The Impact of the Circadian Genes CLOCK and ARNTL on Myocardial Infarction




Abstract
1. Introduction
2. Experimental Section
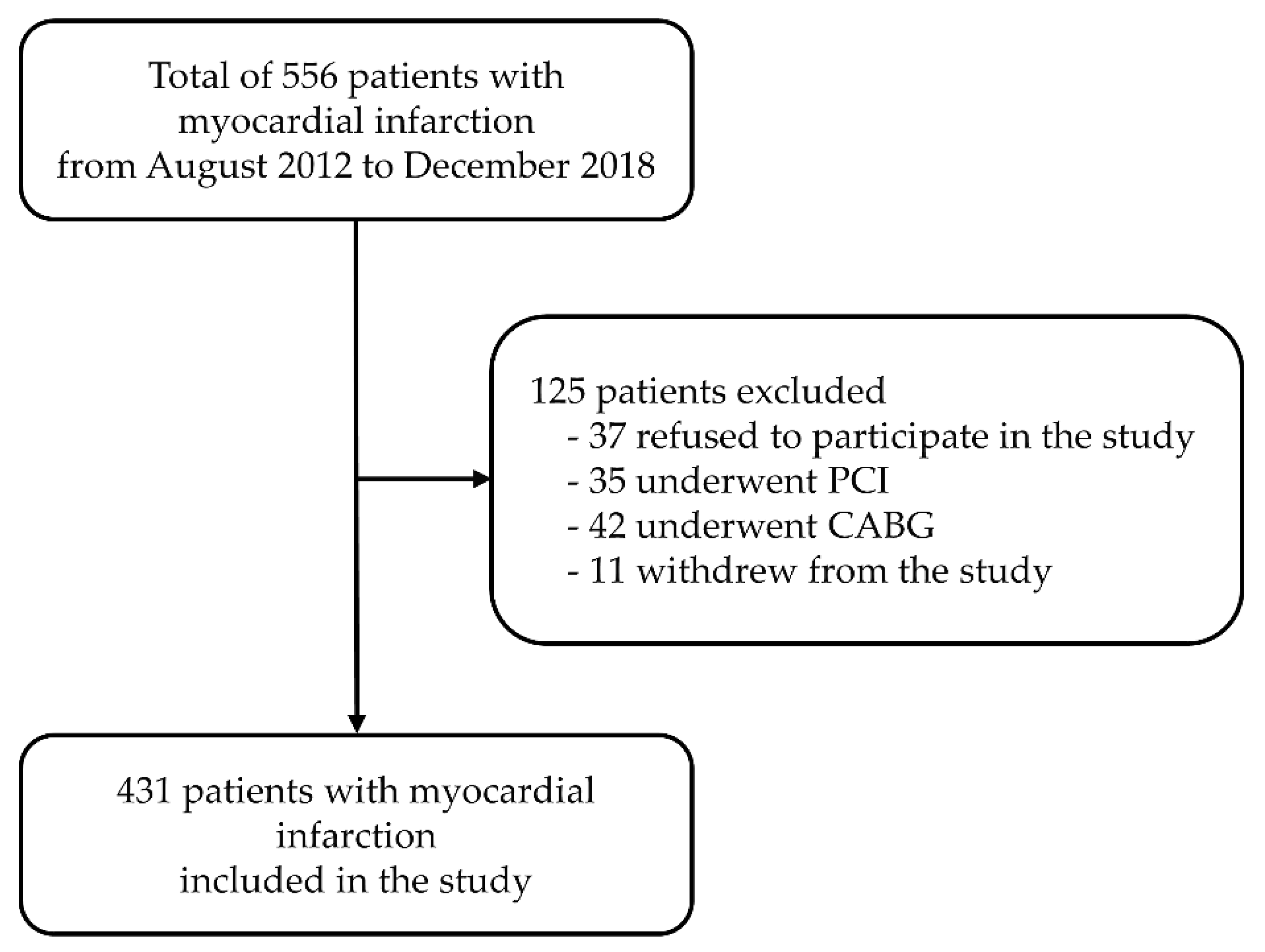
2.1. Participants
2.2. SNP Selection and Genotyping
2.3. Statistical Analysis
3. Results
4. Discussion
Limitations of the Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Noncommunicable Diseases Progress Monitor, 2017; WHO: Geneva, Switzerland, 2017. [Google Scholar]
- Kralj, V.; Brkić Biloš, I. Cardiovascular diseases – mortality and morbidity trends in Croatia, Europe and worldwide. Cardiol. Croat. 2016, 11, 504. [Google Scholar] [CrossRef]
- World Health Organization. Noncommunicable Diseases Country Profiles 2018; WHO: Geneva, Switzerland, 2018. [Google Scholar]
- Stevanovic, R.; Capak, K.; Brkic, K. Croatian Health Statistics Yearbook 2016; Croatian Institute of Public Health: Zagreb, Croatia, 2017. [Google Scholar]
- Thygesen, K.; Alpert, J.S.; White, H.D. Universal definition of myocardial infarction. Eur. Heart J. 2007, 28, 2525–2538. [Google Scholar] [CrossRef]
- Škrlec, I. Circadian rhythm and myocardial infarction. Med. Flum. 2019, 55, 32–42. [Google Scholar] [CrossRef][Green Version]
- Škrlec, I.; Marić, S.; Včev, A. Myocardial infarction and circadian rhythm. In Visions Cardiomyocyte-Fundamental Concepts of Heart and Life Disease; Tsipis, A., Ed.; IntechOpen: London, UK, 2019; pp. 60–75. [Google Scholar] [CrossRef]
- Partch, C.L.; Green, C.B.; Takahashi, J.S. Molecular architecture of the mammalian circadian clock. Trends Cell Biol. 2014, 24, 90–99. [Google Scholar] [CrossRef]
- Touitou, Y.; Bogdan, A. Circadian and seasonal variations of physiological and biochemical determinants of acute myocardial infarction. Biol. Rhythm Res. 2007, 38, 169–179. [Google Scholar] [CrossRef]
- Muller, J.E.; Stone, P.H.; Turi, Z.G.; Rutherford, J.D.; Czeisler, C.A.; Parker, C.; Poole, W.K.; Passamani, E.; Roberts, R.; Robertson, T.; et al. Circadian variation in the frequency of onset of acute myocardial infarction. N. Engl. J. Med. 1985, 313, 1315–1322. [Google Scholar] [CrossRef]
- Mehta, R.H.; Manfredini, R.; Hassan, F.; Sechtem, U.; Bossone, E.; Oh, J.K.; Cooper, J.V.; Smith, D.E.; Portaluppi, F.; Penn, M.; et al. Chronobiological patterns of acute aortic dissection. Circulation 2002, 106, 1110–1115. [Google Scholar] [CrossRef]
- Elliott, W.J. Circadian variation in the timing of stroke onset: A meta-analysis. Stroke 1998, 29, 992–996. [Google Scholar] [CrossRef]
- Manfredini, R.; Boari, B.; Smolensky, M.H.; Salmi, R.; Gallerani, M.; Guerzoni, F.; Guerra, V.; Malagoni, A.M.; Manfredini, F. Seasonal variation in onset of myocardial infarction—A 7-year single-center study in Italy. Chronobiol. Int. 2005, 22, 1121–1135. [Google Scholar] [CrossRef]
