Anti-Inflammatory Action of Heterogeneous Nuclear Ribonucleoprotein A2/B1 in Patients with Autoimmune Endocrine Disorders

 , , , , , and
, , , , , and 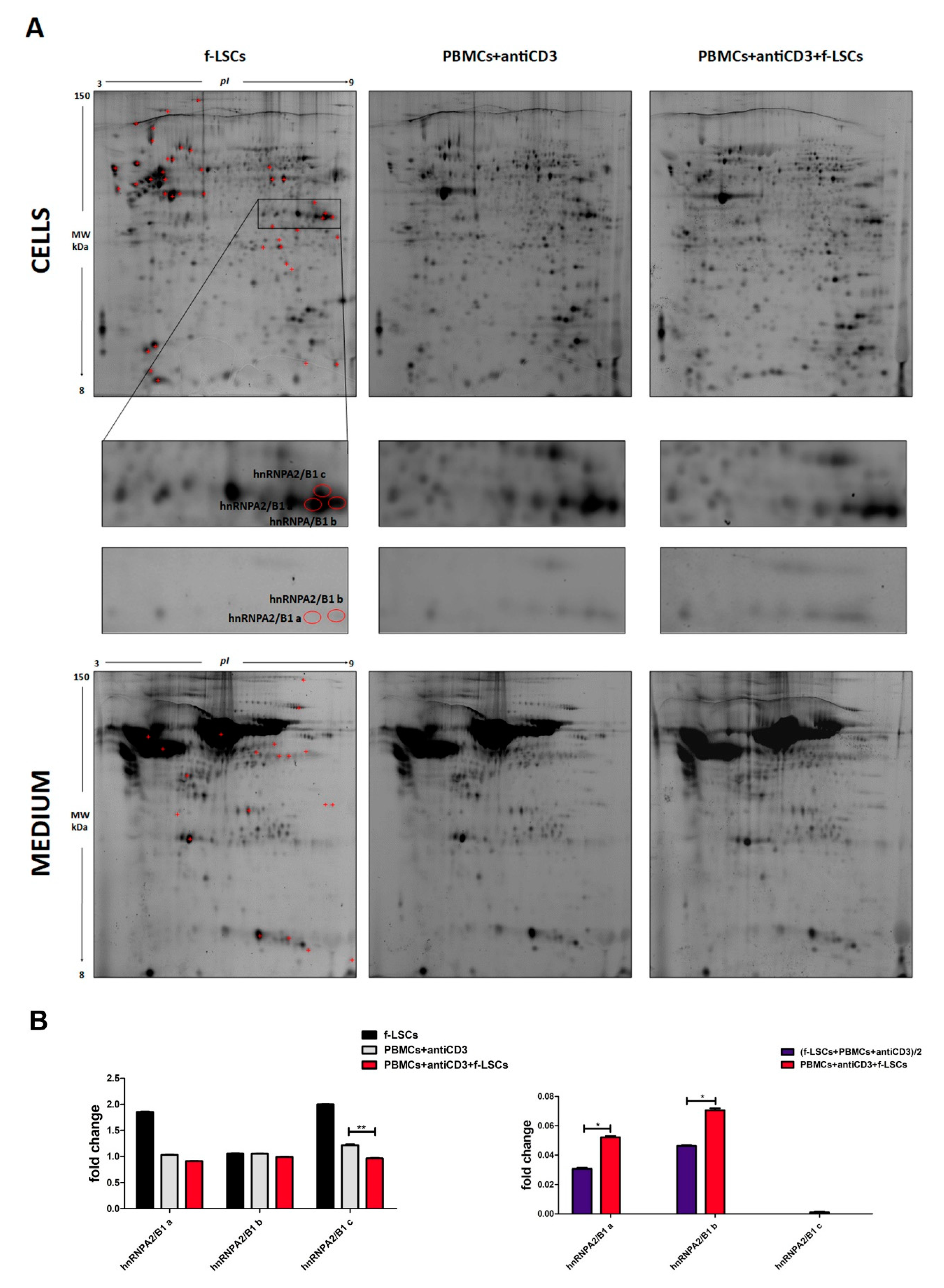{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Limbal Stem Cells
2.2. Patient Selection
2.3. Cell Culture and Cocultivation Experiments
2.4. Proteomic Analysis
2.5. 2D-DIGE Analysis
2.6. In-gel Digestion and MS Aanalysis of Tryptic Digests
2.7. Western Blot
2.8. Isolation of Total RNA and qRT-PCR
2.9. Flow Cytometry
2.10. Immunofluorescence
2.11. PBMCs Electroporation and hnRNPA2/B1 siRNA Knockdown
2.12. In vitro Pharmacological Treatments
2.13. Co-Immunoprecipitation Assay (Co-IP)
2.14. Statistical Analysis
2.15. FACS Analyses Were Performed in CellQuest
3. Results
3.1. Proteome and Secretome Analysis
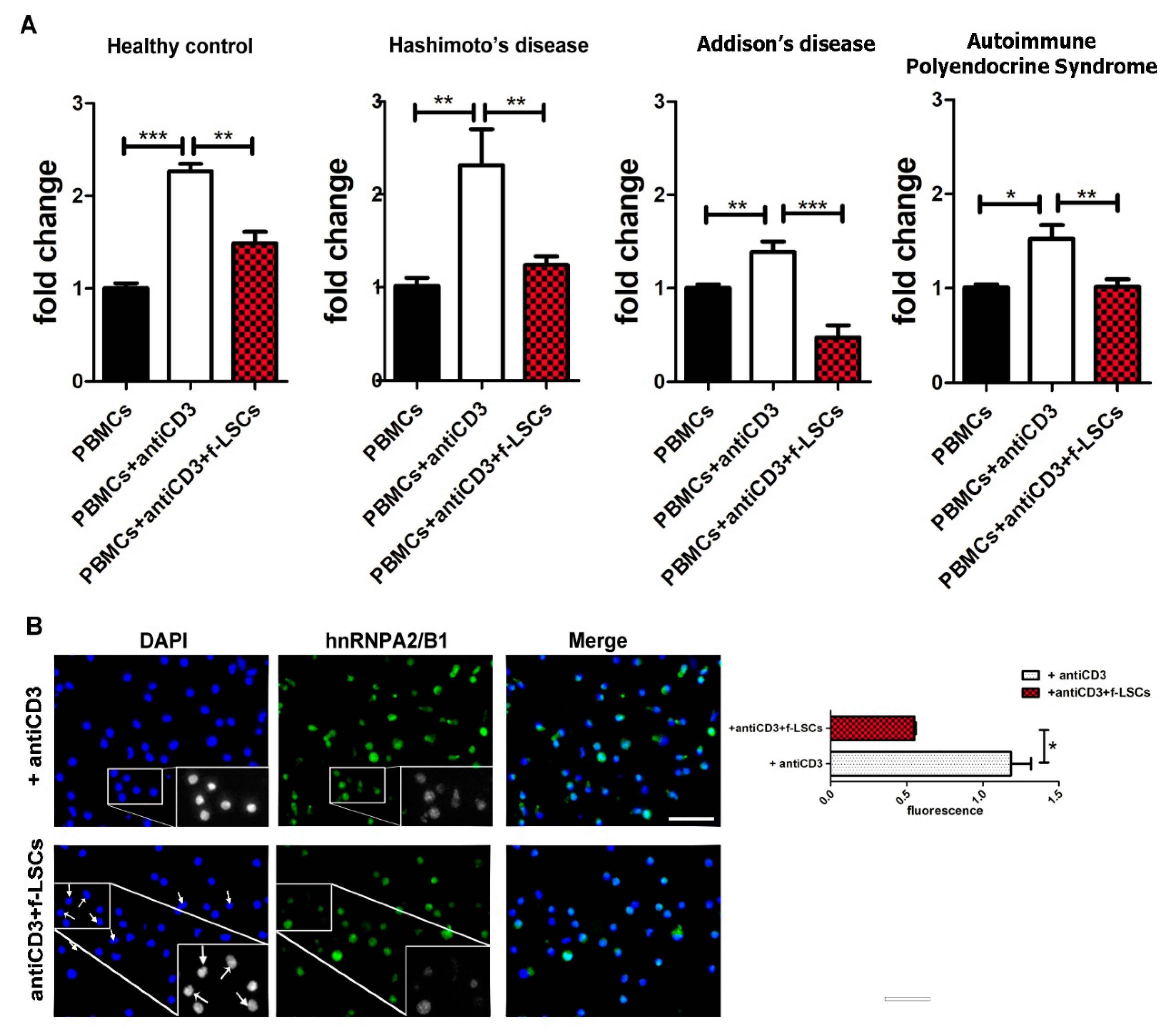
3.2. Downmodulation of hnRNPA2/B1 Expression in AED Patients
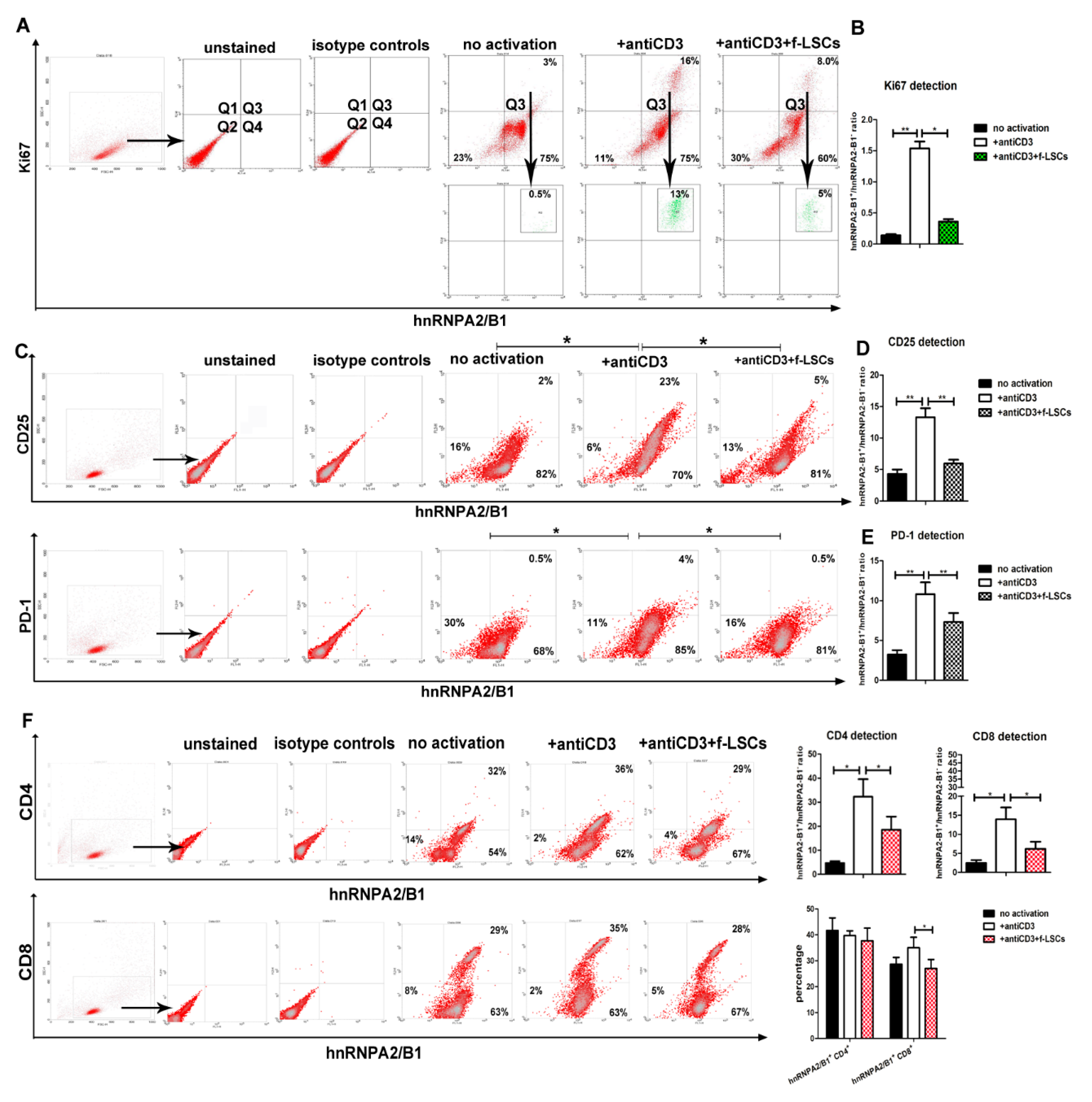
3.3. hnRNPA2/B1 Downmodulation Affects Proliferating and Activates Lympho-Monocytes of AED Patients
