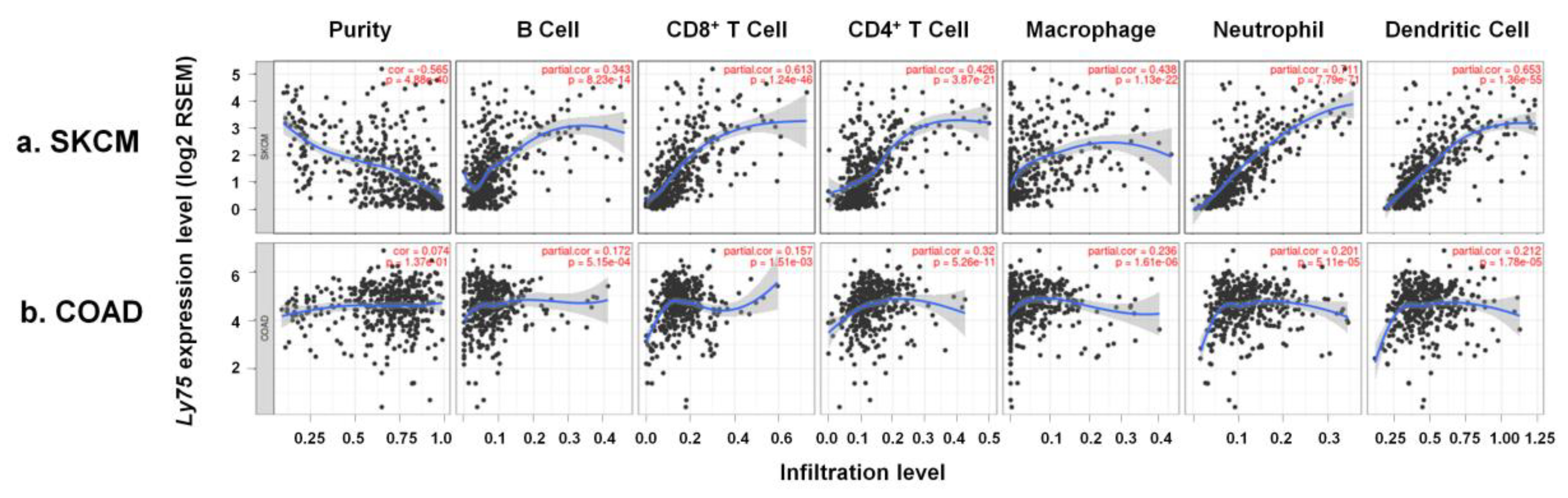
2.3. Analysis of the Association of Ly75 Expression with Immune Infiltration
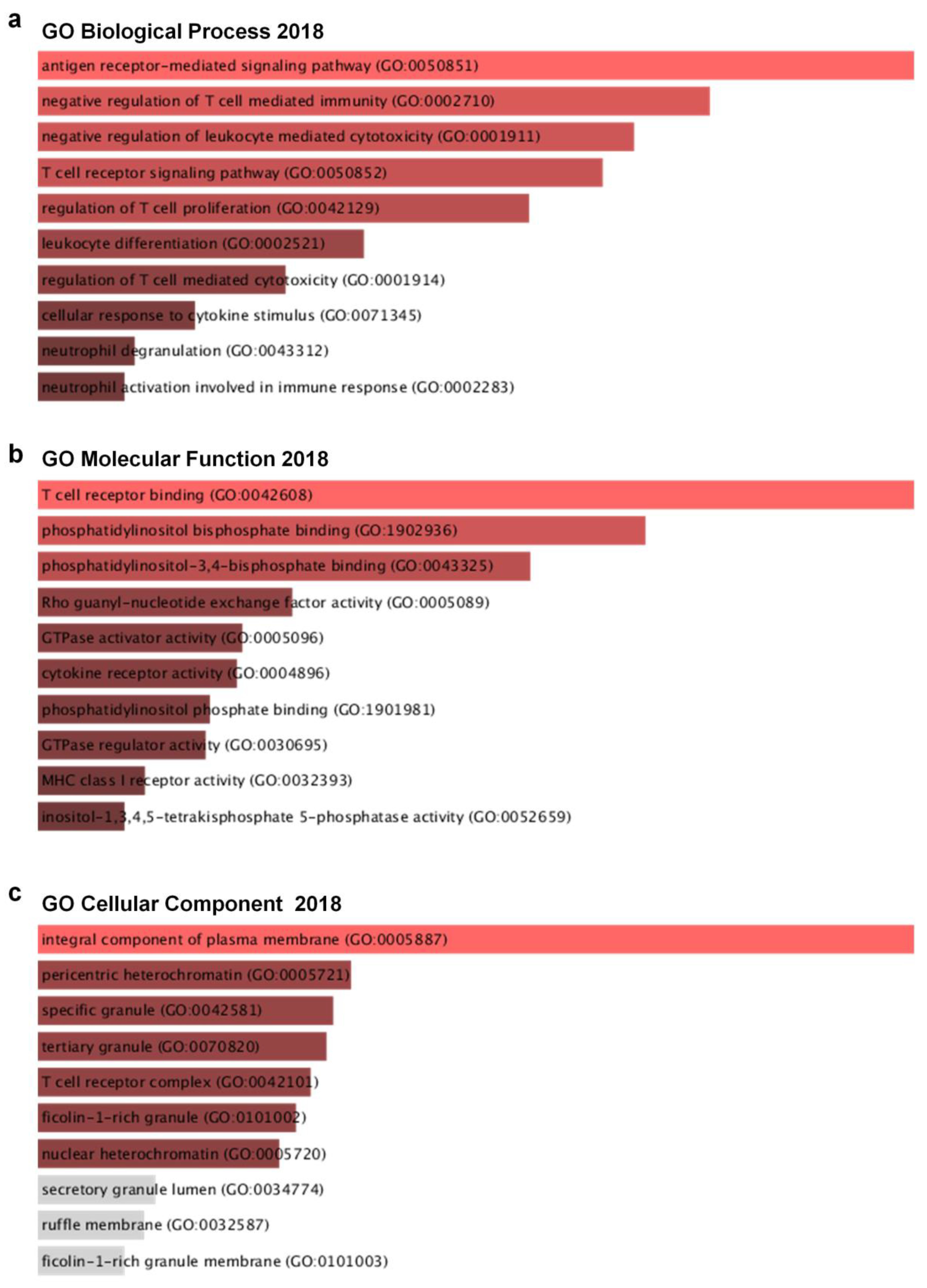
2.4. Profiling and Ontology Analysis of Co-Expressed Genes with Ly75
2.5. Reproducibility of Data Presented from Web Tools
3. Results
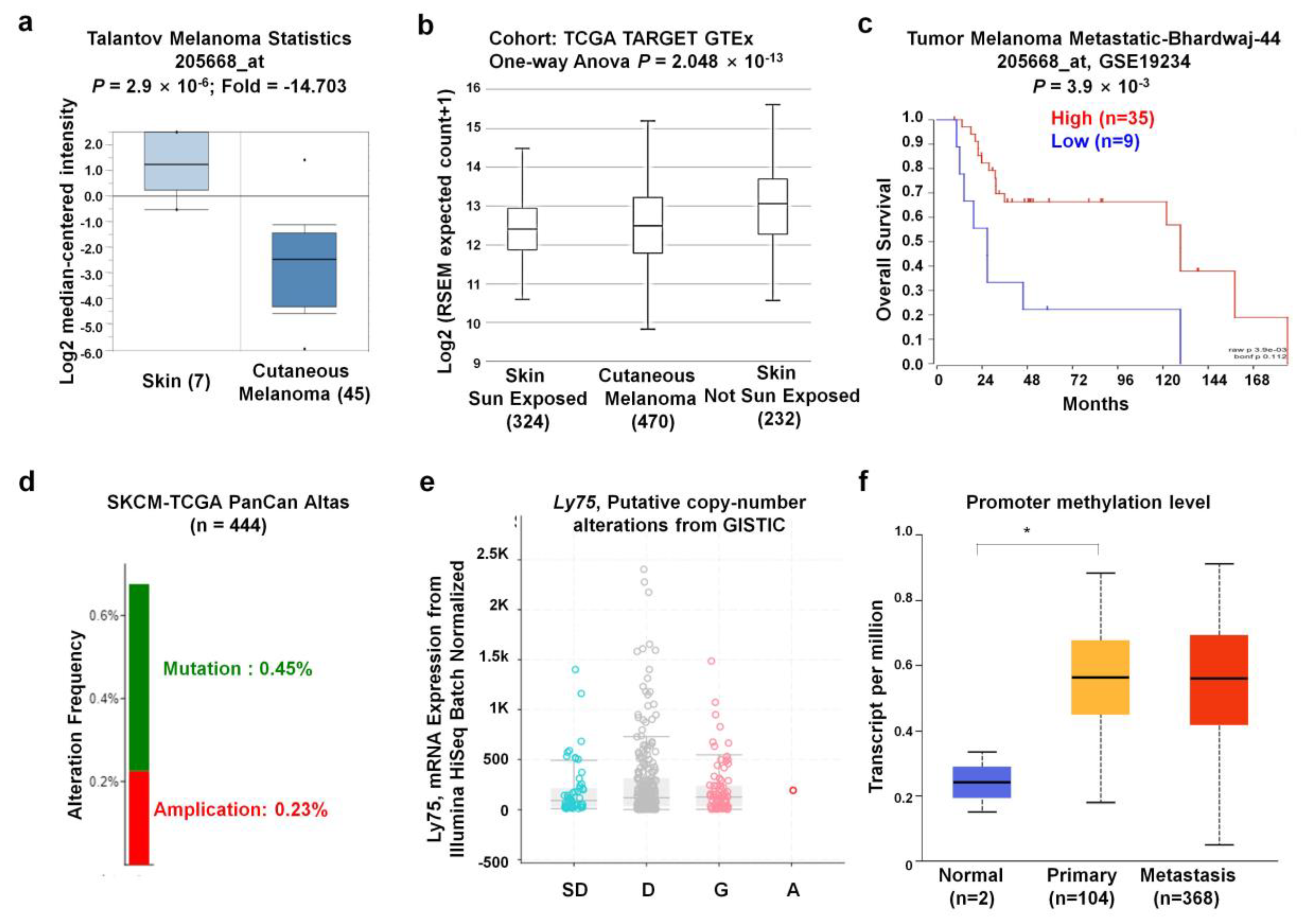