- Garaulet, M.; Madrid, J.A. Chronobiology, genetics and metabolic syndrome. Curr. Opin. Lipidol. 2009, 20, 127–134. [Google Scholar] [CrossRef]
- Virag, J.A.I.; Lust, R.M. Circadian influences on myocardial infarction. Front. Physiol. 2014, 5, 422. [Google Scholar] [CrossRef]
- Leu, H.B.; Chung, C.M.; Lin, S.J.; Chiang, K.M.; Yang, H.C.; Ho, H.Y.; Ting, C.T.; Lin, T.H.; Sheu, S.H.; Tsai, W.C.; et al. Association of circadian genes with diurnal blood pressure changes and non-dipper essential hypertension: A genetic association with young-onset hypertension. Hypertens. Res. 2015, 38, 155–162. [Google Scholar] [CrossRef]
- Kelly, M.A.; Rees, S.D.; Hydrie, M.Z.I.; Shera, A.S.; Bellary, S.; O’Hare, J.P.; Kumar, S.; Taheri, S.; Basit, A.; Barnett, A.H.; et al. Circadian gene variants and susceptibility to type 2 diabetes: A pilot study. PLoS ONE 2012, 7, e32670. [Google Scholar] [CrossRef]
- Englund, A.; Kovanen, L.; Saarikoski, S.T.; Haukka, J.; Reunanen, A.; Aromaa, A.; Lönnqvist, J.; Partonen, T. NPAS2 and PER2 are linked to risk factors of the metabolic syndrome. J. Circadian Rhythm. 2009, 7, 5. [Google Scholar] [CrossRef]
- Suzuki, S.; Ishii, H.; Ichimiya, S.; Kanashiro, M.; Watanabe, J.; Uchida, Y.; Yoshikawa, D.; Maeda, K.; Matsubara, T.; Murohara, T. Impact of the circadian rhythm on microvascular function in patients with ST-elevation myocardial infarction. Int. J. Cardiol. 2013, 168, 4948–4949. [Google Scholar] [CrossRef]
- Poggiogalle, E.; Jamshed, H.; Peterson, C.M. Circadian regulation of glucose, lipid, and energy metabolism in humans. Metabolism 2018, 84, 11–27. [Google Scholar] [CrossRef]
- Manfredini, R.; Fabbian, F.; Cappadona, R.; Modesti, P.A. Daylight saving time, circadian rhythms, and cardiovascular health. Intern. Emerg. Med. 2018, 13, 641–646. [Google Scholar] [CrossRef]
- Martino, T.A.; Young, M.E. Influence of the cardiomyocyte circadian clock on cardiac physiology and pathophysiology. J. Biol. Rhythm. 2015, 30, 183–205. [Google Scholar] [CrossRef]
- Durgan, D.J.; Young, M.E. The cardiomyocyte circadian clock: Emerging roles in health and disease. Circ. Res. 2010, 106, 647–658. [Google Scholar] [CrossRef]
- Gómez-Abellán, P.; Madrid, J.A.; Ordovás, J.M.; Garaulet, M. Chronobiological aspects of obesity and metabolic syndrome. Endocrinol. Nutr. 2012, 59, 50–61. [Google Scholar] [CrossRef]
- Škrlec, I.; Milić, J.; Heffer, M.; Steiner, R.; Peterlin, B.; Wagner, J. Association of Circadian Rhythm with Myocardial Infarction. Acta Clin. Croat. 2018, 57, 480–485. [Google Scholar] [CrossRef] [PubMed]
- Erdmann, J.; Linsel-Nitschke, P.; Schunkert, H. Genetic causes of myocardial infarction: New insights from genome-wide association studies. Dtsch. Arztebl. Int. 2010, 107, 694–699. [Google Scholar] [CrossRef] [PubMed]