3.4. hnRNPA2/B1 Displays Immunoregulation Function Mainly on the CD8+ T Cell Subset
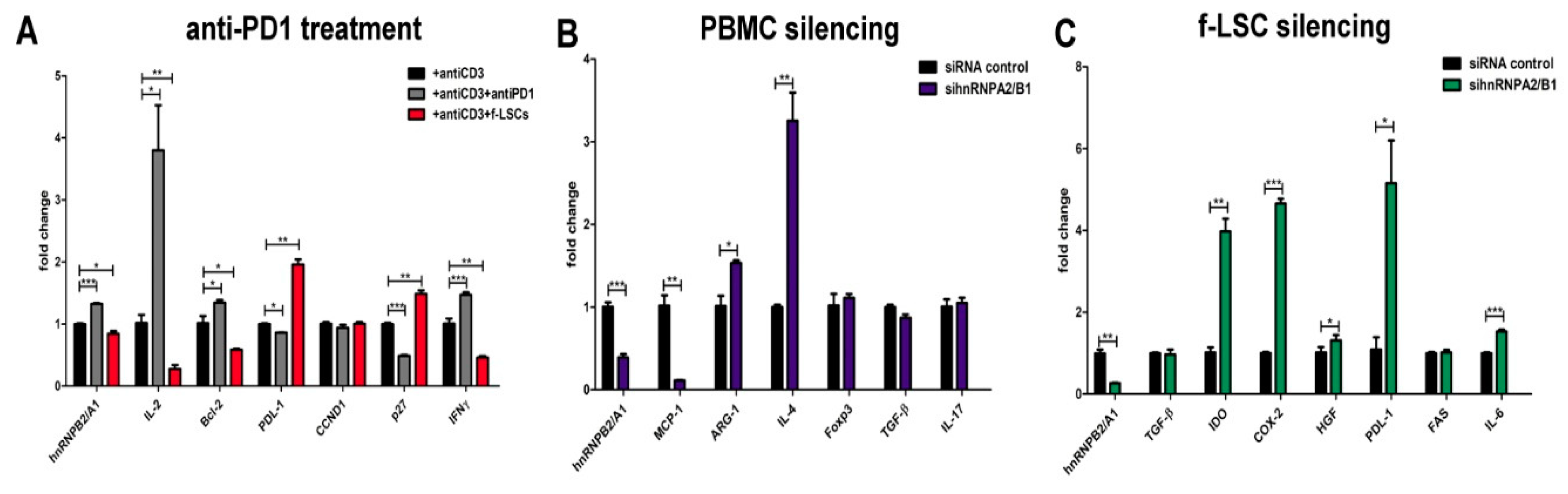
3.5. Anti-PD-1 Treatment Induces hnRNPA2/B1 Overexpression
3.6. hnRNPA2/B1 Silencing Induces M2/Th2 Polarization in Self-Reactive PBMCs of AED Patients
3.7. HnRNPA2/B1 Knockdown Improves Immunosuppressive Potential of f-LSCs
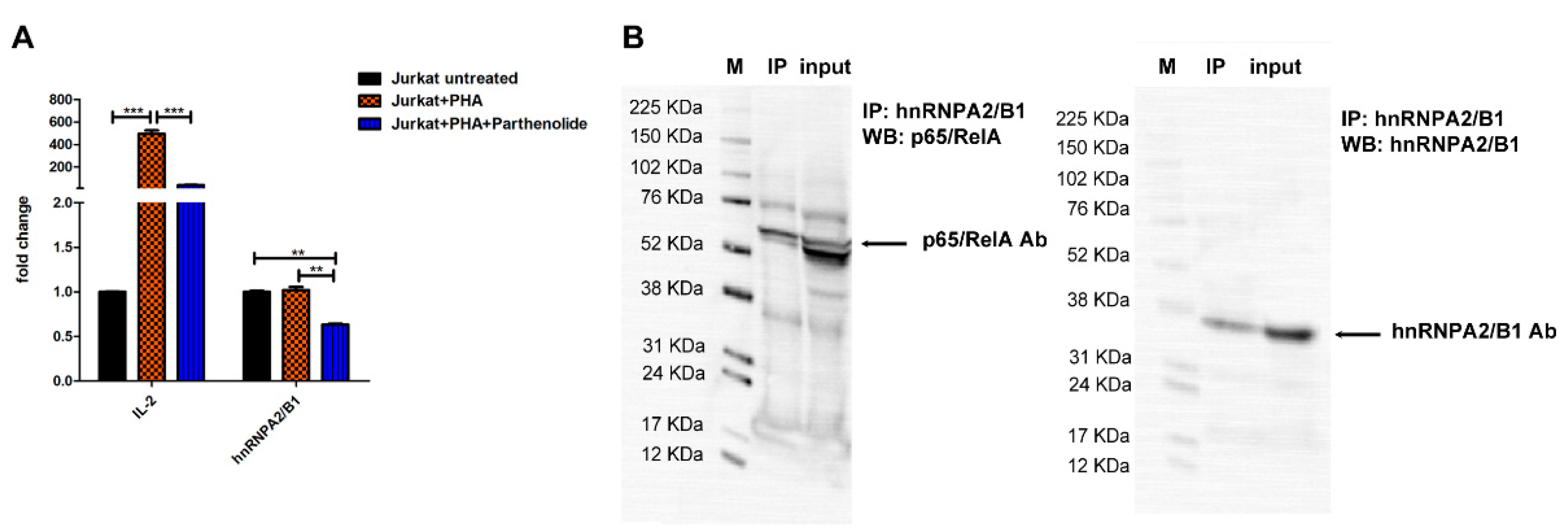
3.8. hnRNPA2/B1 mRNA Levels Are Influenced by NF-ĸB Transcription Fctor Activity in Jurkat T Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Eisenbarth, G.S.; Gottlieb, P.A. Autoimmune Polyendocrine Syndromes. N. Engl. J. Med. 2004, 350, 2068–2079. [Google Scholar] [CrossRef]
- Herman, S.; Fischer, A.; Presumey, J.; Hoffmann, M.; Koenders, M.I.; Escriou, V.; Apparailly, F.; Steiner, G. Inhibition of Inflammation and Bone Erosion by RNA Interference-Mediated Silencing of Heterogeneous Nuclear RNP A2/B1 in Two Experimental Models of Rheumatoid Arthritis. Arthritis Rheumatol. 2015, 67, 2536–2546. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.J.; Kim, N.C.; Wang, Y.