3.1. Ly75 mRNA Expression in Various Types of Cancers
3.2. Prognostic Value of Ly75 mRNA Expression in SKCM
3.3. Ly75 Gene Expression and Genome Alterations in SKCM
3.4. Immune Cell Infiltration by Ly75 mRNA Expression
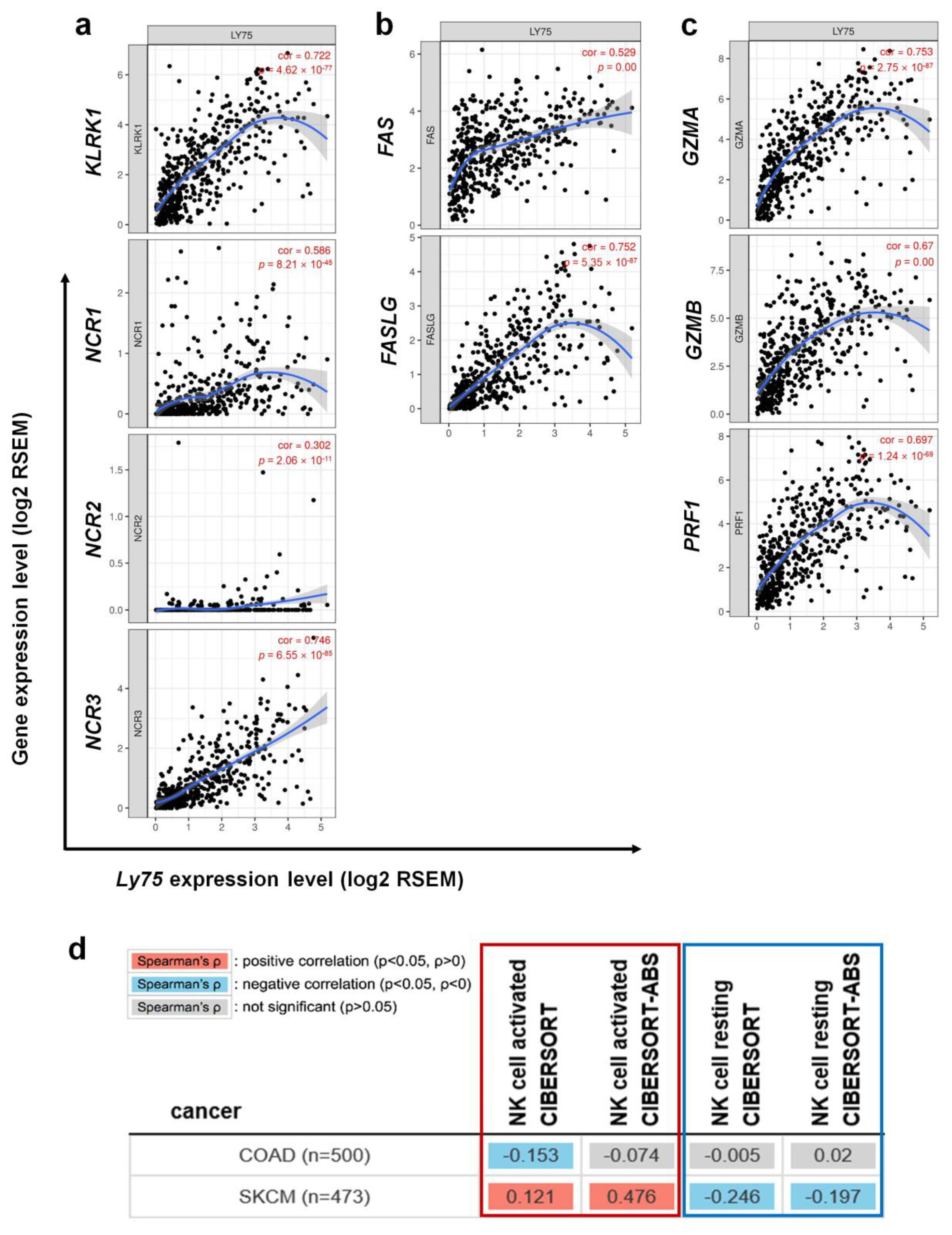
3.5. Association between Ly75 Expression and NK Cell Activation in SKCM