- Takeda, N.; Maemura, K. Circadian clock and the onset of cardiovascular events. Hypertens. Res. 2016, 39, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Kelleher, F.C.; Rao, A.; Maguire, A. Circadian molecular clocks and cancer. Cancer Lett. 2014, 342, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Škrlec, I.; Milić, J.; Heffer, M.; Peterlin, B.; Wagner, J. Genetic variations in circadian rhythm genes and susceptibility for myocardial infarction. Genet. Mol. Biol. 2018, 41, 403–409. [Google Scholar] [CrossRef]
- Škrlec, I.; Milić, J.; Heffer, M.; Wagner, J.; Peterlin, B. Circadian clock genes and circadian phenotypes in patients with myocardial infarction. Adv. Med. Sci. 2019, 64, 224–229. [Google Scholar] [CrossRef]
- Thygesen, K.; Alpert, J.S.; Jaffe, A.S.; Simoons, M.L.; Chaitman, B.R.; White, H.D. Third universal definition of myocardial infarction. Nat. Rev. Cardiol. 2012, 9, 620–633. [Google Scholar] [CrossRef]
- Thygesen, K.; Alpert, J.S.; Jaffe, A.S.; Chaitman, B.R.; Bax, J.J.; Morrow, D.A.; White, H.D.; Mickley, H.; Crea, F.; Van de Werf, F.; et al. Fourth universal definition of myocardial infarction (2018). Eur. Heart J. 2019, 40, 237–269. [Google Scholar] [CrossRef]
- Elder, S.J.; Lichtenstein, A.H.; Pittas, A.G.; Roberts, S.B.; Fuss, P.J.; Greenberg, A.S.; McCrory, MA.; Bouchard, T.J., Jr.; Saltzman, E.; Neale, M.C. Genetic and environmental influences on factors associated with cardiovascular disease and the metabolic syndrome. J. Lipid Res. 2009, 50, 1917–1926. [Google Scholar] [CrossRef]
- de Bakker, P.I.W.; Yelensky, R.; Pe’er, I.; Gabriel, S.B.; Daly, M.J.; Altshuler, D. Efficiency and power in genetic association studies. Nat. Genet. 2005, 37, 1217–1223. [Google Scholar] [CrossRef]
- Solé, X.; Guinó, E.; Valls, J.; Iniesta, R.; Moreno, V. SNPStats: A web tool for the analysis of association studies. Bioinformatics 2006, 22, 1928–1929. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.Y.; He, L. SHEsis, a powerful software platform for analyses of linkage disequilibrium, haplotype construction, and genetic association at polymorphism loci. Cell Res. 2005, 15, 97–98. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, Z.; He, Z.; Tang, W.; Li, T.; Zeng, Z.; He, L.; Shi, Y.Y. A partition-ligation-combination-subdivision EM algorithm for haplotype inference with multiallelic markers: Update of the SHEsis (http://analysis.bio-x.cn). Cell Res. 2009, 19, 519–523. [Google Scholar] [CrossRef] [PubMed]
- Bray, M.S.; Shaw, C.A.; Moore, M.W.S.; Garcia, R.A.P.; Zanquetta, M.M.; Durgan, D.J.; Jeong, W.J.; Tsai, Ju.; Bugger, H.; Zhang, D.; et al. Disruption of the circadian clock within the cardiomyocyte influences myocardial contractile function, metabolism, and gene expression. Am. J. Physiol. Heart Circ. Physiol. 2008, 294, H1036–H1047. [Google Scholar] [CrossRef] [PubMed]