-D.; Scarborough, E.A.; Moore, J.; Diaz, Z.; MacLea, K.S.; Freibaum, B.; Li, S.; Molliex, A.; et al. Mutations in prion-like domains in hnRNPA2B1 and hnRNPA1 cause multisystem proteinopathy and ALS. Nature 2013, 495, 467–473. [Google Scholar] [CrossRef]
- Yano, Y.; Wu, W.; Kamma, H.; Fujiwara, M.; Hara, H.; Yashiro, T.; Ueno, E.; Miwa, M.; Aiyoshi, Y. Expression of heterogeneous nuclear ribonucleoproteins A2 and b1 in the thyroid follicular cells. Endocr. Pathol. 2005, 16, 229–238. [Google Scholar] [CrossRef]
- Carpenter, B.; MacKay, C.; Alnabulsi, A.; MacKay, M.; Telfer, C.; Melvin, W.T.; Murray, G.I. The roles of heterogeneous nuclear ribonucleoproteins in tumour development and progression. Biochim. Biophys. Acta 2006, 1765, 85–100. [Google Scholar] [CrossRef]
- Shin, K.-H. Heterogeneous Nuclear Ribonucleoprotein G Shows Tumor Suppressive Effect against Oral Squamous Cell Carcinoma Cells. Clin. Cancer Res. 2006, 12, 3222–3228. [Google Scholar] [CrossRef] [Green Version]
