Role and Mechanism of LIF in Oral Squamous Cell Carcinoma Progression

Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Lines and Culture
2.2. Transient Transfection and Stable Transfection Clone Selection
2.3. Western Blotting
2.4. Immunohistochemistry
2.5. In Vitro Migration and Invasion Assay
2.6. RNA Isolation
2.7. Reverse Transcription PCR
2.8. Real-Time Quantitative Reverse Transcription PCR
2.9. OSCC Tumor Samples and Clinical Data Collection
2.10. mRNA Microarray Assay
2.11. Statistical Analysis
3. Results
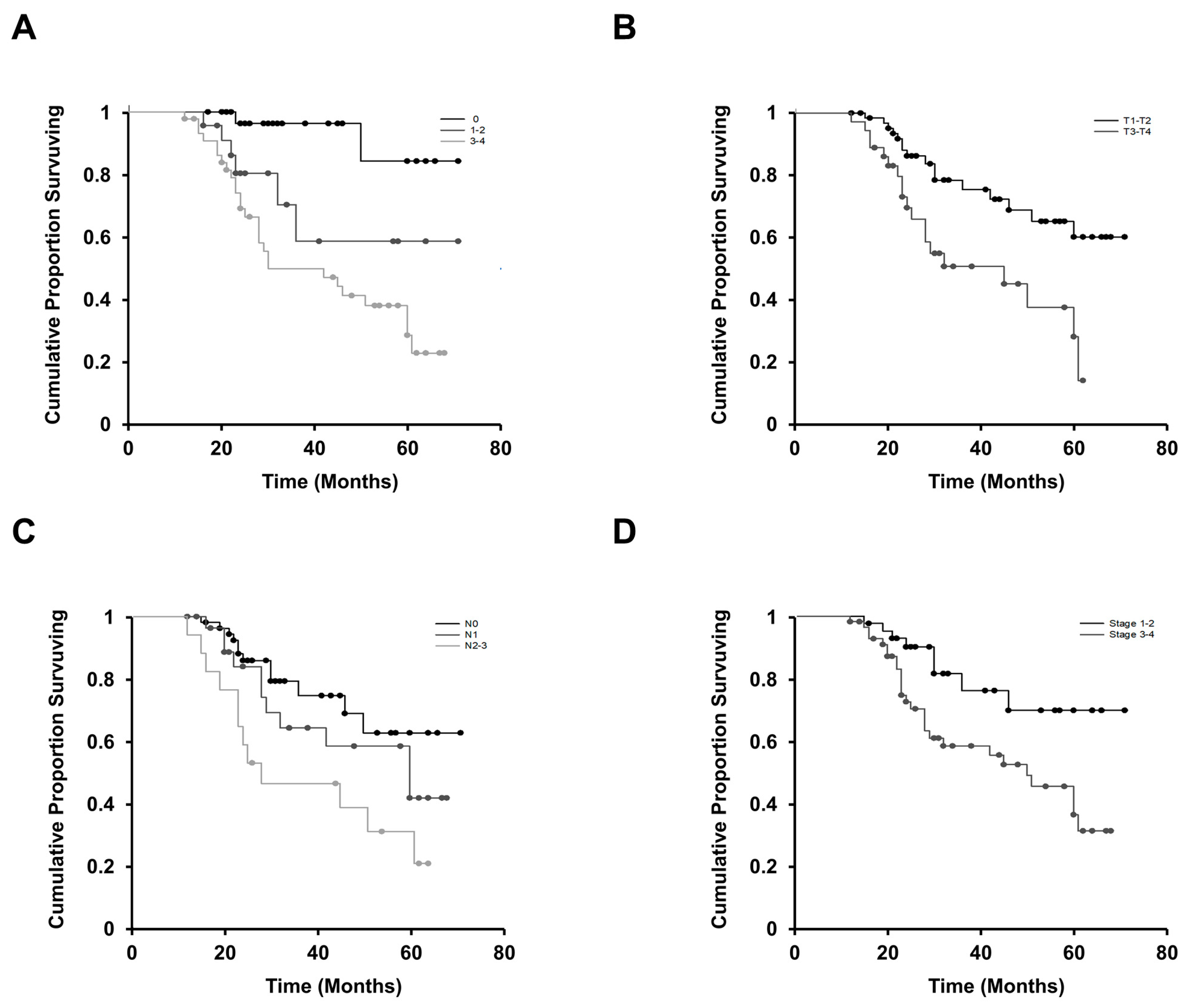
3.1. LIF Affects Regional Lymph Node Involvement and the Advanced Stage of OSCC
3.2. LIF Enhances Cell Migration and Invasion Abilities through Outside-In Signaling in OSCC Cells
3.3. INHBA is a Key Downstream Effector in LIF-Enhanced OSCC Progression
3.4. Relationship between LIF and INHBA Expression in Patients with OSCC
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tian, S.; Jiang, C.; Liu, X.; Xu, S.; Zhang, Z.; Chen, H.; Zhang, Y.; Liu, Y.; Ma, D. Hypermethylation of IFN-gamma in oral cancer tissues. Clin. Oral Investig. 2017, 21, 2535–2542. [Google Scholar] [CrossRef]
- Johnson, N.W.; Jayasekara, P.; Amarasinghe, A.A. Squamous cell carcinoma and precursor lesions of the oral cavity, epidemiology and aetiology. Periodontol. 2000 2011, 57, 19–37. [Google Scholar] [CrossRef]
- Yang, X.; Ruan, H.; Hu, X.; Cao, A.; Song, L. miR-381-3p suppresses the proliferation of oral squamous cell carcinoma cells by directly targeting FGFR2. Am. J. Cancer Res. 2017, 7, 913–922. [Google Scholar]
- Brocklehurst, P.; Kujan, O.; O’Malley, L.A.; Ogden, G.; Shepherd, S.; Glenny, A.M. Screening programmes for the early detection and prevention of oral cancer. Cochrane Database Syst. Rev. 2013, 11, CD004150. [Google Scholar] [CrossRef] [PubMed]
- Hung, K.F.; Lai, K.C.; Liu, T.Y.; Liu, C.J.; Lee, T.C.; Lo, J.F. Asb6 upregulation by Areca nut extracts is associated with betel quid-induced oral carcinogenesis. Oral Oncol. 2009, 45, 543–548. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.L.; Chi, C.W.; Chang, K.W.; Liu, T.Y. Safrole-like DNA adducts in oral tissue from oral cancer patients with a betel quid chewing history. Carcinogenesis 1999, 20, 2331–2334. [Google Scholar] [CrossRef] [PubMed]
- Sasahira, T.; Kirita, T.; Kuniyasu, H. Update of molecular pathobiology in oral cancer: A review. Int. J. Clin. Oncol. 2014, 19, 431–436. [Google Scholar] [CrossRef]
- Li, W.C.; Lee, P.L.; Chou, I.C.; Chang, W.J.; Lin, S.C.; Chang, K.W. Molecular and cellular cues of diet-associated oral carcinogenesis—With an emphasis on areca-nut-induced oral cancer development. J. Oral Pathol. Med. 2015, 44, 167–177. [Google Scholar] [CrossRef]