- Khaper, N.; Bailey, C.D.C.; Ghugre, N.R.; Reitz, C.; Awosanmi, Z.; Waines, R.; Martino, T.A. Implications of disturbances in circadian rhythms for cardiovascular health: A new frontier in free radical biology. Free Radic. Biol. Med. 2017, 119, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Rabinovich-Nikitin, I.; Lieberman, B.; Martino, T.A.; Kirshenbaum, L.A. Circadian-Regulated Cell Death in Cardiovascular Diseases. Circulation 2019, 139, 965–980. [Google Scholar] [CrossRef] [PubMed]
- Scott, E.M. Circadian clocks, obesity and cardiometabolic function. Diabetes Obes. Metab. 2015, 17, 84–89. [Google Scholar] [CrossRef]
- Huang, W.; Ramsey, K.M.; Marcheva, B.; Bass, J. Circadian rhythms, sleep, and metabolism. J. Clin. Investig. 2011, 121, 2133–2141. [Google Scholar] [CrossRef]
- Pedrazzoli, M.; Secolin, R.; Esteves, L.O.B.; Pereira, D.S.; Koike, B.D.V.; Louzada, F.M.; Lopes-Cendes, I.; Tufik, S. Interactions of polymorphisms in different clock genes associated with circadian phenotypes in humans. Genet. Mol. Biol. 2010, 33, 627–632. [Google Scholar] [CrossRef]
- Dashti, H.S.; Smith, C.E.; Lee, Y.C.; Parnell, L.D.; Lai, C.Q.; Arnett, D.K.; Ordovás, JM.; Garaulet, M. CRY1 circadian gene variant interacts with carbohydrate intake for insulin resistance in two independent populations: Mediterranean and North American. Chronobiol. Int. 2014, 31, 660–667. [Google Scholar] [CrossRef]
- Prasai, M.J.; George, J.T.; Scott, E.M. Molecular clocks, type 2 diabetes and cardiovascular disease. Diabetes Vasc. Dis. Res. 2008, 5, 89–95. [Google Scholar] [CrossRef]
- Martín-Timón, I.; Sevillano-Collantes, C.; Segura-Galindo, A.; Del Cañizo-Gómez, F.J. Type 2 diabetes and cardiovascular disease: Have all risk factors the same strength? World J. Diabetes 2014, 5, 444–470. [Google Scholar] [CrossRef]
- Gómez-Abellán, P.; Hernández-Morante, J.J.; Luján, J.A.; Madrid, J.A.; Garaulet, M. Clock genes are implicated in the human metabolic syndrome. Int. J. Obes. 2008, 32, 121–128. [Google Scholar] [CrossRef]
- Paneni, F.; Beckman, J.A.; Creager, M.A.; Cosentino, F. Diabetes and vascular disease: Pathophysiology, clinical consequences, and medical therapy: Part I. Eur. Heart J. 2013, 34, 2436–2443. [Google Scholar] [CrossRef]
- Dibner, C.; Schibler, U. Circadian timing of metabolism in animal models and humans. J. Intern. Med. 2015, 277, 513–527. [Google Scholar] [CrossRef]
- Tsai, J.Y.; Kienesberger, P.C.; Pulinilkunnil, T.; Sailors, M.H.; Durgan, D.J.; Villegas-Montoya, C.; Jahoor, A.; Gonzalez, R.; Garvey, M.E.; Boland, B.; et al. Direct Regulation of Myocardial Triglyceride Metabolism by the Cardiomyocyte Circadian Clock. J. Biol. Chem. 2010, 285, 2918–2929. [Google Scholar] [CrossRef]