- Tomasello, L.; Mauceri, R.; Coppola, A.; Pitrone, M.; Pizzo, G.; Campisi, G.; Pizzolanti, G.; Giordano, C. Mesenchymal stem cells derived from inflamed dental pulpal and gingival tissue: A potential application for bone formation. Stem Cell Res. Ther. 2017, 8, 179. [Google Scholar] [CrossRef] [Green Version]
- Parekkadan, B.; Tilles, A.W.; Yarmush, M.L. Bone Marrow-Derived Mesenchymal Stem Cells Ameliorate Autoimmune Enteropathy Independently of Regulatory T Cells. Stem Cells 2008, 26, 1913–1919. [Google Scholar] [CrossRef]
- Bernardo, M.E.; Fibbe, W.E. Mesenchymal Stromal Cells: Sensors and Switchers of Inflammation. Cell Stem Cell 2013, 13, 392–402. [Google Scholar] [CrossRef] [Green Version]
- Le Blanc, K.; Mougiakakos, D. Multipotent mesenchymal stromal cells and the innate immune system. Nat. Rev. Immunol. 2012, 12, 383–396. [Google Scholar] [CrossRef]
- Chen, Z.; de Paiva, C.S.; Luo, L.; Kretzer, F.L.; Pflugfelder, S.C.; Li, D.-Q. Characterization of Putative Stem Cell Phenotype in Human Limbal Epithelia: Characterization of Putative Stem Cell Phenotype in Human Limbal Epithelia. Stem Cells 2004, 22, 355–366. [Google Scholar] [CrossRef] [Green Version]