- Wang, T.Y.; Peng, C.Y.; Lee, S.S.; Chou, M.Y.; Yu, C.C.; Chang, Y.C. Acquisition cancer stemness, mesenchymal transdifferentiation, and chemoresistance properties by chronic exposure of oral epithelial cells to arecoline. Oncotarget 2016, 7, 84072–84081. [Google Scholar] [CrossRef]
- Hsieh, C.H.; Hsu, H.H.; Shibu, M.A.; Day, C.H.; Bau, D.T.; Ho, C.C.; Lin, Y.M.; Chen, M.C.; Wang, S.H.; Huang, C.Y. Down-regulation of beta-catenin and the associated migration ability by Taiwanin C in arecoline and 4-NQO-induced oral cancer cells via GSK-3beta activation. Mol. Carcinog. 2017, 56, 1055–1067. [Google Scholar] [CrossRef]
- Chang, M.C.; Chan, C.P.; Chen, Y.J.; Hsien, H.C.; Chang, Y.C.; Yeung, S.Y.; Jeng, P.Y.; Cheng, R.H.; Hahn, L.J.; Jeng, J.H. Areca nut components stimulate ADAM17, IL-1alpha, PGE2 and 8-isoprostane production in oral keratinocyte: Role of reactive oxygen species, EGF and JAK signaling. Oncotarget 2016, 7, 16879–16894. [Google Scholar] [PubMed]
- Tsai, K.Y.; Su, C.C.; Chiang, C.T.; Tseng, Y.T.; Lian, I.B. Environmental heavy metal as a potential risk factor for the progression of oral potentially malignant disorders in central Taiwan. Cancer Epidemiol. 2017, 47, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Tsai, S.T.; Wong, T.Y.; Ou, C.Y.; Fang, S.Y.; Chen, K.C.; Hsiao, J.R.; Huang, C.C.; Lee, W.T.; Lo, H.I.; Huang, J.S.; et al. The interplay between alcohol consumption, oral hygiene, ALDH2 and ADH1B in the risk of head and neck cancer. Int. J. Cancer 2014, 135, 2424–2436. [Google Scholar] [CrossRef] [PubMed]
- Cheng, E.H.; Liu, J.Y.; Lee, T.H.; Huang, C.C.; Chen, C.I.; Huang, L.S.; Lee, M.S. Requirement of Leukemia Inhibitory Factor or Epidermal Growth Factor for Pre-Implantation Embryogenesis via JAK/STAT3 Signaling Pathways. PLoS ONE 2016, 11, e0153086. [Google Scholar] [CrossRef]
- Smith, A.G.; Nichols, J.; Robertson, M.; Rathjen, P.D. Differentiation inhibiting activity (DIA/LIF) and mouse development. Dev. Biol. 1992, 151, 339–351. [Google Scholar] [CrossRef]
- Gearing, D.P.; Gough, N.M.; King, J.A.; Hilton, D.J.; Nicola, N.A.; Simpson, R.J.; Nice, E.C.; Kelso, A.; Metcalf, D. Molecular cloning and expression of cDNA encoding a murine myeloid leukaemia inhibitory factor (LIF). EMBO J. 1987, 6, 3995–4002. [Google Scholar] [CrossRef]
- Junk, D.J.; Bryson, B.L.; Jackson, M.W. HiJAK’d Signaling; the STAT3 Paradox in Senescence and Cancer Progression. Cancers 2014, 6, 741–755. [Google Scholar] [CrossRef]
- Liu, S.C.; Tsang, N.M.; Chiang, W.C.; Chang, K.P.; Hsueh, C.; Liang, Y.; Juang, J.L.; Chow, K.P.; Chang, Y.S. Leukemia inhibitory factor promotes nasopharyngeal carcinoma progression and radioresistance. J. Clin. Investig. 2013, 123, 5269–5283. [Google Scholar] [CrossRef]