- Paschos, G.K.; Ibrahim, S.; Song, W.L.; Kunieda, T.; Grant, G.; Reyes, T.M.; Bradfield, C.A.; Vaughan, C.H.; Eiden, M.; Masoodi, M.; et al. Obesity in mice with adipocyte-specific deletion of clock component Arntl. Nat. Med. 2012, 18, 1768–1777. [Google Scholar] [CrossRef]
- Woon, P.Y.; Kaisaki, P.J.; Bragança, J.; Bihoreau, M.; Levy, J.C.; Farrall, M.; Gauguier, D. Aryl hydrocarbon receptor nuclear translocator-like (BMAL1) is associated with susceptibility to hypertension and type 2 diabetes. Proc. Natl. Acad. Sci. USA 2007, 104, 14412–14417. [Google Scholar] [CrossRef]
- Škrlec, I.; Milić, J.; Cilenšek, I.; Petrovič, D.; Wagner, J.; Peterlin, B. Circadian clock genes and myocardial infarction in patients with type 2 diabetes mellitus. Gene 2019, 701, 98–103. [Google Scholar] [CrossRef]
- Crnko, S.; Du Pré, B.C.; Sluijter, J.P.G.; Van Laake, L.W. Circadian rhythms and the molecular clock in cardiovascular biology and disease. Nat. Rev. Cardiol. 2019, 16, 437–447. [Google Scholar] [CrossRef]
- Curtis, A.M.; Cheng, Y.; Kapoor, S.; Reilly, D.; Price, T.S.; Fitzgerald, G.A. Circadian variation of blood pressure and the vascular response to asynchronous stress. Proc. Natl. Acad. Sci. USA 2007, 104, 3450–3455. [Google Scholar] [CrossRef]
- Lefta, M.; Campbell, K.S.; Feng, H.Z.; Jin, J.P.; Esser, K.A. Development of dilated cardiomyopathy in Bmal1-deficient mice. Am. J. Physiol.-Heart Circ. Physiol. 2012, 303, H475–H485. [Google Scholar] [CrossRef]
- Corella, D.; Asensio, E.M.; Coltell, O.; Sorlí, J.V.; Estruch, R.; Martínez-González, M.Á.; Salas-Salvadó, J.; Castañer, O.; Arós, F.; Lapetra, J.; et al. CLOCK gene variation is associated with incidence of type-2 diabetes and cardiovascular diseases in type-2 diabetic subjects: Dietary modulation in the PREDIMED randomized trial. Cardiovasc. Diabetol. 2016, 15, 4. [Google Scholar] [CrossRef]
- Riestra, P.; Gebreab, S.Y.; Xu, R.; Khan, R.J.; Gaye, A.; Correa, A.; Min, N.; Sims, M.; Davis, S.K. Circadian CLOCK gene polymorphisms in relation to sleep patterns and obesity in African Americans: Findings from the Jackson heart study. BMC Genet. 2017, 18, 58. [Google Scholar] [CrossRef]
- Bandín, C.; Martinez-Nicolas, A.; Ordovás, J.M.; Ros Lucas, J.A.; Castell, P.; Silvente, T.; Madrid, J.A.; Garaulet, M. Differences in circadian rhythmicity in CLOCK 3111T/C genetic variants in moderate obese women as assessed by thermometry, actimetry and body position. Int. J. Obes. 2013, 37, 1044–1050. [Google Scholar] [CrossRef]
- Garcia-Rios, A.; Gomez-Delgado, F.J.; Garaulet, M.; Alcala-Diaz, J.F.; Delgado-Lista, F.J.; Marin, C.; Rangel-Zuniga, O.A.; Rodriguez-Cantalejo, F.; Gomez-Luna, P.; Ordovas, J.M.; et al. Beneficial effect of CLOCK gene polymorphism rs1801260 in combination with low-fat diet on insulin metabolism in the patients with metabolic syndrome. Chronobiol. Int. 2014, 31, 401–408. [Google Scholar] [CrossRef]