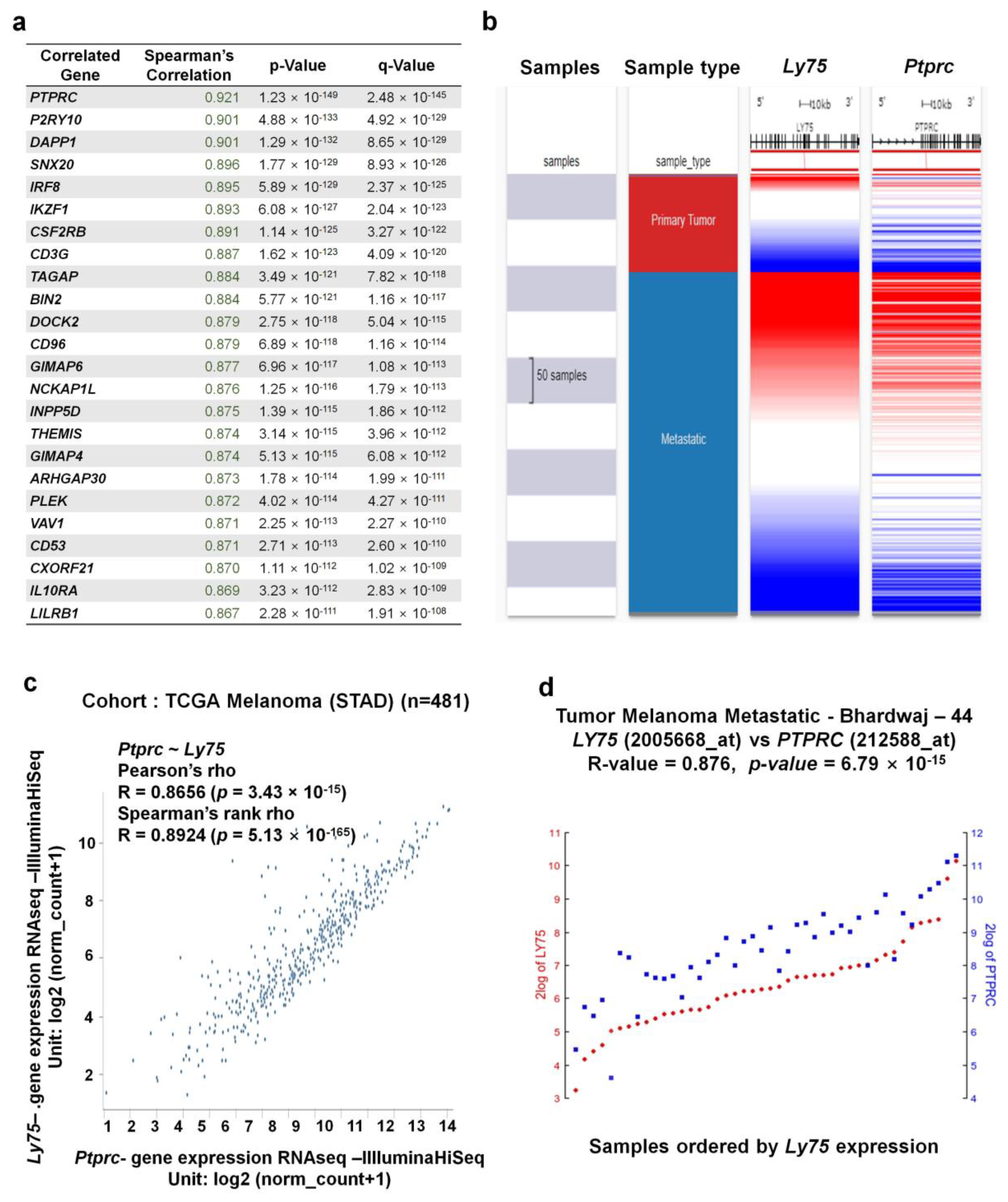
3.6. Co-Expressed Genes with Ly75 in SKCM
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- 2019 Surveillance of Melanoma: Assessment and Management (NICE Guideline NG14) and Improving Outcomes for People with Skin Tumours Including Melanoma (NICE Guideline CSG8); NICE: London, UK, 2019.
