A Novel Liquid Biopsy Strategy to Detect Small Amounts of Cancer Cells Using Cancer-Specific Replication Adenoviruses



Abstract
1. Introduction
2. Detection of CTCs in Gynecological Cancer Using Conventional Technologies and Their Clinical Significance
2.1. Endometrial Cancer
2.2. Cervical Cancer
2.3. Ovarian Cancer
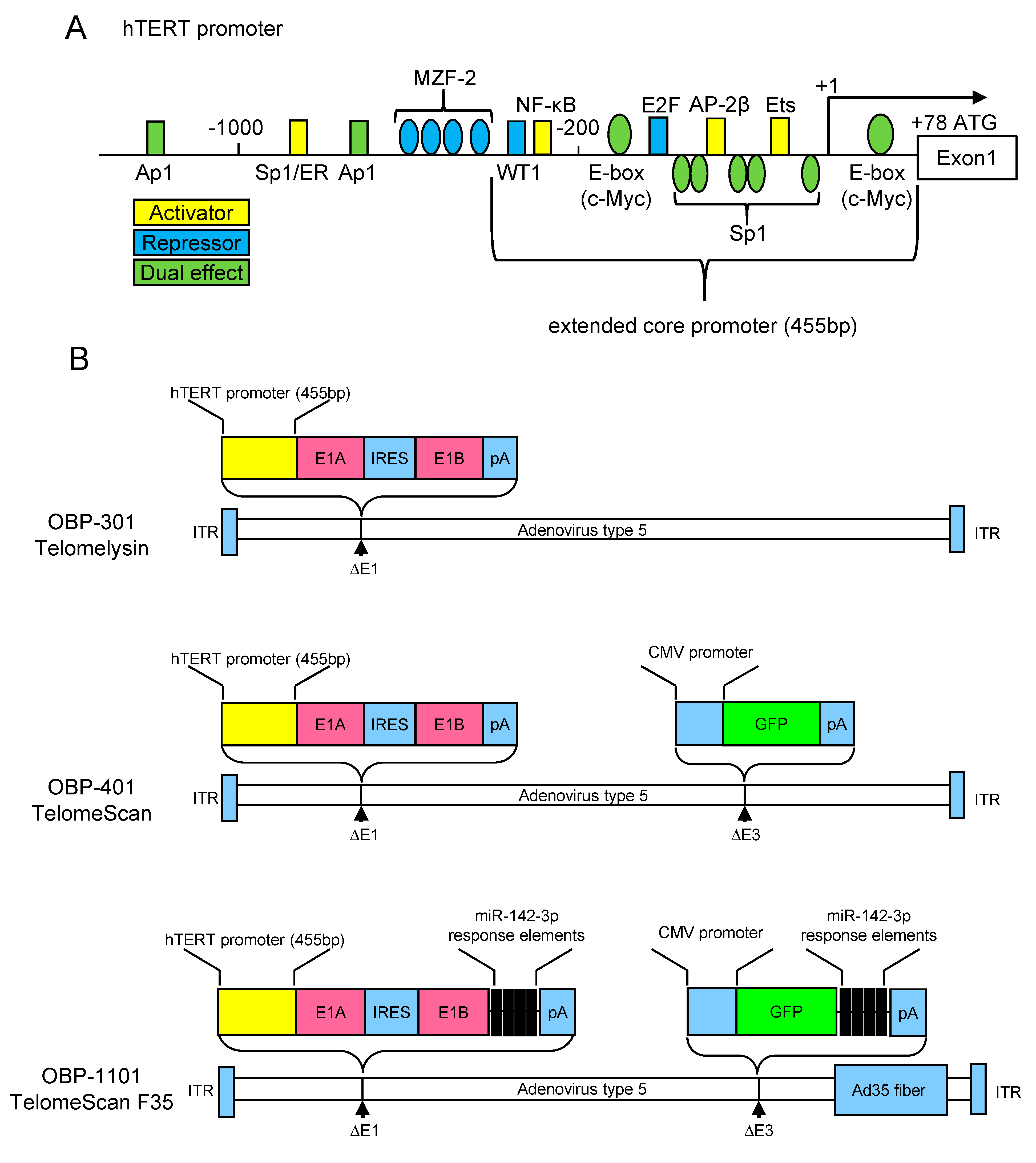
3. The hTERT Promoter Activity as Hallmark of the Cancer Cells
4. Development of Telomerase-Specific Replication Adenovirus and Its Application for Diagnostic Use
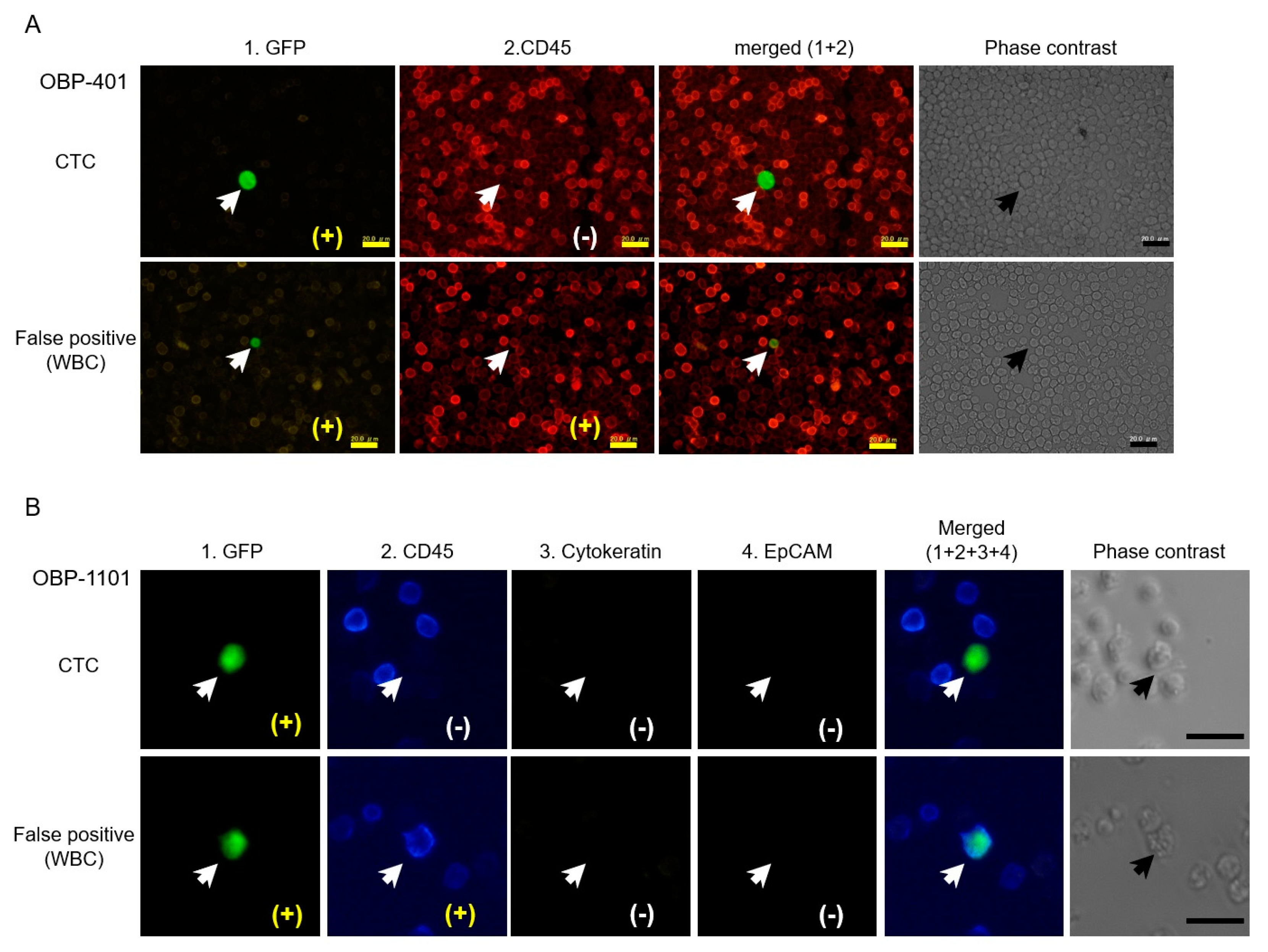
5. Detection of Telomerase-Specific Replication Adenovirus to Detect CTCs in Gynecological Cancers
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Hu, Y.; An, Q.; Sheu, K.; Trejo, B.; Fan, S.; Guo, Y. Single Cell Multi-Omics Technology: Methodology and Application. Front. Cell Dev. Biol. 2018, 6, 28. [Google Scholar] [CrossRef] [PubMed]
- Allard, W.J.; Matera, J.; Miller, M.C.; Repollet, M.; Connelly, M.C.; Rao, C.; Tibbe, A.G.J.; Uhr, J.W.; Terstappen, L.W.M.M. Tumor cells circulate in the peripheral blood of all major carcinomas but not in healthy subjects or patients with nonmalignant diseases. Clin. Cancer Res. 2004, 10, 6897–6904. [Google Scholar] [CrossRef] [PubMed]
- Zhu, P.; Stanton, M.L.; Castle, E.P.; Joseph, R.W.; Adams, D.L.; Li, S.; Amstutz, P.; Tang, C.-M.; Ho, T.H. Detection of tumor-associated cells in cryopreserved peripheral blood mononuclear cell samples for retrospective analysis. J. Transl. Med. 2016, 14, 198. [Google Scholar] [CrossRef] [PubMed]
- Adams, D.L.; Adams, D.K.; Alpaugh, R.K.; Cristofanilli, M.; Martin, S.S.; Chumsri, S.; Tang, C.-M.; Marks, J.R. Circulating Cancer-Associated Macrophage-Like Cells Differentiate Malignant Breast Cancer and Benign Breast Conditions. Cancer Epidemiol. Biomarkers Prev. 2016, 25, 1037–1042. [Google Scholar] [CrossRef]
- Kou, R.; Zhao, J.; Gogoi, P.; Carskadon, S.; Chow, W.; Hwang, C.; Palanisamy, N.; Leung, C.; Wang, Y. Enrichment and mutation detection of circulating tumor cells from blood samples. Oncol. Rep. 2018, 39, 2537–2544. [Google Scholar] [CrossRef]
- Fabbri, F.; Carloni, S.; Zoli, W.; Ulivi, P.; Gallerani, G.; Fici, P.; Chiadini, E.; Passardi, A.; Frassineti, G.L.; Ragazzini, A.; et al. Detection and recovery of circulating colon cancer cells using a dielectrophoresis-based device: KRAS mutation status in pure CTCs. Cancer Lett. 2013, 335, 225–231. [Google Scholar] [CrossRef]
- Jie, X.-X.; Zhang, X.-Y.; Xu, C.-J. Epithelial-to-mesenchymal transition, circulating tumor cells and cancer metastasis: Mechanisms and clinical applications. Oncotarget 2017, 8, 81558–81571. [Google Scholar] [CrossRef]
- Gorges, T.M.; Tinhofer, I.; Drosch, M.; Röse, L.; Zollner, T.M.; Krahn, T.; von Ahsen, O. Circulating tumour cells escape from EpCAM-based detection due to epithelial-to-mesenchymal transition. BMC Cancer 2012, 12, 178. [Google Scholar] [CrossRef]
- Wang, L.; Li, Y.; Xu, J.; Zhang, A.; Wang, X.; Tang, R.; Zhang, X.; Yin, H.; Liu, M.; Wang, D.D.; et al. Quantified postsurgical small cell size CTCs and EpCAM+ circulating tumor stem cells with cytogenetic abnormalities in hepatocellular carcinoma patients determine cancer relapse. Cancer Lett. 2018, 412, 99–107. [Google Scholar] [CrossRef]
- Hayflick, L. Mortality and immortality at the cellular level. A review. Biochemistry 1997, 62, 1180–1190. [Google Scholar]
- Kim, N.W.; Piatyszek, M.A.; Prowse, K.R.; Harley, C.B.; West, M.D.; Ho, P.L.; Coviello, G.M.; Wright, W.E.; Weinrich, S.L.; Shay, J.W. Specific association of human telomerase activity with immortal cells and cancer. Science 1994, 266, 2011–2015. [Google Scholar] [CrossRef] [PubMed]
- Shay, J.W.; Bacchetti, S. A survey of telomerase activity in human cancer. Eur. J. Cancer 1997, 33, 787–791. [Google Scholar] [CrossRef]