- Suarez-Barrientos, A.; Lopez-Romero, P.; Vivas, D.; Castro-Ferreira, F.; Nunez-Gil, I.; Franco, E.; Ruiz-Mateos, B.; Garcia-Rubira, J.C.; Fernandez-Ortiz, A.; Macaya, C.; et al. Circadian variations of infarct size in acute myocardial infarction. Heart 2011, 97, 970–976. [Google Scholar] [CrossRef]
- Bonney, S.; Kominsky, D.; Brodsky, K.; Eltzschig, H.; Walker, L.; Eckle, T. Cardiac Per2 functions as novel link between fatty acid metabolism and myocardial inflammation during ischemia and reperfusion injury of the heart. PLoS ONE 2013, 8, e71493. [Google Scholar] [CrossRef]
- Vukolic, A.; Antic, V.; Van Vliet, B.N.; Yang, Z.; Albrecht, U.; Montani, J. Role of mutation of the circadian clock gene Per2 in cardiovascular circadian rhythms. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2010, 298, 627–634. [Google Scholar] [CrossRef]
- Bonney, S.; Hughes, K.; Harter, P.N.; Mittelbronn, M.; Walker, L.; Eckle, T. Cardiac period 2 in myocardial ischemia: Clinical implications of a light dependent protein. Int. J. Biochem. Cell Biol. 2013, 45, 667–671. [Google Scholar] [CrossRef]
- Eckle, T.; Hartmann, K.; Bonney, S.; Reithel, S.; Mittelbronn, M.; Walker, L.A.; Lowes, B.D.; Han, J.; Borchers, C.H.; Buttrick, PM.; et al. Adora2b-elicited Per2 stabilization promotes a HIF-dependent metabolic switch crucial for myocardial adaptation to ischemia. Nat. Med. 2012, 18, 774–782. [Google Scholar] [CrossRef]
- Virag, J.A.I.; Dries, J.L.; Easton, P.R.; Friesland, A.M.; Deantonio, J.H.; Chintalgattu, V.; DeAntonio, J.H.; Chintalgattu, V.; Cozzi, E.; Lehmann, B.D.; et al. Attenuation of myocardial injury in mice with functional deletion of the circadian rhythm gene mPer2. Am. J. Physiol. Heart Cric. Physiol. 2010, 198, H1088–H1095. [Google Scholar] [CrossRef]
- Vetter, C.; Devore, E.E.; Wegrzyn, L.R.; Massa, J.; Speizer, F.E.; Kawachi, I.; Rosner, B.; Stampfer, M.J.; Schernhammer, E.S. Association between rotating night shiftwork and risk of coronary heart disease among women. JAMA 2016, 315, 1726–1734. [Google Scholar] [CrossRef]
- Hermansson, J.; Bøggild, H.; Hallqvist, J.; Karlsson, B.; Knutsson, A.; Nilsson, T.; Reuterwall, C.; Gillander Gådin, K. Interaction between shift work and established coronary risk factors. Int. J. Occup. Environ. Med. 2019, 10, 57–65. [Google Scholar] [CrossRef]
- Hermansson, J.; Hallqvist, J.; Karlsson, B.; Knutsson, A.; Gillander Gådin, K. Shift work, parental cardiovascular disease and myocardial infarction in males. Occup. Med. 2018, 68, 120–125. [Google Scholar] [CrossRef]
- Manfredini, R.; Fabbian, F.; Cappadona, R.; De Giorgi, A.; Bravi, F.; Carradori, T.; Flacco, M.E.; Manzoli, L. Daylight Saving Time and Acute Myocardial Infarction: A Meta-Analysis. J. Clin. Med. 2019, 8, 404. [Google Scholar] [CrossRef]