- Morton, S.D.; Cadamuro, M.; Brivio, S.; Vismara, M.; Stecca, T.; Massani, M.; Bassi, N.; Furlanetto, A.; Joplin, R.E.; Floreani, A.; et al. Leukemia inhibitory factor protects cholangiocarcinoma cells from drug-induced apoptosis via a PI3K/AKT-dependent Mcl-1 activation. Oncotarget 2015, 6, 26052–26064. [Google Scholar] [CrossRef]
- Yue, X.; Zhao, Y.; Zhang, C.; Li, J.; Liu, Z.; Liu, J.; Hu, W. Leukemia inhibitory factor promotes EMT through STAT3-dependent miR-21 induction. Oncotarget 2016, 7, 3777–3790. [Google Scholar] [CrossRef]
- Humbert, L.; Ghozlan, M.; Canaff, L.; Tian, J.; Lebrun, J.J. The leukemia inhibitory factor (LIF) and p21 mediate the TGFbeta tumor suppressive effects in human cutaneous melanoma. BMC Cancer 2015, 15, 200. [Google Scholar] [CrossRef] [PubMed]
- Hergovich, A. YAP-Hippo signalling downstream of leukemia inhibitory factor receptor: Implications for breast cancer. Breast Cancer Res. 2012, 14, 326. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Sun, Y.; Wei, Y.; Zhang, P.; Rezaeian, A.H.; Teruya-Feldstein, J.; Gupta, S.; Liang, H.; Lin, H.K.; Hung, M.C.; et al. LIFR is a breast cancer metastasis suppressor upstream of the Hippo-YAP pathway and a prognostic marker. Nat. Med. 2012, 18, 1511–1517. [Google Scholar] [CrossRef] [PubMed]
- Wysoczynski, M.; Miekus, K.; Jankowski, K.; Wanzeck, J.; Bertolone, S.; Janowska-Wieczorek, A.; Ratajczak, J.; Ratajczak, M.Z. Leukemia inhibitory factor: A newly identified metastatic factor in rhabdomyosarcomas. Cancer Res. 2007, 67, 2131–2140. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Yue, X.; Zhao, Y.; Li, X.; Wu, L.; Zhang, C.; Liu, Z.; Lin, K.; Xu-Monette, Z.Y.; Young, K.H.; et al. LIF negatively regulates tumour-suppressor p53 through Stat3/ID1/MDM2 in colorectal cancers. Nat. Commun. 2014, 5, 5218. [Google Scholar] [CrossRef]
- Li, X.; Yang, Q.; Yu, H.; Wu, L.; Zhao, Y.; Zhang, C.; Yue, X.; Liu, Z.; Wu, H.; Haffty, B.G.; et al. LIF promotes tumorigenesis and metastasis of breast cancer through the AKT-mTOR pathway. Oncotarget 2014, 5, 788–801. [Google Scholar] [CrossRef]
- Vale, W.; Rivier, J.; Vaughan, J.; McClintock, R.; Corrigan, A.; Woo, W.; Karr, D.; Spiess, J. Purification and characterization of an FSH releasing protein from porcine ovarian follicular fluid. Nature 1986, 321, 776–779. [Google Scholar] [CrossRef]
- Bufalino, A.; Cervigne, N.K.; de Oliveira, C.E.; Fonseca, F.P.; Rodrigues, P.C.; Macedo, C.C.; Sobral, L.M.; Miguel, M.C.; Lopes, M.A.; Paes Leme, A.F.; et al. Low miR-143/miR-145 Cluster Levels Induce Activin A Overexpression in Oral Squamous Cell Carcinomas, Which Contributes to Poor Prognosis. PLoS ONE 2015, 10, e0136599. [Google Scholar] [CrossRef]