- Feng, J.; Funk, W.D.; Wang, S.S.; Weinrich, S.L.; Avilion, A.A.; Chiu, C.P.; Adams, R.R.; Chang, E.; Allsopp, R.C.; Yu, J. The RNA component of human telomerase. Science 1995, 269, 1236–1241. [Google Scholar] [CrossRef] [PubMed]
- Harrington, L.; McPhail, T.; Mar, V.; Zhou, W.; Oulton, R.; Bass, M.B.; Arruda, I.; Robinson, M.O. A mammalian telomerase-associated protein. Science 1997, 275, 973–977. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.M.; Morin, G.B.; Chapman, K.B.; Weinrich, S.L.; Andrews, W.H.; Lingner, J.; Harley, C.B.; Cech, T.R. Telomerase catalytic subunit homologs from fission yeast and human. Science 1997, 277, 955–959. [Google Scholar] [CrossRef] [PubMed]
- Meyerson, M.; Counter, C.M.; Eaton, E.N.; Ellisen, L.W.; Steiner, P.; Caddle, S.D.; Ziaugra, L.; Beijersbergen, R.L.; Davidoff, M.J.; Liu, Q.; et al. hEST2, the putative human telomerase catalytic subunit gene, is up-regulated in tumor cells and during immortalization. Cell 1997, 90, 785–795. [Google Scholar] [CrossRef]
- Takakura, M.; Kyo, S.; Kanaya, T.; Hirano, H.; Takeda, J.; Yutsudo, M.; Inoue, M. Cloning of human telomerase catalytic subunit (hTERT) gene promoter and identification of proximal core promoter sequences essential for transcriptional activation in immortalized and cancer cells. Cancer Res. 1999, 59, 551–557. [Google Scholar]
- Horikawa, I.; Cable, P.L.; Afshari, C.; Barrett, J.C. Cloning and characterization of the promoter region of human telomerase reverse transcriptase gene. Cancer Res. 1999, 59, 826–830. [Google Scholar]
- Cong, Y.S.; Wen, J.; Bacchetti, S. The human telomerase catalytic subunit hTERT: Organization of the gene and characterization of the promoter. Hum. Mol. Genet. 1999, 8, 137–142. [Google Scholar] [CrossRef]
- Kiss, I.; Kolostova, K.; Matkowski, R.; Jędryka, M.; Czekański, A.; Pavlasek, J.; Bobek, V. Correlation Between Disease Stage and the Presence of Viable Circulating Tumor Cells in Endometrial Cancer. Anticancer Res. 2018, 38, 2983–2987. [Google Scholar] [CrossRef]
- Bogani, G.; Liu, M.C.; Dowdy, S.C.; Cliby, W.A.; Kerr, S.E.; Kalli, K.R.; Kipp, B.R.; Halling, K.C.; Campion, M.B.; Mariani, A. Detection of circulating tumor cells in high-risk endometrial cancer. Anticancer Res. 2015, 35, 683–687. [Google Scholar] [PubMed]
- Ni, T.; Sun, X.; Shan, B.; Wang, J.; Liu, Y.; Gu, S.-L.; Wang, Y.-D. Detection of circulating tumour cells may add value in endometrial cancer management. Eur. J. Obstet. Gynecol. Reprod. Biol. 2016, 207, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Lemech, C.R.; Ensell, L.; Paterson, J.C.; Eminowicz, G.; Lowe, H.; Arora, R.; Arkenau, H.-T.; Widschwendter, M.; MacDonald, N.; Olaitan, A.; et al. Enumeration and Molecular Characterisation of Circulating Tumour Cells in Endometrial Cancer. Oncology 2016, 91, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Casas-Arozamena, C.; Díaz, E.; Moiola, C.P.; Alonso-Alconada, L.; Ferreiros, A.; Abalo, A.; López Gil, C.; Oltra, S.S.; de Santiago, J.; Cabrera, S.; et al. Genomic Profiling of Uterine Aspirates and cfDNA as an Integrative Liquid Biopsy Strategy in Endometrial Cancer. J. Clin. Med. 2020, 9, 585. [Google Scholar] [CrossRef] [PubMed]
- Obermayr, E.; Sanchez-Cabo, F.; Tea, M.-K.M.; Singer, C.F.; Krainer, M.; Fischer, M.B.; Sehouli, J.; Reinthaller, A.; Horvat, R.; Heinze, G.; et al. Assessment of a six gene panel for the molecular detection of circulating tumor cells in the blood of female cancer patients. BMC Cancer 2010, 10, 666. [Google Scholar] [CrossRef] [PubMed]
- Obermayr, E.; Maritschnegg, E.; Agreiter, C.; Pecha, N.; Speiser, P.; Helmy-Bader, S.; Danzinger, S.; Krainer, M.; Singer, C.; Zeillinger, R. Efficient leukocyte depletion by a novel microfluidic platform enables the molecular detection and characterization of circulating tumor cells. Oncotarget 2018, 9, 812–823. [Google Scholar] [CrossRef] [PubMed]