- Kurose, T.; Yabe, D.; Inagaki, N. Circadian rhythms and diabetes. J. Diabetes Investig. 2011, 2, 176–177. [Google Scholar] [CrossRef]
- Allebrandt, K.V.; Roenneberg, T. The search for circadian clock components in humans: New perspectives for association studies. Braz. J. Med. Biol. Res. 2008, 41, 716–721. [Google Scholar] [CrossRef]


{kind=link}
| Gene | SNP | SNP ID | Location | SNP Type |
|---|---|---|---|---|
| ARNTL | rs3789327 | C_2160503_20 | Chromosome 11 | intronic region |
| ARNTL | rs4757144 | C_1870683_20 | Chromosome 11 | intronic region |
| ARNTL | rs12363415 | C_31248677_10 | Chromosome 11 | intronic region |
| CLOCK | rs11932595 | C_296556_10 | Chromosome 4 | intronic region |
| CLOCK | rs6811520 | C_31137409_30 | Chromosome 4 | intronic region |
| CLOCK | rs13124436 | C_11821304_10 | Chromosome 4 | intronic region |
| PER2 | rs35333999 | C_25992030_10 | Chromosome 2 | Coding (Iso/Val) |
| PER2 | rs934945 | C_8740718_20 | Chromosome 2 | Coding (Glu/Gly) |
| MI | no-MI | p-Value * | |
|---|---|---|---|
| Number | 431 | 626 | |
| Male sex (%) | 243 (56.4%) | 321 (51.3%) | 0.102 |
| Age (years) | 66 ±11 | 62 ± 10 | <0.001 |
| History of hypertension (%) | 351 (73.1%) | 408 (65.2%) | 0.007 |
| Smokers (%) | 119 (27.6%) | 193 (30.8%) | <0.001 |
| History of type 2 diabetes mellitus (%) | 275 (63.8%) | 426 (68.1%) | 0.151 |
| Diastolic blood pressure (mm Hg) | 79.12 ± 11.39 | 83.46 ± 11.13 | <0.001 |
| Systolic blood pressure (mm Hg) | 140.04 ± 22.20 | 142.3 ± 21.42 | 0.085 |
| BMI (kg/m2) | 29.78 ± 4.46 | 29.37 ± 4.59 | 0.198 |
| Gene | SNP | Minor Allele | MAF MI | MAF no-MI | p-Value | q-Value |
|---|---|---|---|---|---|---|
| ARNTL | rs3789327 * | G | 0.425 | 0.518 | 2.27 × 10−5 | 6.05 × 10−5 |
| rs4757144 | G | 0.420 | 0.442 | 0.324 | 0.432 | |
| rs12363415 * | A | 0.413 | 0.338 | 4.37 × 10−4 | 8.74 × 10−4 | |
| CLOCK | rs11932595 | G | 0.428 | 0.437 | 0.685 | 0.782 |
| rs6811520 * | C | 0.444 | 0.620 | 7.11 × 10−15 | 5.68 × 10−14 | |
| rs13124436 * | A | 0.200 | 0.301 | 1.72 × 10−7 | 6.88 × 10−7 | |
| PER2 | rs35333999 | T | 0.038 | 0.055 | 0.077 | 0.123 |
| rs934945 | T | 0.172 | 0.174 | 0.885 | 0.885 |
| Gene | SNP | Genotype | Genotype Frequency, N (%) | ||||
|---|---|---|---|---|---|---|---|
| MI | no-MI | p-value | q-Value | ||||
| ARNTL | rs3789327 * | AA | 161 (37.4%) | 164 (26.3%) | 2.7 × 10−4 | 16.46 | 7.2 × 10−4 |
| AG | 174 (40.4%) | 237 (43.8%) | |||||
| GG | 96 (22.3%) | 187 (30%) | |||||
| rs4757144 | AA | 153 (35.5%) | 199 (31.8%) | 0.456 | 1.57 | 0.608 | |
| AG | 194 (45%) | 300 (48%) | |||||
| GG | 84 (19.5%) | 126 (20.2%) | |||||
| rs12363415 * | AA | 142 (32.9%) | 151 (24.1%) | 0.007 | 9.95 | 0.014 | |
| AG | 72 (16.7%) | 121 (19.3%) | |||||
| GG | 217 (50.3%) | 354 (56.5%) | |||||