- Whiteman, D.C.; Green, A.C.; Olsen, C.M. The Growing Burden of Invasive Melanoma: Projections of Incidence Rates and Numbers of New Cases in Six Susceptible Populations through 2031. J. Investig. Dermatol. 2016, 136, 1161–1171. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- Soengas, M.S.; Lowe, S.W. Apoptosis and melanoma chemoresistance. Oncogene 2003, 22, 3138–3151. [Google Scholar] [CrossRef] [PubMed]
- Kluger, H.M.; Zito, C.R.; Barr, M.L.; Baine, M.K.; Chiang, V.L.; Sznol, M.; Rimm, D.L.; Chen, L.; Jilaveanu, L.B. Characterization of PD-L1 Expression and Associated T-cell Infiltrates in Metastatic Melanoma Samples from Variable Anatomic Sites. Clin. Cancer Res. 2015, 21, 3052–3060. [Google Scholar] [CrossRef] [PubMed]
- Goff, S.L.; Dudley, M.E.; Citrin, D.E.; Somerville, R.P.; Wunderlich, J.R.; Danforth, D.N.; Zlott, D.A.; Yang, J.C.; Sherry, R.M.; Kammula, U.S.; et al. Randomized, Prospective Evaluation Comparing Intensity of Lymphodepletion Before Adoptive Transfer of Tumor-Infiltrating Lymphocytes for Patients with Metastatic Melanoma. J. Clin. Oncol. 2016, 34, 2389–2397. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.; Zakka, L.R.; Mihm, M.C., Jr.; Schatton, T. Tumour-infiltrating lymphocytes in melanoma prognosis and cancer immunotherapy. Pathology 2016, 48, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Fu, Q.; Chen, N.; Ge, C.; Li, R.; Li, Z.; Zeng, B.; Li, C.; Wang, Y.; Xue, Y.; Song, X.; et al. Prognostic value of tumor-infiltrating lymphocytes in melanoma: A systematic review and meta-analysis. Oncoimmunology 2019, 8, 1593806. [Google Scholar] [CrossRef]
- Gata, V.A.; Lisencu, C.I.; Vlad, C.I.; Piciu, D.; Irimie, A.; Achimas-Cadariu, P. Tumor infiltrating lymphocytes as a prognostic factor in malignant melanoma. Review of the literature. J. BUON 2017, 22, 592–598. [Google Scholar]
- Shang, B.; Liu, Y.; Jiang, S.J.; Liu, Y. Prognostic value of tumor-infiltrating FoxP3+ regulatory T cells in cancers: A systematic review and meta-analysis. Sci. Rep. 2015, 5, 15179. [Google Scholar] [CrossRef]
- Quan, H.; Shan, Z.; Liu, Z.; Liu, S.; Yang, L.; Fang, X.; Li, K.; Wang, B.; Deng, Z.; Hu, Y.; et al. The repertoire of tumor-infiltrating lymphocytes within the microenvironment of oral squamous cell carcinoma reveals immune dysfunction. Cancer Immunol. Immunother. 2020, 69, 465–476. [Google Scholar] [CrossRef]
- Wang, J.; Matosevic, S. NT5E/CD73 as Correlative Factor of Patient Survival and Natural Killer Cell Infiltration in Glioblastoma. J. Clin. Med. 2019, 8, 1526. [Google Scholar] [CrossRef] [PubMed]
- Gil, M.; Kim, K.E. Interleukin-18 Is a Prognostic Biomarker Correlated with CD8(+) T Cell and Natural Killer Cell Infiltration in Skin Cutaneous Melanoma. J. Clin. Med. 2019, 8, 1993. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.H.; Zhou, H.; Cooper, L.; Huang, J.L.; Zhu, S.B.; Zhao, X.X.; Ding, H.; Pan, Y.L.; Rong, L. LAYN Is a Prognostic Biomarker and Correlated with Immune Infiltrates in Gastric and Colon Cancers. Front. Immunol. 2019, 10, 6. [Google Scholar] [CrossRef] [PubMed]
- Gentles, A.J.; Newman, A.M.; Liu, C.L.; Bratman, S.V.; Feng, W.; Kim, D.; Nair, V.S.; Xu, Y.; Khuong, A.; Hoang, C.D.; et al. The prognostic landscape of genes and infiltrating immune cells across human cancers. Nat. Med. 2015, 21, 938–945. [Google Scholar] [CrossRef] [PubMed]
- Bjorkstrom, N.K.; Ljunggren, H.G.; Michaelsson, J. Emerging insights into natural killer cells in human peripheral tissues. Nat. Rev. Immunol. 2016, 16, 310–320. [Google Scholar] [CrossRef] [PubMed]
- Waldhauer, I.; Steinle, A. NK cells and cancer immunosurveillance. Oncogene 2008, 27, 5932–5943. [Google Scholar] [CrossRef]
- Gras Navarro, A.; Bjorklund, A.T.; Chekenya, M. Therapeutic potential and challenges of natural killer cells in treatment of solid tumors. Front. Immunol. 2015, 6, 202. [Google Scholar] [CrossRef]
- Chavez-Galan, L.; Arenas-Del Angel, M.C.; Zenteno, E.; Chavez, R.; Lascurain, R. Cell death mechanisms induced by cytotoxic lymphocytes. Cell Mol. Immunol. 2009, 6, 15–25. [Google Scholar] [CrossRef]
- Smyth, M.J.; Hayakawa, Y.; Takeda, K.; Yagita, H. New aspects of natural-killer-cell surveillance and therapy of cancer. Nat. Rev. Cancer 2002, 2, 850–861. [Google Scholar] [CrossRef]
- Konjevic, G.; Vuletic, A.; Mirjacic Martinovic, K.; Krivokuca, A.; Jankovic, R.; Babovic, N. Evaluation of the Functional Capacity of NK Cells of Melanoma Patients in an In Vitro Model of NK Cell Contact with K562 and FemX Tumor Cell Lines. J. Membr. Biol. 2017, 250, 507–516. [Google Scholar] [CrossRef]
- Ali, T.H.; Pisanti, S.; Ciaglia, E.; Mortarini, R.; Anichini, A.; Garofalo, C.; Tallerico, R.; Santinami, M.; Gulletta, E.; Ietto, C.; et al. Enrichment of CD56(dim)KIR + CD57 + highly cytotoxic NK cells in tumour-infiltrated lymph nodes of melanoma patients. Nat. Commun. 2014, 5, 5639. [Google Scholar] [CrossRef] [PubMed]
- East, L.; Isacke, C.M. The mannose receptor family. Biochim. Biophys. Acta 2002, 1572, 364–386. [Google Scholar] [CrossRef]