- Tewari, K.S.; Sill, M.W.; Monk, B.J.; Penson, R.T.; Moore, D.H.; Lankes, H.A.; Ramondetta, L.M.; Landrum, L.M.; Randall, L.M.; Oaknin, A.; et al. Circulating Tumor Cells In Advanced Cervical Cancer: NRG Oncology—Gynecologic Oncology Group Study 240 (NCT 00803062). Mol. Cancer Ther. 2020. [Google Scholar] [CrossRef]
- Pfitzner, C.; Schröder, I.; Scheungraber, C.; Dogan, A.; Runnebaum, I.B.; Dürst, M.; Häfner, N. Digital-Direct-RT-PCR: A sensitive and specific method for quantification of CTC in patients with cervical carcinoma. Sci. Rep. 2014, 4, 3970. [Google Scholar] [CrossRef]
- Wen, Y.-F.; Cheng, T.-T.; Chen, X.-L.; Huang, W.-J.; Peng, H.-H.; Zhou, T.-C.; Lin, X.-D.; Zeng, L.-S. Elevated circulating tumor cells and squamous cell carcinoma antigen levels predict poor survival for patients with locally advanced cervical cancer treated with radiotherapy. PLoS ONE 2018, 13, e0204334. [Google Scholar] [CrossRef]
- Du, K.; Huang, Q.; Bu, J.; Zhou, J.; Huang, Z.; Li, J. Circulating Tumor Cells Counting Act as a Potential Prognostic Factor in Cervical Cancer. Technol. Cancer Res. Treat. 2020, 19, 1533033820957005. [Google Scholar] [CrossRef]
- Poveda, A.; Kaye, S.B.; McCormack, R.; Wang, S.; Parekh, T.; Ricci, D.; Lebedinsky, C.A.; Tercero, J.C.; Zintl, P.; Monk, B.J. Circulating tumor cells predict progression free survival and overall survival in patients with relapsed/recurrent advanced ovarian cancer. Gynecol. Oncol. 2011, 122, 567–572. [Google Scholar] [CrossRef] [PubMed]
- Behbakht, K.; Sill, M.W.; Darcy, K.M.; Rubin, S.C.; Mannel, R.S.; Waggoner, S.; Schilder, R.J.; Cai, K.Q.; Godwin, A.K.; Alpaugh, R.K. Phase II trial of the mTOR inhibitor, temsirolimus and evaluation of circulating tumor cells and tumor biomarkers in persistent and recurrent epithelial ovarian and primary peritoneal malignancies: A Gynecologic Oncology Group study. Gynecol. Oncol. 2011, 123, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.F.; Kindelberger, D.; Doyle, C.; Lowe, A.; Barry, W.T.; Matulonis, U.A. Predictive value of circulating tumor cells (CTCs) in newly-diagnosed and recurrent ovarian cancer patients. Gynecol. Oncol. 2013, 131, 352–356. [Google Scholar] [CrossRef] [PubMed]
- Banys-Paluchowski, M.; Fehm, T.; Neubauer, H.; Paluchowski, P.; Krawczyk, N.; Meier-Stiegen, F.; Wallach, C.; Kaczerowsky, A.; Gebauer, G. Clinical relevance of circulating tumor cells in ovarian, fallopian tube and peritoneal cancer. Arch. Gynecol. Obstet. 2020, 301, 1027–1035. [Google Scholar] [CrossRef] [PubMed]
- Aktas, B.; Kasimir-Bauer, S.; Heubner, M.; Kimmig, R.; Wimberger, P. Molecular Profiling and Prognostic Relevance of Circulating Tumor Cells in the Blood of Ovarian Cancer Patients at Primary Diagnosis and After Platinum-Based Chemotherapy. Int. J. Gynecol. Cancer 2011, 21. [Google Scholar] [CrossRef]
- Kuhlmann, J.D.; Wimberger, P.; Bankfalvi, A.; Keller, T.; Schöler, S.; Aktas, B.; Buderath, P.; Hauch, S.; Otterbach, F.; Kimmig, R.; et al. ERCC1-Positive Circulating Tumor Cells in the Blood of Ovarian Cancer Patients as a Predictive Biomarker for Platinum Resistance. Clin. Chem. 2014, 60, 1282–1289. [Google Scholar] [CrossRef]
- Chebouti, I.; Kuhlmann, J.D.; Buderath, P.; Weber, S.; Wimberger, P.; Bokeloh, Y.; Hauch, S.; Kimmig, R.; Kasimir-Bauer, S. ERCC1-expressing circulating tumor cells as a potential diagnostic tool for monitoring response to platinum-based chemotherapy and for predicting post-therapeutic outcome of ovarian cancer. Oncotarget 2017, 8, 24303–24313. [Google Scholar] [CrossRef]
- Chebouti, I.; Kasimir-Bauer, S.; Buderath, P.; Wimberger, P.; Hauch, S.; Kimmig, R.; Kuhlmann, J.D. EMT-like circulating tumor cells in ovarian cancer patients are enriched by platinum-based chemotherapy. Oncotarget 2017, 8, 48820–48831. [Google Scholar] [CrossRef]
- Kolostova, K.; Matkowski, R.; Jędryka, M.; Soter, K.; Cegan, M.; Pinkas, M.; Jakabova, A.; Pavlasek, J.; Spicka, J.; Bobek, V. The added value of circulating tumor cells examination in ovarian cancer staging. Am. J. Cancer Res. 2015, 5, 3363–3375. [Google Scholar]