| CLOCK | rs11932595 | AA | 142 (33%) | 191 (30.6%) | 0.589 | 1.06 | 0.673 |
| AG | 208 (48.4%) | 322 (51.5%) | |||||
| GG | 80 (18.6%) | 112 (17.9%) | |||||
| rs6811520 * | CC | 123 (28.5%) | 255 (40.8%) | 3.44 × 10−15 | 69.33 | 2.75 × 10−14 | |
| CT | 137 (31.8%) | 265 (42.4%) | |||||
| TT | 171 (39.7%) | 105 (16.8%) | |||||
| rs13124436 * | AA | 39 (9%) | 123 (19.6%) | 1.26 × 10−5 | 22.61 | 5.04 × 10−5 | |
| AG | 94 (21.8%) | 131 (20.9%) | |||||
| GG | 298 (69.1%) | 372 (59.4%) | |||||
| PER2 | rs35333999 | CC | 399 (92.8%) | 560 (89.6%) | 0.208 | 3.14 | 0.333 |
| CT | 29 (6.7%) | 61 (9.8%) | |||||
| TT | 2 (0.5%) | 4 (0.6%) | |||||
| rs934945 | CC | 296 (68.7%) | 427 (68.2%) | 0.986 | 0.03 | 0.986 | |
| CT | 122 (28.3%) | 180 (28.8%) | |||||
| TT | 13 (3%) | 19 (3%) | |||||
| Risk Factor | OR (95% CI) | p-Value |
|---|---|---|
| Age | 0.96 (0.95–0.97) | <0.001 |
| History of hypertension | 1.23 (0.91–1.68) | 0.182 |
| Diastolic blood pressure | 1.36 (1.02–1.05) | <0.001 |
| Systolic blood pressure | 1.01 (0.99–1.01) | 0.217 |
| BMI | 0.98 (0.95–1.07) | 0.114 |
| Gene | Sex | Age | History of Hypertension | Smoking | History of T2DM | Diastolic Blood Pressure | Systolic Blood Pressure | BMI |
|---|---|---|---|---|---|---|---|---|
| ARNTL | ||||||||
| rs3789327 | 0.045 | 0.953 | 0.247 | 0.133 | 0.006 | 0.309 | 0.113 | 0.561 |
| rs4757144 | 0.844 | 0.818 | 0.319 | 0.273 | 0.537 | 0.129 | 0.121 | 0.777 |
| rs12363415 | 0.071 | 0.872 | <0.001 | 0.168 | <0.001 | 0.005 | <0.001 | <0.001 |
| CLOCK | ||||||||
| rs11932595 | 0.177 | 0.487 | 0.263 | 0.319 | 0.118 | 0.266 | 0.029 | 0.229 |
| rs6811520 | <0.001 | 0.641 | <0.001 | 0.950 | <0.001 | 0.820 | <0.001 | 0.200 |
| rs13124436 | 0.003 | 0.665 | <0.001 | 0.309 | <0.001 | 0.402 | <0.001 | 0.028 |
| PER2 | ||||||||
| rs35333999 | 0.956 | 0.002 | 0.720 | 0.422 | 0.102 | 0.521 | 0.934 | 0.772 |
| rs934945 | 0.156 | 0.104 | 0.085 | 0.048 | 0.518 | 0.734 | 0.589 | 0.302 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Škrlec, I.; Milić, J.; Steiner, R. The Impact of the Circadian Genes CLOCK and ARNTL on Myocardial Infarction. J. Clin. Med. 2020, 9, 484. https://doi.org/10.3390/jcm9020484
Škrlec I, Milić J, Steiner R. The Impact of the Circadian Genes CLOCK and ARNTL on Myocardial Infarction. Journal of Clinical Medicine. 2020; 9(2):484. https://doi.org/10.3390/jcm9020484
Chicago/Turabian StyleŠkrlec, Ivana, Jakov Milić, and Robert Steiner. 2020. "The Impact of the Circadian Genes CLOCK and ARNTL on Myocardial Infarction" Journal of Clinical Medicine 9, no. 2: 484. https://doi.org/10.3390/jcm9020484
APA StyleŠkrlec, I., Milić, J., & Steiner, R. (2020). The Impact of the Circadian Genes CLOCK and ARNTL on Myocardial Infarction. Journal of Clinical Medicine, 9(2), 484. https://doi.org/10.3390/jcm9020484