- Tomasello, L.; Musso, R.; Cillino, G.; Pitrone, M.; Pizzolanti, G.; Coppola, A.; Arancio, W.; Di Cara, G.; Pucci-Minafra, I.; Cillino, S.; et al. Donor age and long-term culture do not negatively influence the stem potential of limbal fibroblast-like stem cells. Stem Cell Res. Ther. 2016, 7, 83. [Google Scholar] [CrossRef] [Green Version]
- Coppola, A.; Tomasello, L.; Pitrone, M.; Cillino, S.; Richiusa, P.; Pizzolanti, G.; Giordano, C. Human limbal fibroblast-like stem cells induce immune-tolerance in autoreactive T lymphocytes from female patients with Hashimoto’s thyroiditis. Stem Cell Res. Ther. 2017, 8, 154. [Google Scholar] [CrossRef] [Green Version]
- Nagamine, K.; Peterson, P.; Scott, H.S.; Kudoh, J.; Minoshima, S.; Heino, M.; Krohn, K.J.E.; Lalioti, M.D.; Mullis, P.E.; Antonarakis, S.E.; et al. Positional cloning of the APECED gene. Nat. Genet. 1997, 17, 393–398. [Google Scholar] [CrossRef]
- Musso, R.; Di Cara, G.; Albanese, N.N.; Marabeti, M.R.; Cancemi, P.; Martini, D.; Orsini, E.; Giordano, C.; Pucci-Minafra, I. Differential proteomic and phenotypic behaviour of papillary and anaplastic thyroid cell lines. J. Proteom. 2013, 90, 115–125. [Google Scholar] [CrossRef]