- Kolostova, K.; Pinkas, M.; Jakabova, A.; Pospisilova, E.; Svobodova, P.; Spicka, J.; Cegan, M.; Matkowski, R.; Bobek, V. Molecular characterization of circulating tumor cells in ovarian cancer. Am. J. Cancer Res. 2016, 6, 973–980. [Google Scholar]
- Obermayr, E.; Bednarz-Knoll, N.; Orsetti, B.; Weier, H.-U.; Lambrechts, S.; Castillo-Tong, D.C.; Reinthaller, A.; Braicu, E.I.; Mahner, S.; Sehouli, J.; et al. Circulating tumor cells: Potential markers of minimal residual disease in ovarian cancer? A study of the OVCAD consortium. Oncotarget 2017, 8, 106415–106428. [Google Scholar] [CrossRef]
- Zhang, X.; Li, H.; Yu, X.; Li, S.; Lei, Z.; Li, C.; Zhang, Q.; Han, Q.; Li, Y.; Zhang, K.; et al. Analysis of Circulating Tumor Cells in Ovarian Cancer and Their Clinical Value as a Biomarker. CPB 2018, 48, 1983–1994. [Google Scholar] [CrossRef] [PubMed]
- Witek, M.A.; Aufforth, R.D.; Wang, H.; Kamande, J.W.; Jackson, J.M.; Pullagurla, S.R.; Hupert, M.L.; Usary, J.; Wysham, W.Z.; Hilliard, D.; et al. Discrete microfluidics for the isolation of circulating tumor cell subpopulations targeting fibroblast activation protein alpha and epithelial cell adhesion molecule. NPJ Precis. Oncol. 2017, 1. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Kim, E.J.; Cho, Y.; Kim, S.; Chung, H.H.; Park, N.H.; Song, Y.-S. Predictive value of circulating tumor cells (CTCs) captured by microfluidic device in patients with epithelial ovarian cancer. Gynecol. Oncol. 2017, 145, 361–365. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Lim, M.; Kim, J.Y.; Shin, S.-J.; Cho, Y.-K.; Cho, C.H. Circulating Tumor Cells Enumerated by a Centrifugal Microfluidic Device as a Predictive Marker for Monitoring Ovarian Cancer Treatment: A Pilot Study. Diagnostics 2020, 10, 249. [Google Scholar] [CrossRef] [PubMed]
- Dorsey, J.F.; Kao, G.D.; MacArthur, K.M.; Ju, M.; Steinmetz, D.; Wileyto, E.P.; Simone, C.B.; Hahn, S.M. Tracking viable circulating tumor cells (CTCs) in the peripheral blood of non-small cell lung cancer (NSCLC) patients undergoing definitive radiation therapy: Pilot study results. Cancer 2015, 121, 139–149. [Google Scholar] [CrossRef]
- Ito, H.; Inoue, H.; Kimura, S.; Ohmori, T.; Ishikawa, F.; Gohda, K.; Sato, J. Prognostic impact of the number of viable circulating cells with high telomerase activity in gastric cancer patients: A prospective study. Int. J. Oncol. 2014, 45, 227–234. [Google Scholar] [CrossRef]
- Togo, S.; Katagiri, N.; Namba, Y.; Tulafu, M.; Nagahama, K.; Kadoya, K.; Takamochi, K.; Oh, S.; Suzuki, K.; Sakurai, F.; et al. Sensitive detection of viable circulating tumor cells using a novel conditionally telomerase-selective replicating adenovirus in non-small cell lung cancer patients. Oncotarget 2017, 8, 34884–34895. [Google Scholar] [CrossRef]
- Kim, M.; Suh, D.H.; Choi, J.Y.; Bu, J.; Kang, Y.-T.; Kim, K.; No, J.H.; Kim, Y.B.; Cho, Y.-H. Post-debulking circulating tumor cell as a poor prognostic marker in advanced stage ovarian cancer: A prospective observational study. Medicine 2019, 98, e15354. [Google Scholar] [CrossRef]
- Kyo, S.; Takakura, M.; Taira, T.; Kanaya, T.; Itoh, H.; Yutsudo, M.; Ariga, H.; Inoue, M. Sp1 cooperates with c-Myc to activate transcription of the human telomerase reverse transcriptase gene (hTERT). Nucleic Acids Res. 2000, 28, 669–677. [Google Scholar] [CrossRef]
- Oh, S.; Song, Y.-H.; Yim, J.; Kim, T.K. Identification of Mad as a repressor of the human telomerase (hTERT) gene. Oncogene 2000, 19, 1485–1490. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Ishihara, K.; Ichimura, T.; Fujita, N.; Hino, S.; Tomita, S.; Watanabe, S.; Saitoh, N.; Ito, T.; Nakao, M. MCAF1/AM is involved in Sp1-mediated maintenance of cancer-associated telomerase activity. J. Biol. Chem. 2009, 284, 5165–5174. [Google Scholar] [CrossRef] [PubMed]