- Lee, H.Y.; Li, C.C.; Huang, C.N.; Li, W.M.; Yeh, H.C.; Ke, H.L.; Yang, K.F.; Liang, P.I.; Li, C.F.; Wu, W.J. INHBA overexpression indicates poor prognosis in urothelial carcinoma of urinary bladder and upper tract. J. Surg. Oncol. 2015, 111, 414–422. [Google Scholar] [CrossRef]
- Luo, Q.; Wang, C.; Jin, G.; Gu, D.; Wang, N.; Song, J.; Jin, H.; Hu, F.; Zhang, Y.; Ge, T.; et al. LIFR functions as a metastasis suppressor in hepatocellular carcinoma by negatively regulating phosphoinositide 3-kinase/AKT pathway. Carcinogenesis 2015, 36, 1201–1212. [Google Scholar] [CrossRef]
- Shen, L.; Shi, Q.; Wang, W. Double agents: Genes with both oncogenic and tumor-suppressor functions. Oncogenesis 2018, 7, 25. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Han, Y.; Suarez Saiz, F.; Minden, M.D. A tumor suppressor and oncogene: The WT1 story. Leukemia 2007, 21, 868–876. [Google Scholar] [CrossRef] [PubMed]
- Zamarron, B.F.; Chen, W. Dual roles of immune cells and their factors in cancer development and progression. Int. J. Biol. Sci. 2011, 7, 651–658. [Google Scholar] [CrossRef] [PubMed]
- Carafa, V.; Altucci, L.; Nebbioso, A. Dual Tumor Suppressor and Tumor Promoter Action of Sirtuins in Determining Malignant Phenotype. Front. Pharmacol. 2019, 10, 38. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Powers, S.; Zhu, W.; Hannun, Y.A. Substantial contribution of extrinsic risk factors to cancer development. Nature 2016, 529, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Reya, T.; Morrison, S.J.; Clarke, M.F.; Weissman, I.L. Stem cells, cancer, and cancer stem cells. Nature 2001, 414, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Shrihari, T.G. Dual role of inflammatory mediators in cancer. Ecancermedicalscience 2017, 11, 721. [Google Scholar] [CrossRef]
- Ohata, Y.; Tsuchiya, M.; Hirai, H.; Yamaguchi, S.; Akashi, T.; Sakamoto, K.; Yamaguchi, A.; Ikeda, T.; Kayamori, K. Leukemia inhibitory factor produced by fibroblasts within tumor stroma participates in invasion of oral squamous cell carcinoma. PLoS ONE 2018, 13, e0191865. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinicopathological Parameters | Degree of LIF Staining | |||
|---|---|---|---|---|
| 0 (n = 32) | 1 or 2 (n = 22) | 3 or 4 (n = 46) | p-Value a | |
| Patient’s ages (years) | ||||
| <50 | 7 (21.9%) | 6 (27.3%) | 11 (23.9%) | 0.828 |
| 50–59 | 18 (56.2%) | 9 (40.9%) | 19 (41.3%) | |
| 60–69 | 4 (12.5%) | 5 (22.7%) | 9 (19.6%) | |
| ≥70 | 3 (9.4%) | 2 (9.1%) | 7 (15.2%) | |
| Patient’s sex | ||||
| Male | 28 (87.5%) | 19 (95.5%) | 44 (95.6%) | 0.335 |