- Jiang, W.; Swiggard, W.J.; Heufler, C.; Peng, M.; Mirza, A.; Steinman, R.M.; Nussenzweig, M.C. The receptor DEC-205 expressed by dendritic cells and thymic epithelial cells is involved in antigen processing. Nature 1995, 375, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Kato, M.; McDonald, K.J.; Khan, S.; Ross, I.L.; Vuckovic, S.; Chen, K.; Munster, D.; MacDonald, K.P.; Hart, D.N. Expression of human DEC-205 (CD205) multilectin receptor on leukocytes. Int. Immunol. 2006, 18, 857–869. [Google Scholar] [CrossRef] [PubMed]
- Tel, J.; Benitez-Ribas, D.; Hoosemans, S.; Cambi, A.; Adema, G.J.; Figdor, C.G.; Tacken, P.J.; de Vries, I.J. DEC-205 mediates antigen uptake and presentation by both resting and activated human plasmacytoid dendritic cells. Eur. J. Immunol. 2011, 41, 1014–1023. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.K.; Wall, K.A. Use of Dendritic Cell Receptors as Targets for Enhancing Anti-Cancer Immune Responses. Cancers 2019, 11, 418. [Google Scholar] [CrossRef]
- Fukaya, T.; Takagi, H.; Uto, T.; Arimura, K.; Sato, K. Analysis of DC Functions Using CD205-DTR Knock-In Mice. Methods Mol. Biol. 2016, 1423, 291–308. [Google Scholar] [CrossRef]
- Huarte, E.; Cubillos-Ruiz, J.R.; Nesbeth, Y.C.; Scarlett, U.K.; Martinez, D.G.; Buckanovich, R.J.; Benencia, F.; Stan, R.V.; Keler, T.; Sarobe, P.; et al. Depletion of dendritic cells delays ovarian cancer progression by boosting antitumor immunity. Cancer Res. 2008, 68, 7684–7691. [Google Scholar] [CrossRef]
- Tang, Z.; Kang, B.; Li, C.; Chen, T.; Zhang, Z. GEPIA2: An enhanced web server for large-scale expression profiling and interactive analysis. Nucleic Acids Res. 2019, 47, W556–W560. [Google Scholar] [CrossRef]
- Rhodes, D.R.; Kalyana-Sundaram, S.; Mahavisno, V.; Varambally, R.; Yu, J.; Briggs, B.B.; Barrette, T.R.; Anstet, M.J.; Kincead-Beal, C.; Kulkarni, P.; et al. Oncomine 3.0: Genes, pathways, and networks in a collection of 18,000 cancer gene expression profiles. Neoplasia 2007, 9, 166–180. [Google Scholar] [CrossRef]
- Shin, G.; Kang, T.W.; Yang, S.; Baek, S.J.; Jeong, Y.S.; Kim, S.Y. GENT: Gene expression database of normal and tumor tissues. Cancer Inform. 2011, 10, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Park, S.J.; Yoon, B.H.; Kim, S.K.; Kim, S.Y. GENT2: An updated gene expression database for normal and tumor tissues. BMC Med. Genom. 2019, 12, 101. [Google Scholar] [CrossRef] [PubMed]
- Vivian, J.; Rao, A.A.; Nothaft, F.A.; Ketchum, C.; Armstrong, J.; Novak, A.; Pfeil, J.; Narkizian, J.; Deran, A.D.; Musselman-Brown, A.; et al. Toil enables reproducible, open source, big biomedical data analyses. Nat. Biotechnol. 2017, 35, 314–316. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, D.R.; Yu, J.; Shanker, K.; Deshpande, N.; Varambally, R.; Ghosh, D.; Barrette, T.; Pandey, A.; Chinnaiyan, A.M. ONCOMINE: A cancer microarray database and integrated data-mining platform. Neoplasia 2004, 6, 1–6. [Google Scholar] [CrossRef]
- Chandrashekar, D.S.; Bashel, B.; Balasubramanya, S.A.H.; Creighton, C.J.; Ponce-Rodriguez, I.; Chakravarthi, B.; Varambally, S. UALCAN: A Portal for Facilitating Tumor Subgroup Gene Expression and Survival Analyses. Neoplasia 2017, 19, 649–658. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci. Signal 2013, 6, pl1. [Google Scholar] [CrossRef] [PubMed]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio cancer genomics portal: An open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef]
- Anaya, J. OncoLnc: Linking TCGA survival data to mRNAs, miRNAs, and lncRNAs. Peerj Comput. Sci. 2016, 2, e67. [Google Scholar] [CrossRef]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef]
- Faddaoui, A.; Bachvarova, M.; Plante, M.; Gregoire, J.; Renaud, M.C.; Sebastianelli, A.; Gobeil, S.; Morin, C.; Macdonald, E.; Vanderhyden, B.; et al. The mannose receptor LY75 (DEC205/CD205) modulates cellular phenotype and metastatic potential of ovarian cancer cells. Oncotarget 2016, 7, 14125–14142. [Google Scholar] [CrossRef]
- Holcakova, J. Effect of DNA Methylation on the Development of Cancer. Klin. Onkol. 2018, 31, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Wang, J.; Deng, X.; Xiong, F.; Ge, J.; Xiang, B.; Wu, X.; Ma, J.; Zhou, M.; Li, X.; et al. Role of the tumor microenvironment in PD-L1/PD-1-mediated tumor immune escape. Mol. Cancer 2019, 18, 10. [Google Scholar] [CrossRef] [PubMed]
- Cooper, L.A.; Gutman, D.A.; Chisolm, C.; Appin, C.; Kong, J.; Rong, Y.; Kurc, T.; Van Meir, E.G.; Saltz, J.H.; Moreno, C.S.; et al. The tumor microenvironment strongly impacts master transcriptional regulators and gene expression class of glioblastoma. Am. J. Pathol. 2012, 180, 2108–2119. [Google Scholar] [CrossRef] [PubMed]
- Galon, J.; Pages, F.; Marincola, F.M.; Thurin, M.; Trinchieri, G.; Fox, B.A.; Gajewski, T.F.; Ascierto, P.A. The immune score as a new possible approach for the classification of cancer. J. Transl. Med. 2012, 10, 1. [Google Scholar] [CrossRef] [PubMed]