- Won, J.; Yim, J.; Kim, T.K. Sp1 and Sp3 recruit histone deacetylase to repress transcription of human telomerase reverse transcriptase (hTERT) promoter in normal human somatic cells. J. Biol. Chem. 2002, 277, 38230–38238. [Google Scholar] [CrossRef] [PubMed]
- Maida, Y.; Kyo, S.; Kanaya, T.; Wang, Z.; Yatabe, N.; Tanaka, M.; Nakamura, M.; Ohmichi, M.; Gotoh, N.; Murakami, S.; et al. Direct activation of telomerase by EGF through Ets-mediated transactivation of TERT via MAP kinase signaling pathway. Oncogene 2002, 21, 4071–4079. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.-G.; Jayachandran, G.; Wu, G.; Xu, K.; Roth, J.A.; Ji, L. Tumor-specific activation of human telomerase reverses transcriptase promoter activity by activating enhancer-binding protein-2beta in human lung cancer cells. J. Biol. Chem. 2007, 282, 26460–26470. [Google Scholar] [CrossRef] [PubMed]
- Crowe, D.L.; Nguyen, D.C.; Tsang, K.J.; Kyo, S. E2F-1 represses transcription of the human telomerase reverse transcriptase gene. Nucleic Acids Res. 2001, 29, 2789–2794. [Google Scholar] [CrossRef]
- Sinha-Datta, U.; Horikawa, I.; Michishita, E.; Datta, A.; Sigler-Nicot, J.C.; Brown, M.; Kazanji, M.; Barrett, J.C.; Nicot, C. Transcriptional activation of hTERT through the NF-κB pathway in HTLV-I–transformed cells. Blood 2004, 104, 2523–2531. [Google Scholar] [CrossRef]
- Papanikolaou, V.; Athanassiou, E.; Dubos, S.; Dimou, I.; Papathanasiou, I.; Kitsiou-Tzeli, S.; Kappas, C.; Tsezou, A. hTERT regulation by NF-κB and c-myc in irradiated HER2-positive breast cancer cells. Int. J. Radiat. Biol. 2011, 87, 609–621. [Google Scholar] [CrossRef]
- Sitaram, R.T.; Degerman, S.; Ljungberg, B.; Andersson, E.; Oji, Y.; Sugiyama, H.; Roos, G.; Li, A. Wilms’ tumour 1 can suppress hTERT gene expression and telomerase activity in clear cell renal cell carcinoma via multiple pathways. Br. J. Cancer 2010, 103, 1255–1262. [Google Scholar] [CrossRef]
- Takakura, M.; Kyo, S.; Sowa, Y.; Wang, Z.; Yatabe, N.; Maida, Y.; Tanaka, M.; Inoue, M. Telomerase activation by histone deacetylase inhibitor in normal cells. Nucleic Acids Res. 2001, 29, 3006–3011. [Google Scholar] [CrossRef]
- Bischoff, J.R.; Kirn, D.H.; Williams, A.; Heise, C.; Horn, S.; Muna, M.; Ng, L.; Nye, J.A.; Sampson-Johannes, A.; Fattaey, A.; et al. An Adenovirus Mutant That Replicates Selectively in p53- Deficient Human Tumor Cells. Science 1996, 274, 373–376. [Google Scholar] [CrossRef]
- Rodriguez, R.; Schuur, E.R.; Lim, H.Y.; Henderson, G.A.; Simons, J.W.; Henderson, D.R. Prostate attenuated replication competent adenovirus (ARCA) CN706: A selective cytotoxic for prostate-specific antigen-positive prostate cancer cells. Cancer Res. 1997, 57, 2559–2563. [Google Scholar]
- Kurihara, T.; Brough, D.E.; Kovesdi, I.; Kufe, D.W. Selectivity of a replication-competent adenovirus for human breast carcinoma cells expressing the MUC1 antigen. J. Clin. Investig. 2000, 106, 763–771. [Google Scholar] [CrossRef][Green Version]
- Matsubara, S.; Wada, Y.; Gardner, T.A.; Egawa, M.; Park, M.S.; Hsieh, C.L.; Zhau, H.E.; Kao, C.; Kamidono, S.; Gillenwater, J.Y.; et al. A conditional replication-competent adenoviral vector, Ad-OC-E1a, to cotarget prostate cancer and bone stroma in an experimental model of androgen-independent prostate cancer bone metastasis. Cancer Res. 2001, 61, 6012–6019. [Google Scholar]
- Peng, X.Y.; Won, J.H.; Rutherford, T.; Fujii, T.; Zelterman, D.; Pizzorno, G.; Sapi, E.; Leavitt, J.; Kacinski, B.; Crystal, R.; et al. The use of the L-plastin promoter for adenoviral-mediated, tumor-specific gene expression in ovarian and bladder cancer cell lines. Cancer Res. 2001, 61, 4405–4413. [Google Scholar]
- Adachi, Y.; Reynolds, P.N.; Yamamoto, M.; Wang, M.; Takayama, K.; Matsubara, S.; Muramatsu, T.; Curiel, D.T. A midkine promoter-based conditionally replicative adenovirus for treatment of pediatric solid tumors and bone marrow tumor purging. Cancer Res. 2001, 61, 7882–7888. [Google Scholar]