| Female | 4 (12.5%) | 3 (4.5%) | 2 (4.4%) | |
| Cancer locations | ||||
| Buccal and lip SCC | 16 (50%) | 10 (45.5%) | 22 (47.8%) | 0.526 b |
| Gingival SCC | 5 (15.6%) | 5 (22.7%) | 2 (4.3%) | |
| Mouth floor SCC | 0 (0%) | 0 (0%) | 1 (2.2%) | |
| Palate SCC | 1 (3.1%) | 1 (4.5%) | 3 (6.5%) | |
| Tongue SCC | 10 (31.3%) | 6 (27.3%) | 18 (39.1%) | |
| T status | ||||
| T1–T2 | 22 (68.8%) | 19 (86.4%) | 23 (50%) | 0.051 |
| T3–T4 | 10 (31.2%) | 3 (13.6%) | 23 (50%) | |
| N status | ||||
| N0 | 23 (71.9%) | 12 (54.5%) | 18 (39.1%) | 0.022 * |
| N1 | 6 (18.8%) | 4 (18.2%) | 20 (43.5%) | |
| N2–N3 | 3 (9.3%) | 6 (27.3%) | 8 (17.4%) | |
| Clinical staging | ||||
| Stage 1 | 10 (31.25%) | 5 (22.7%) | 5 (10.9%) | 0.022 * |
| Stage 2 | 10 (31.25%) | 7 (31.8%) | 6 (13.0%) | |
| Stage 3 | 2 (6.25%) | 3 (13.6%) | 13 (28.3%) | |
| Stage 4 | 10 (31.25%) | 7 (31.8%) | 22 (47.8%) | |
| Stages 1–2 | 20 (62.5%) | 12 (54.5%) | 11 (23.9%) | 0.002 * |
| Stages 3–4 | 12 (37.5%) | 10 (45.5%) | 35 (76.1%) | |
| Histological differentiation | ||||
| Well-diff. SCC | 30 (93.8%) | 20 (90.1%) | 41 (89.1%) | 0.773 |
| Moderately-diff. SCC | 1 (3.1%) | 2 (9.9%) | 3 (6.5%) | |
| Poorly-diff. SCC | 1 (3.1%) | 0 (0.0%) | 2 (4.4%) | |
| Depth of invasion (DOI) | ||||
| <5 mm | 25 (78.1%) | 13 (59.1%) | 9 (19.6%) | 0.001 * |
| 5–9 mm | 5 (15.6%) | 5 (22.7%) | 29 (63.0%) | |
| >9 mm | 2 (6.3%) | 4 (18.2%) | 8 (17.4%) | |
| Margin status | ||||
| ≥5 mm | 29 (90.6%) | 17 (72.3%) | 27 (58.7%) | 0.023 * |
| <5 mm | 1 (3.1%) | 4 (18.2%) | 15 (32.6%) | |
| Involved | 2 (6.3%) | 1 (4.5%) | 4 (8.7%) | |
| Perineural invasion | ||||
| No | 27 (84.4%) | 18 (81.8%) | 36 (78.3%) | 0.79 |
| Yes | 5 (15.6%) | 4 (18.2%) | 10 (21.7%) | |
| Factor | Hazard Ratio (95% CI) | p-Value a |
|---|---|---|
| Univariate | ||
| Cancer locations (palatal vs. buccal and lip) | 4.95 (0.92–26.88) | 0.062 |
| Cancer locations (palatal vs. gingival) | 3.52 (0.46–26.73) | 0.22 |
| Cancer locations (palatal vs. tongue) | 4.18 (0.53–32.68) | 0.172 |
| T status (T3 + T4 vs. T1 + T2) | 1.44 (0.30–6.94) | 0.217 |
| N status (N2 + N3 vs. N0) | 2.78 (0.37–20.45) | 0.314 |
| N status (N2 + N3 vs. N1) | 2.91 (1.04–8.06) | 0.041 * |
| Clinical staging (stages 3 + 4 vs. 1 + 2) | 0.48 (0.06–3.64) | 0.481 |
| Histological differentiation (poor vs. well) | 13.10 (1.35–127.41) | 0.027 * |
| Histological differentiation (moderate vs. well) | 1.17 (0.31–4.33) | 0.81 |
| DOI (>10 mm vs. 5–9 mm) | 5.43 (1.58–18.66) | 0.007 * |
| DOI (>10 mm vs. <5 mm) | 16.08 (2.96–87.71) | 0.001 * |
| Margin status (margin involved vs. A > 5 mm) | 1.88 (0.38–9.26) | 0.435 |