- Senbabaoglu, Y.; Gejman, R.S.; Winer, A.G.; Liu, M.; Van Allen, E.M.; de Velasco, G.; Miao, D.; Ostrovnaya, I.; Drill, E.; Luna, A.; et al. Tumor immune microenvironment characterization in clear cell renal cell carcinoma identifies prognostic and immunotherapeutically relevant messenger RNA signatures. Genome Biol. 2016, 17, 231. [Google Scholar] [CrossRef] [PubMed]
- Schatton, T.; Scolyer, R.A.; Thompson, J.F.; Mihm, M.C., Jr. Tumor-infiltrating lymphocytes and their significance in melanoma prognosis. Methods Mol. Biol. 2014, 1102, 287–324. [Google Scholar] [CrossRef]
- Li, T.; Fan, J.; Wang, B.; Traugh, N.; Chen, Q.; Liu, J.S.; Li, B.; Liu, X.S. TIMER: A Web Server for Comprehensive Analysis of Tumor-Infiltrating Immune Cells. Cancer Res. 2017, 77, e108–e110. [Google Scholar] [CrossRef]
- Dhar, P.; Wu, J.D. NKG2D and its ligands in cancer. Curr. Opin. Immunol. 2018, 51, 55–61. [Google Scholar] [CrossRef]
- Chester, C.; Fritsch, K.; Kohrt, H.E. Natural Killer Cell Immunomodulation: Targeting Activating, Inhibitory, and Co-stimulatory Receptor Signaling for Cancer Immunotherapy. Front. Immunol. 2015, 6, 601. [Google Scholar] [CrossRef]
- Prager, I.; Watzl, C. Mechanisms of natural killer cell-mediated cellular cytotoxicity. J. Leukoc. Biol. 2019, 105, 1319–1329. [Google Scholar] [CrossRef]
- Bonifaz, L.; Bonnyay, D.; Mahnke, K.; Rivera, M.; Nussenzweig, M.C.; Steinman, R.M. Efficient targeting of protein antigen to the dendritic cell receptor DEC-205 in the steady state leads to antigen presentation on major histocompatibility complex class I products and peripheral CD8+ T cell tolerance. J. Exp. Med. 2002, 196, 1627–1638. [Google Scholar] [CrossRef] [PubMed]
- Birkholz, K.; Schwenkert, M.; Kellner, C.; Gross, S.; Fey, G.; Schuler-Thurner, B.; Schuler, G.; Schaft, N.; Dorrie, J. Targeting of DEC-205 on human dendritic cells results in efficient MHC class II-restricted antigen presentation. Blood 2010, 116, 2277–2285. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Jia, R.; Ge, S. Role of Epigenetics in Uveal Melanoma. Int. J. Biol. 2017, 13, 426–433. [Google Scholar] [CrossRef] [PubMed]
- Weisenberger, D.J.; Siegmund, K.D.; Campan, M.; Young, J.; Long, T.I.; Faasse, M.A.; Kang, G.H.; Widschwendter, M.; Weener, D.; Buchanan, D.; et al. CpG island methylator phenotype underlies sporadic microsatellite instability and is tightly associated with BRAF mutation in colorectal cancer. Nat. Genet. 2006, 38, 787–793. [Google Scholar] [CrossRef]
- Jung, H.; Kim, H.S.; Kim, J.Y.; Sun, J.M.; Ahn, J.S.; Ahn, M.J.; Park, K.; Esteller, M.; Lee, S.H.; Choi, J.K. DNA methylation loss promotes immune evasion of tumours with high mutation and copy number load. Nat. Commun. 2019, 10, 4278. [Google Scholar] [CrossRef]
- Mitra, S.; Lauss, M.; Cabrita, R.; Choi, J.; Zhang, T.; Isaksson, K.; Olsson, H.; Ingvar, C.; Carneiro, A.; Staaf, J.; et al. Analysis of DNA methylation patterns in the tumor immune microenvironment of metastatic melanoma. Mol. Oncol. 2020. [Google Scholar] [CrossRef]
- Ohtani, H. Focus on TILs: Prognostic significance of tumor infiltrating lymphocytes in human colorectal cancer. Cancer Immun. 2007, 7, 4. [Google Scholar]
- Jin, S.; Deng, Y.; Hao, J.W.; Li, Y.; Liu, B.; Yu, Y.; Shi, F.D.; Zhou, Q.H. NK cell phenotypic modulation in lung cancer environment. PLoS ONE 2014, 9, e109976. [Google Scholar] [CrossRef]
- Cursons, J.; Souza-Fonseca-Guimaraes, F.; Foroutan, M.; Anderson, A.; Hollande, F.; Hediyeh-Zadeh, S.; Behren, A.; Huntington, N.D.; Davis, M.J. A Gene Signature Predicting Natural Killer Cell Infiltration and Improved Survival in Melanoma Patients. Cancer Immunol. Res. 2019, 7, 1162–1174. [Google Scholar] [CrossRef]
- Vivier, E.; Raulet, D.H.; Moretta, A.; Caligiuri, M.A.; Zitvogel, L.; Lanier, L.L.; Yokoyama, W.M.; Ugolini, S. Innate or adaptive immunity? The example of natural killer cells. Science 2011, 331, 44–49. [Google Scholar] [CrossRef]
- Rossi, G.R.; Trindade, E.S.; Souza-Fonseca-Guimaraes, F. Tumor Microenvironment-Associated Extracellular Matrix Components Regulate NK Cell Function. Front. Immunol. 2020, 11, 73. [Google Scholar] [CrossRef] [PubMed]
- Vitale, M.; Cantoni, C.; Pietra, G.; Mingari, M.C.; Moretta, L. Effect of tumor cells and tumor microenvironment on NK-cell function. Eur. J. Immunol. 2014, 44, 1582–1592. [Google Scholar] [CrossRef] [PubMed]
- Gajewski, T.F.; Schreiber, H.; Fu, Y.X. Innate and adaptive immune cells in the tumor microenvironment. Nat. Immunol. 2013, 14, 1014–1022. [Google Scholar] [CrossRef] [PubMed]
- Zou, W. Regulatory T cells, tumour immunity and immunotherapy. Nat. Rev. Immunol. 2006, 6, 295–307. [Google Scholar] [CrossRef]
- Betts, G.; Jones, E.; Junaid, S.; El-Shanawany, T.; Scurr, M.; Mizen, P.; Kumar, M.; Jones, S.; Rees, B.; Williams, G.; et al. Suppression of tumour-specific CD4(+) T cells by regulatory T cells is associated with progression of human colorectal cancer. Gut 2012, 61, 1163–1171. [Google Scholar] [CrossRef]
- Colegio, O.R.; Chu, N.Q.; Szabo, A.L.; Chu, T.; Rhebergen, A.M.; Jairam, V.; Cyrus, N.; Brokowski, C.E.; Eisenbarth, S.C.; Phillips, G.M.; et al. Functional polarization of tumour-associated macrophages by tumour-derived lactic acid. Nature 2014, 513, 559–563. [Google Scholar] [CrossRef]