- Tsukuda, K.; Wiewrodt, R.; Molnar-Kimber, K.; Jovanovic, V.P.; Amin, K.M. An E2F-responsive replication-selective adenovirus targeted to the defective cell cycle in cancer cells: Potent antitumoral efficacy but no toxicity to normal cell. Cancer Res. 2002, 62, 3438–3447. [Google Scholar]
- Kim, E.; Kim, J.-H.; Shin, H.-Y.; Lee, H.; Yang, J.M.; Kim, J.; Sohn, J.-H.; Kim, H.; Yun, C.-O. Ad-mTERT-delta19, a conditional replication-competent adenovirus driven by the human telomerase promoter, selectively replicates in and elicits cytopathic effect in a cancer cell-specific manner. Hum. Gene Ther. 2003, 14, 1415–1428. [Google Scholar] [CrossRef]
- Lanson, N.A.; Friedlander, P.L.; Schwarzenberger, P.; Kolls, J.K.; Wang, G. Replication of an adenoviral vector controlled by the human telomerase reverse transcriptase promoter causes tumor-selective tumor lysis. Cancer Res. 2003, 63, 7936–7941. [Google Scholar]
- Wirth, T.; Zender, L.; Schulte, B.; Mundt, B.; Plentz, R.; Rudolph, K.L.; Manns, M.; Kubicka, S.; Kühnel, F. A telomerase-dependent conditionally replicating adenovirus for selective treatment of cancer. Cancer Res. 2003, 63, 3181–3188. [Google Scholar]
- Irving, J.; Wang, Z.; Powell, S.; O’Sullivan, C.; Mok, M.; Murphy, B.; Cardoza, L.; Lebkowski, J.S.; Majumdar, A.S. Conditionally replicative adenovirus driven by the human telomerase promoter provides broad-spectrum antitumor activity without liver toxicity. Cancer Gene Ther. 2004, 11, 174–185. [Google Scholar] [CrossRef][Green Version]
- Kawashima, T.; Kagawa, S.; Kobayashi, N.; Shirakiya, Y.; Umeoka, T.; Teraishi, F.; Taki, M.; Kyo, S.; Tanaka, N.; Fujiwara, T. Telomerase-specific replication-selective virotherapy for human cancer. Clin. Cancer Res. 2004, 10, 285–292. [Google Scholar] [CrossRef]
- Fujiwara, T. Multidisciplinary oncolytic virotherapy for gastrointestinal cancer. Ann. Gastroenterol. Surg. 2019, 3, 396–404. [Google Scholar] [CrossRef]
- Kishimoto, H.; Kojima, T.; Watanabe, Y.; Kagawa, S.; Fujiwara, T.; Uno, F.; Teraishi, F.; Kyo, S.; Mizuguchi, H.; Hashimoto, Y.; et al. In vivo imaging of lymph node metastasis with telomerase-specific replication-selective adenovirus. Nat. Med. 2006, 12, 1213–1219. [Google Scholar] [CrossRef]
- Kojima, T.; Hashimoto, Y.; Watanabe, Y.; Kagawa, S.; Uno, F.; Kuroda, S.; Tazawa, H.; Kyo, S.; Mizuguchi, H.; Urata, Y.; et al. A simple biological imaging system for detecting viable human circulating tumor cells. J. Clin. Investig. 2009, 119, 3172–3181. [Google Scholar] [CrossRef]
- Kim, S.J.; Masago, A.; Tamaki, Y.; Akazawa, K.; Tsukamoto, F.; Sato, J.; Ozawa, T.; Tsujino, Y.; Noguchi, S. A novel approach using telomerase-specific replication-selective adenovirus for detection of circulating tumor cells in breast cancer patients. Breast Cancer Res. Treat. 2011, 128, 765–773. [Google Scholar] [CrossRef]
- Ju, M.; Kao, G.D.; Steinmetz, D.; Chandrasekaran, S.; Keefe, S.M.; Guzzo, T.J.; Christodouleas, J.P.; Hahn, S.M.; Dorsey, J.F. Application of a telomerase-based circulating tumor cell (CTC) assay in bladder cancer patients receiving postoperative radiation therapy: A case study. Cancer Biol. Ther. 2014, 15, 683–687. [Google Scholar] [CrossRef]
- Takakura, M.; Kyo, S.; Nakamura, M.; Maida, Y.; Mizumoto, Y.; Bono, Y.; Zhang, X.; Hashimoto, Y.; Urata, Y.; Fujiwara, T.; et al. Circulating tumour cells detected by a novel adenovirus-mediated system may be a potent therapeutic marker in gynaecological cancers. Br. J. Cancer 2012, 107, 448–454. [Google Scholar] [CrossRef]
- Macarthur, K.M.; Kao, G.D.; Chandrasekaran, S.; Alonso-Basanta, M.; Chapman, C.; Lustig, R.A.; Wileyto, E.P.; Hahn, S.M.; Dorsey, J.F. Detection of brain tumor cells in the peripheral blood by a telomerase promoter-based assay. Cancer Res. 2014, 74, 2152–2159. [Google Scholar] [CrossRef]