| Margin status (margin involved vs. closed < 5 mm) | 4.95 (0.71–34.48) | 0.104 |
| PNI (negative vs. positive) | 0.87 (0.28–2.72) | 0.817 |
| LIF label index (3 + 4 vs. 0) | 8.84 (5.71–136.99) | 0.001 * |
| LIF label index (3 + 4 vs. 1 + 2) | 1.36 (0.0–2.7) | 0.05 |
| Multivariate | ||
| LIF Label index (3 + 4 vs. 0) | 6.83 (2.88–133.68) | 0.026 * |
| Clinicopathological Parameters | Degree of LIF Staining | |||
|---|---|---|---|---|
| 0 (n = 32) | 1 or 2 (n = 22) | 3 or 4 (n = 46) | p-Value a | |
| Daily alcohol consumption | ||||
| Nondrinkers | 6 (18.8%) | 5 (22.7%) | 10 (21.7%) | 0.997 |
| ≤3500 mL | 17 (53.1%) | 11 (50%) | 23 (50.00%) | |
| >3500 mL | 9 (28.1%) | 6 (27.3%) | 13 (28.3%) | |
| Duration of drinking alcohol | ||||
| Nondrinkers | 6 (18.8%) | 5 (22.7%) | 10 (21.7%) | 0.794 |
| ≤10 years | 7 (21.9%) | 7 (31.8%) | 9 (19.6%) | |
| >10 years | 19 (59.3%) | 10 (45.5%) | 27 (58.7%) | |
| Daily AQ consumption | ||||
| Nonchewers | 10 (31.3%) | 4 (18.2%) | 5 (10.9%) | 0.176 |
| ≤10 quids | 10 (31.2%) | 7 (31.8%) | 22 (47.8%) | |
| >10 quids | 12 (37.5%) | 11 (50%) | 19 (41.3%) | |
| Duration of chewing AQs | ||||
| Nonchewers | 10 (31.3%) | 4 (18.2%) | 5 (10.9%) | 0.219 |
| ≤10 years | 5 (15.6%) | 6 (27.3%) | 13 (28.3%) | |
| >10 years | 17 (53.1%) | 12 (54.5%) | 28 (60.8%) | |
| Daily cigarette consumption | ||||
| Nonsmokers | 6 (18.8%) | 3 (13.6%) | 8 (17.4%) | 0.487 |
| ≤1 pack | 17 (53.1%) | 15 (68.2%) | 21 (45.6%) | |
| >1 pack | 9 (28.1%) | 4 (18.2%) | 17 (37.0%) | |
| Duration of smoking | ||||
| Nonsmokers | 6 (18.8%) | 3 (13.6%) | 8 (17.4%) | 0.092 |
| ≤10 years | 2 (6.2%) | 2 (9.1%) | 4 (8.7%) | |
| >10 years | 24 (75%) | 17 (77.2%) | 34 (73.9%) | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, T.-A.; Wu, T.-S.; Li, Y.-J.; Yang, C.-N.; Illescas Ralda, M.M.; Chang, H.-H. Role and Mechanism of LIF in Oral Squamous Cell Carcinoma Progression. J. Clin. Med. 2020, 9, 295. https://doi.org/10.3390/jcm9020295
Lin T-A, Wu T-S, Li Y-J, Yang C-N, Illescas Ralda MM, Chang H-H. Role and Mechanism of LIF in Oral Squamous Cell Carcinoma Progression. Journal of Clinical Medicine. 2020; 9(2):295. https://doi.org/10.3390/jcm9020295
Chicago/Turabian StyleLin, Ting-An, Tai-Sheng Wu, Yue-Ju Li, Cheng-Ning Yang, Monica Maria Illescas Ralda, and Hao-Hueng Chang. 2020. "Role and Mechanism of LIF in Oral Squamous Cell Carcinoma Progression" Journal of Clinical Medicine 9, no. 2: 295. https://doi.org/10.3390/jcm9020295
APA StyleLin, T.-A., Wu, T.-S., Li, Y.-J., Yang, C.-N., Illescas Ralda, M. M., & Chang, H.-H. (2020). Role and Mechanism of LIF in Oral Squamous Cell Carcinoma Progression. Journal of Clinical Medicine, 9(2), 295. https://doi.org/10.3390/jcm9020295