- Holmes, N. CD45: All is not yet crystal clear. Immunology 2006, 117, 145–155. [Google Scholar] [CrossRef]
- Krzywinska, E.; Cornillon, A.; Allende-Vega, N.; Vo, D.N.; Rene, C.; Lu, Z.Y.; Pasero, C.; Olive, D.; Fegueux, N.; Ceballos, P.; et al. CD45 Isoform Profile Identifies Natural Killer (NK) Subsets with Differential Activity. PLoS ONE 2016, 11, e0150434. [Google Scholar] [CrossRef]
- Freud, A.G.; Yu, J.; Caligiuri, M.A. Human natural killer cell development in secondary lymphoid tissues. Semin. Immunol. 2014, 26, 132–137. [Google Scholar] [CrossRef]
- Beziat, V.; Descours, B.; Parizot, C.; Debre, P.; Vieillard, V. NK cell terminal differentiation: Correlated stepwise decrease of NKG2A and acquisition of KIRs. PLoS ONE 2010, 5, e11966. [Google Scholar] [CrossRef]
- Clausen, J.; Vergeiner, B.; Enk, M.; Petzer, A.L.; Gastl, G.; Gunsilius, E. Functional significance of the activation-associated receptors CD25 and CD69 on human NK-cells and NK-like T-cells. Immunobiology 2003, 207, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Berridge, M.J.; Irvine, R.F. Inositol phosphates and cell signalling. Nature 1989, 341, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Karathanassis, D.; Stahelin, R.V.; Bravo, J.; Perisic, O.; Pacold, C.M.; Cho, W.; Williams, R.L. Binding of the PX domain of p47(phox) to phosphatidylinositol 3,4-bisphosphate and phosphatidic acid is masked by an intramolecular interaction. EMBO J. 2002, 21, 5057–5068. [Google Scholar] [CrossRef] [PubMed]
- Tuosto, L.; Capuano, C.; Muscolini, M.; Santoni, A.; Galandrini, R. The multifaceted role of PIP2 in leukocyte biology. Cell. Mol. Life Sci. 2015, 72, 4461–4474. [Google Scholar] [CrossRef] [PubMed]
- Porciello, N.; Kunkl, M.; Viola, A.; Tuosto, L. Phosphatidylinositol 4-Phosphate 5-Kinases in the Regulation of T Cell Activation. Front. Immunol. 2016, 7, 186. [Google Scholar] [CrossRef]
- Colonna, M. Below the NK-cell surface: PIP2. Blood 2008, 111, 3916. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gumbleton, M.; Kerr, W.G. Role of inositol phospholipid signaling in natural killer cell biology. Front. Immunol. 2013, 4, 47. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Description | Gene Markers | SKCM | COAD | ||||||
|---|---|---|---|---|---|---|---|---|---|
| None | Purity | None | Purity | ||||||
| Cor | p | Cor | p | Cor | p | Cor | p | ||
| T cell (general) | CD3D | 0.776 | *** | 0.642 | *** | 0.000 | 0.994 | 0.019 | 0.704 |
| CD3E | 0.769 | *** | 0.634 | *** | 0.034 | 0.467 | 0.072 | 0.150 | |
| CD2 | 0.807 | *** | 0.695 | *** | 0.097 | 0.037 | 0.147 | * | |
| CD8+ T cell | CD8A | 0.748 | *** | 0.618 | *** | 0.030 | 0.520 | 0.047 | 0.347 |
| CD8B | 0.746 | *** | 0.609 | *** | 0.067 | 0.152 | 0.086 | 0.082 | |
| Th1 | T-bet (TBX21) | 0.765 | *** | 0.635 | *** | 0.047 | 0.317 | 0.099 | 0.047 |
| STAT4 | 0.790 | *** | 0.694 | *** | 0.124 | * | 0.143 | 0.004 | |
| STAT1 | 0.666 | *** | 0.588 | *** | 0.244 | *** | 0.268 | *** | |
| IFNγ (IFNG) | 0.707 | *** | 0.570 | *** | 0.081 | 0.082 | 0.087 | 0.080 | |
| TNFα (TNF) | 0.687 | *** | 0.540 | *** | 0.073 | 0.121 | 0.102 | 0.039 | |
| Th2 | GATA3 | 0.748 | *** | 0.600 | *** | 0.082 | 0.080 | 0.118 | 0.018 |
| STAT6 | 0.087 | 0.059 | 0.136 | * | 0.270 | *** | 0.262 | *** | |
| STAT5A | 0.272 | *** | 0.327 | *** | 0.216 | *** | 0.227 | *** | |
| IL13 | 0.208 | *** | 0.130 | * | 0.017 | 0.719 | 0.022 | 0.657 | |
| Tfh | BCL6 | 0.435 | *** | 0.400 | *** | 0.113 | 0.016 | 0.153 | * |
| IL21 | 0.583 | *** | 0.482 | *** | 0.104 | 0.026 | 0.102 | 0.040 | |
| Th17 | STAT3 | 0.419 | *** | 0.444 | *** | 0.379 | *** | 0.414 | *** |
| IL17A | −0.048 | 0.294 | −0.119 | 0.011 | 0.160 | * | 0.169 | * | |
| γδ T cell | RORγt (RORC) | 0.400 | *** | 0.280 | *** | 0.158 | * | 0.145 | * |
| CCR5 | 0.816 | *** | 0.722 | *** | 0.148 | * | 0.188 | ** | |
| CXCR6 | 0.768 | *** | 0.644 | *** | 0.072 | 0.125 | 0.111 | 0.026 | |
| VNN2 | 0.784 | *** | 0.689 | *** | 0.010 | 0.826 | 0.033 | 0.507 | |
| CD300A | 0.710 | *** | 0.575 | *** | −0.048 | 0.304 | −0.028 | 0.578 | |
| Treg | FOXP3 | 0.728 | *** | 0.587 | *** | 0.082 | 0.081 | 0.112 | 0.024 |
| CCR8 | 0.815 | *** | 0.737 | *** | 0.183 | *** | 0.209 | *** | |
| STAT5B | 0.403 | *** | 0.509 | *** | 0.455 | *** | 0.467 | *** | |
| TGFβ (TGFB1) | 0.432 | *** | 0.287 | *** | −0.037 | 0.425 | −0.009 | 0.849 | |
| B cell | CD19 | 0.654 | *** | 0.528 | *** | 0.091 | 0.053 | 0.158 | * |
| CD79A | 0.647 | *** | 0.495 | *** | 0.110 | 0.019 | 0.163 | * | |
| Monocyte | CD86 | 0.822 | *** | 0.732 | *** | 0.081 | 0.084 | 0.121 | 0.015 |
| CD115 (CSF1R) | 0.753 | *** | 0.648 | *** | 0.059 | 0.210 | 0.085 | 0.088 | |
| TAM | CCL2 | 0.614 | *** | 0.464 | *** | 0.123 | * | 0.141 | * |
| CD68 | 0.422 | *** | 0.231 | *** | 0.078 | 0.094 | 0.113 | 0.023 | |
| IL10 | 0.627 | *** | 0.489 | *** | 0.042 | 0.367 | 0.073 | 0.140 | |
| M1 Macrophage | INOS (NOS2) | 0.100 | 0.029 | 0.118 | 0.012 | −0.037 | 0.432 | −0.033 | 0.511 |
| IRF5 | 0.641 | *** | 0.478 | *** | 0.218 | *** | 0.227 | 0.000 | |