- Xu, M.J.; Cooke, M.; Steinmetz, D.; Karakousis, G.; Saxena, D.; Bartlett, E.; Xu, X.; Hahn, S.M.; Dorsey, J.F.; Kao, G.D. A novel approach for the detection and genetic analysis of live melanoma circulating tumor cells. PLoS ONE 2015, 10, e0123376. [Google Scholar] [CrossRef]
- Ito, H.; Inoue, H.; Sando, N.; Kimura, S.; Gohda, K.; Sato, J.; Murakami, K.; Ito, S.; Odaka, N.; Satodate, H.; et al. Prognostic impact of detecting viable circulating tumour cells in gastric cancer patients using a telomerase-specific viral agent: A prospective study. BMC Cancer 2012, 12, 346. [Google Scholar] [CrossRef]
- Yabusaki, M.; Sato, J.; Kohyama, A.; Kojima, T.; Nobuoka, D.; Yoshikawa, T.; Sawada, Y.; Murakami, K.; Gohda, K.; Okegawa, T.; et al. Detection and preliminary evaluation of circulating tumor cells in the peripheral blood of patients with eight types of cancer using a telomerase-specific adenovirus. Oncol. Rep. 2014, 32, 1772–1778. [Google Scholar] [CrossRef][Green Version]
- Vincent, T.; Neve, E.P.A.; Johnson, J.R.; Kukalev, A.; Rojo, F.; Albanell, J.; Pietras, K.; Virtanen, I.; Philipson, L.; Leopold, P.L.; et al. A SNAIL1-SMAD3/4 transcriptional repressor complex promotes TGF-beta mediated epithelial-mesenchymal transition. Nat. Cell Biol. 2009, 11, 943–950. [Google Scholar] [CrossRef]
- Matsumura-Arioka, Y.; Ohtani, K.; Hara, T.; Iwanaga, R.; Nakamura, M. Identification of two distinct elements mediating activation of telomerase (hTERT) gene expression in association with cell growth in human T cells. Int. Immunol. 2005, 17, 207–215. [Google Scholar] [CrossRef]
- Ge, Z.; Liu, C.; Björkholm, M.; Gruber, A.; Xu, D. Mitogen-activated protein kinase cascade-mediated histone H3 phosphorylation is critical for telomerase reverse transcriptase expression/telomerase activation induced by proliferation. Mol. Cell Biol. 2006, 26, 230–237. [Google Scholar] [CrossRef]
- Sakurai, F.; Narii, N.; Tomita, K.; Togo, S.; Takahashi, K.; Machitani, M.; Tachibana, M.; Ouchi, M.; Katagiri, N.; Urata, Y.; et al. Efficient detection of human circulating tumor cells without significant production of false-positive cells by a novel conditionally replicating adenovirus. Mol. Ther. Methods Clin. Dev. 2016, 3, 16001. [Google Scholar] [CrossRef][Green Version]
- Gaggar, A.; Shayakhmetov, D.M.; Lieber, A. CD46 is a cellular receptor for group B adenoviruses. Nat. Med. 2003, 9, 1408–1412. [Google Scholar] [CrossRef]
- Takakura, M.; Matsumoto, T.; Nakamura, M.; Mizumoto, Y.; Myojyo, S.; Yamazaki, R.; Iwadare, J.; Bono, Y.; Orisaka, S.; Obata, T.; et al. Detection of circulating tumor cells in cervical cancer using a conditionally replicative adenovirus targeting telomerase-positive cells. Cancer Sci. 2018, 109, 231–240. [Google Scholar] [CrossRef]
- Ma, Y.; Hao, S.; Wang, S.; Zhao, Y.; Lim, B.; Lei, M.; Spector, D.J.; El-Deiry, W.S.; Zheng, S.-Y.; Zhu, J. A Combinatory Strategy for Detection of Live CTCs Using Microfiltration and a New Telomerase-Selective Adenovirus. Mol. Cancer Ther. 2015, 14, 835–843. [Google Scholar] [CrossRef]
- Zhang, W.; Bao, L.; Yang, S.; Qian, Z.; Dong, M.; Yin, L.; Zhao, Q.; Ge, K.; Deng, Z.; Zhang, J.; et al. Tumor-selective replication herpes simplex virus-based technology significantly improves clinical detection and prognostication of viable circulating tumor cells. Oncotarget 2016, 7, 39768–39783. [Google Scholar] [CrossRef]
- Liu, Z.; Li, Q.; Li, K.; Chen, L.; Li, W.; Hou, M.; Liu, T.; Yang, J.; Lindvall, C.; Björkholm, M.; et al. Telomerase reverse transcriptase promotes epithelial-mesenchymal transition and stem cell-like traits in cancer cells. Oncogene 2013, 32, 4203–4213. [Google Scholar] [CrossRef]
- El-Badawy, A.; Ghoneim, N.I.; Nasr, M.A.; Elkhenany, H.; Ahmed, T.A.; Ahmed, S.M.; El-Badri, N. Telomerase reverse transcriptase coordinates with the epithelial-to-mesenchymal transition through a feedback loop to define properties of breast cancer stem cells. Biol. Open 2018, 7. [Google Scholar] [CrossRef]