| COX2 (PTGS2) | 0.164 | ** | 0.106 | 0.023 | 0.049 | 0.300 | 0.057 | 0.248 | |
| M2 Macrophage | CD163 | 0.663 | *** | 0.550 | *** | 0.086 | 0.066 | 0.110 | 0.026 |
| VSIG4 | 0.595 | *** | 0.473 | *** | −0.019 | 0.683 | −0.004 | 0.930 | |
| MS4A4A | 0.698 | *** | 0.580 | *** | 0.023 | 0.625 | 0.049 | 0.326 | |
| Neutrophils | CD66b (CEACAM8) | 0.027 | 0.561 | 0.067 | 0.153 | 0.033 | 0.479 | 0.012 | 0.812 |
| CD11b (ITGAM) | 0.671 | *** | 0.570 | *** | −0.013 | 0.779 | 0.011 | 0.829 | |
| CCR7 | 0.747 | *** | 0.604 | *** | 0.114 | 0.015 | 0.158 | * | |
| Natural killer cell | KIR2DL1 | 0.360 | *** | 0.203 | *** | −0.161 | * | −0.198 | *** |
| KIR2DL3 | 0.490 | *** | 0.302 | *** | −0.122 | 0.009 | −0.129 | * | |
| KIR2DL4 | 0.571 | *** | 0.390 | *** | −0.100 | 0.032 | −0.093 | 0.060 | |
| KIR3DL1 | 0.461 | *** | 0.283 | *** | −0.044 | 0.352 | −0.053 | 0.290 | |
| KIR3DL2 | 0.576 | *** | 0.405 | *** | −0.048 | 0.301 | −0.049 | 0.323 | |
| KIR3DL3 | 0.138 | * | 0.067 | 0.155 | −0.066 | 0.160 | −0.058 | 0.241 | |
| KIR2DS4 | 0.401 | *** | 0.264 | *** | −0.085 | 0.068 | −0.100 | 0.044 | |
| KLRK1 (NKG2D) | 0.722 | *** | 0.597 | *** | 0.026 | 0.575 | 0.047 | 0.344 | |
| NCR1 (NKp46) | 0.586 | *** | 0.475 | *** | −0.046 | 0.330 | −0.029 | 0.557 | |
| NCR2 (NKp44) | 0.302 | *** | 0.247 | *** | −0.053 | 0.254 | −0.049 | 0.327 | |
| NCR3 (NKp30) | 0.746 | *** | 0.612 | *** | −0.030 | 0.523 | 0.017 | 0.733 | |
| Dendritic cell | HLA-DPB1 | 0.748 | *** | 0.611 | *** | −0.055 | 0.240 | −0.026 | 0.604 |
| HLA-DQB1 | 0.690 | *** | 0.535 | *** | 0.019 | 0.690 | 0.047 | 0.349 | |
| HLA-DRA | 0.785 | *** | 0.668 | *** | 0.033 | 0.481 | 0.062 | 0.211 | |
| HLA-DPA1 | 0.734 | *** | 0.606 | *** | 0.073 | 0.118 | 0.114 | 0.022 | |
| BDCA1 (CD1C) | 0.671 | *** | 0.535 | *** | 0.150 | * | 0.186 | ** | |
| BDCA4 (NRP1) | 0.505 | *** | 0.456 | *** | 0.151 | * | 0.206 | *** | |
| CD11c (ITGAX) | 0.578 | *** | 0.398 | *** | 0.038 | 0.412 | 0.079 | 0.110 | |
| Cell Type | Gene Markers | SKCM | COAD | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Tumor | Normal | Tumor | Normal | ||||||
| R | p | R | p | R | p | R | p | ||
| CD8+ T cells | CD8A | 0.43 | *** | 0.099 | 0.019 | 0.0028 | 0.96 | 0.56 | *** |
| CD8B | 0.45 | *** | 0.14 | ** | 0.062 | 0.31 | 0.6 | *** | |
| NK cells | KIR2DL1 | 0.098 | * | 0.17 | 0.059 | 0.012 | 0.84 | 0.073 | 0.17 |
| KIR2DL3 | 0.17 | ** | 0.038 | 0.038 | 0.024 | 0.69 | 0.24 | *** | |
| KIR2DL4 | 0.25 | *** | 0.055 | 0.036 | 0.55 | 0.064 | 0.7 | *** | |
| KIR3DL1 | 0.31 | *** | 0.067 | 0.12 | 0.0053 | 0.93 | 0.19 | ** | |
| KIR3DL2 | 0.42 | *** | 0.043 | 0.42 | 0.014 | 0.81 | 0.5 | *** | |
| KIR3DL3 | 0.095 | * | 0.033 | 0.43 | −0.04 | 0.51 | 0.16 | * | |
| KIR2DS4 | 0.073 | 0.12 | 0.041 | 0.34 | 0.057 | 0.34 | 0.39 | *** | |
| KLRK1 (NKG2D) | 0.48 | *** | 0.18 | *** | 0.0091 | 0.88 | 0.39 | *** | |
| NCR1 (NKp46) | 0.21 | *** | 0.087 | 0.039 | 0.047 | 0.44 | 0.16 | * | |
| NCR2 (NKp44) | 0.25 | *** | 0.03 | 0.48 | 0.021 | 0.73 | 0.51 | *** | |
| NCR3 (NKp30) | 0.62 | *** | 0.2 | *** | −0.011 | 0.85 | 0.42 | *** | |
| γδ T cell | RORC (RORγt) | 0.05 | 0.28 | 0.016 | 0.7 | 0.06 | 0.32 | 0.81 | *** |
| CCR5 | 0.51 | *** | 0.14 | ** | 0.092 | 0.13 | −0.081 | 0.13 | |
| CXCR6 | 0.48 | *** | 0.17 | *** | 0.1 | 0.083 | 0.65 | *** | |
| VNN2 | 0.65 | *** | −0.09 | 0.24 | −0.12 | 0.046 | 0.029 | 0.59 | |
| CD300A | 0.51 | *** | 0.12 | * | −0.072 | 0.23 | 0.54 | *** | |
| Gene Markers | SKCM | COAD | |||
|---|---|---|---|---|---|
| R | p | R | p | ||
| Activation receptors | KLRK1 | 0.722 | *** | 0.026 | 0.575 |
| NCR1 | 0.586 | *** | −0.046 | 0.330 | |
| NCR2 | 0.302 | *** | −0.053 | 0.254 | |
| NCR3 | 0.746 | *** | −0.030 | 0.523 | |
| FAS & FASL | FAS | 0.529 | *** | 0.032 | 0.494 |
| FASLG | 0.752 | *** | 0.078 | 0.097 | |
| Cytolytic molecules | GZMA | 0.753 | *** | 0.006 | 0.891 |
| GZMB | 0.670 | *** | 0.096 | 0.040 | |
| PRF1 | 0.697 | *** | 0.004 | 0.928 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gil, M.; Kim, K.E. Systematic Multiomic Analysis of Ly75 Gene Expression and Its Prognostic Value through the Infiltration of Natural Killer (NK) Cells in Skin Cutaneous Melanoma. J. Clin. Med. 2020, 9, 1383. https://doi.org/10.3390/jcm9051383
Gil M, Kim KE. Systematic Multiomic Analysis of Ly75 Gene Expression and Its Prognostic Value through the Infiltration of Natural Killer (NK) Cells in Skin Cutaneous Melanoma. Journal of Clinical Medicine. 2020; 9(5):1383. https://doi.org/10.3390/jcm9051383
Chicago/Turabian StyleGil, Minchan, and Kyung Eun Kim. 2020. "Systematic Multiomic Analysis of Ly75 Gene Expression and Its Prognostic Value through the Infiltration of Natural Killer (NK) Cells in Skin Cutaneous Melanoma" Journal of Clinical Medicine 9, no. 5: 1383. https://doi.org/10.3390/jcm9051383
APA StyleGil, M., & Kim, K. E. (2020). Systematic Multiomic Analysis of Ly75 Gene Expression and Its Prognostic Value through the Infiltration of Natural Killer (NK) Cells in Skin Cutaneous Melanoma. Journal of Clinical Medicine, 9(5), 1383. https://doi.org/10.3390/jcm9051383