- Huang, F.W.; Hodis, E.; Xu, M.J.; Kryukov, G.V.; Chin, L.; Garraway, L.A. Highly recurrent TERT promoter mutations in human melanoma. Science 2013, 339, 957–959. [Google Scholar] [CrossRef]
- Horn, S.; Figl, A.; Rachakonda, P.S.; Fischer, C.; Sucker, A.; Gast, A.; Kadel, S.; Moll, I.; Nagore, E.; Hemminki, K.; et al. TERT promoter mutations in familial and sporadic melanoma. Science 2013, 339, 959–961. [Google Scholar] [CrossRef]
- Barthel, F.P.; Wei, W.; Tang, M.; Martinez-Ledesma, E.; Hu, X.; Amin, S.B.; Akdemir, K.C.; Seth, S.; Song, X.; Wang, Q.; et al. Systematic analysis of telomere length and somatic alterations in 31 cancer types. Nat. Genet. 2017, 49, 349–357. [Google Scholar] [CrossRef]
- Zhang, A.; Zheng, C.; Lindvall, C.; Hou, M.; Ekedahl, J.; Lewensohn, R.; Yan, Z.; Yang, X.; Henriksson, M.; Blennow, E.; et al. Frequent amplification of the telomerase reverse transcriptase gene in human tumors. Cancer Res. 2000, 60, 6230–6235. [Google Scholar]



{kind=link}
{kind=link}
| Enrichment Method | Detection Method | Description | References | |
|---|---|---|---|---|
| CellSearch™ | Immunomagnetic beads (anti-EpCAM) | Cytokeratin (+)/DAPI (+)/ CD45 (−) | FDA approved Automated system | [20,21,22,23,26,30,31,32,33] |
| MetaCell® | Size based | RT-PCR | Commercially available kits Recoverable living cells | [19,38,39] |
| AdnaTest | Immunomagnetic beads | RT-PCR | Commercially available kits Specialized for target organ | [34,35,36,37] |
| Density-gradient centrifugation | Including OncoQuick® | RT-PCR, immunofluorescence | Recoverable living cells | [24,40] |
| Digital RT-PCR | Direct detection | RT-PCR (E6/E7) | No necessity of enrichment | [27] |
| WBC depletion | Immunomagnetic beads (anti-CD45) | Immunofluorescence (chromosome 8 aneuploidy) | [28,29] | |
| Immunomagnetic beads | Anti-EpCAM, -HER2, -MUC1 | RT-PCR | [41] | |
| Microfluidic device | Anti-EpCAM, -FAPα | Next-generation sequencing | Recoverable living cells | [42,43] |
| Microfluidic device | Size and/or deformability based | RT-PCR, immunofluorescence | Recoverable living cells | [25,44,45] |
| Telomerase-specific replication adenovirus | Expressing GFP | Immunofluorescence, PCR (E6/E7) | Infect only living cells Independent of surface markers | [46,47,48] |
| Author | Year | Methodology | Subjects | CTCs (+) Rate | CTCs Number | Remarks |
|---|---|---|---|---|---|---|
| Takakura et al. [46] | 2012 | OBP-401 + CD45 | 19 cervical 17 endometrial 5 ovarian 4 vulvar/vaginal | 40% | 2.9/5 mL | Change in CTCs was a surrogate marker of treatment response. |
| Yabusaki et al. [47] | 2014 | OBP-401 + size threshold | 8 cervical 4 endometrial | 42% | Not available | Cells >8.4 µm and GFP (+) were recognized as CTCs. |
| Takakura et al. [48] | 2018 | OBP-1101 + CD45 | 19 cervical | 26% | 2.5/7.5 mL | CTCs were negative for epithelial markers and were proved of tumor origin by HPV PCR. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takakura, M.; Takata, E.; Sasagawa, T. A Novel Liquid Biopsy Strategy to Detect Small Amounts of Cancer Cells Using Cancer-Specific Replication Adenoviruses. J. Clin. Med. 2020, 9, 4044. https://doi.org/10.3390/jcm9124044
Takakura M, Takata E, Sasagawa T. A Novel Liquid Biopsy Strategy to Detect Small Amounts of Cancer Cells Using Cancer-Specific Replication Adenoviruses. Journal of Clinical Medicine. 2020; 9(12):4044. https://doi.org/10.3390/jcm9124044
Chicago/Turabian StyleTakakura, Masahiro, Emi Takata, and Toshiyuki Sasagawa. 2020. "A Novel Liquid Biopsy Strategy to Detect Small Amounts of Cancer Cells Using Cancer-Specific Replication Adenoviruses" Journal of Clinical Medicine 9, no. 12: 4044. https://doi.org/10.3390/jcm9124044
APA StyleTakakura, M., Takata, E., & Sasagawa, T. (2020). A Novel Liquid Biopsy Strategy to Detect Small Amounts of Cancer Cells Using Cancer-Specific Replication Adenoviruses. Journal of Clinical Medicine, 9(12), 4044. https://doi.org/10.3390/jcm9124044