- Buttacavoli, M.; Albanese, N.N.; Di Cara, G.; Alduina, R.; Faleri, C.; Gallo, M.; Pizzolanti, G.; Gallo, G.; Feo, S.; Baldi, F.; et al. Anticancer activity of biogenerated silver nanoparticles: An integrated proteomic investigation. Oncotarget 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- DI Cara, G.; Marengo, G.; Albanese, N.N.; Marabeti, M.R.; Musso, R.; Cancemi, P.; Pucci-Minafra, I. Proteomic profiling of Trastuzumab (Herceptin(R))-sensitive and -resistant SKBR-3 breast cancer cells. Anticancer Res. 2013, 33, 489–503. [Google Scholar]
- Coppola, A.; Tomasello, L.; Pizzolanti, G.; Pucci-Minafra, I.; Albanese, N.; Di Cara, G.; Cancemi, P.; Pitrone, M.; Bommarito, A.; Carissimi, E.; et al. In Vitro Phenotypic, Genomic and Proteomic Characterization of a Cytokine-Resistant Murine β-TC3 Cell Line. PLoS ONE 2012, 7, e32109. [Google Scholar] [CrossRef] [Green Version]
- Cancemi, P.; Di Cara, G.; Albanese, N.N.; Costantini, F.; Marabeti, M.R.; Musso, R.; Riili, I.; Lupo, C.; Roz, E.; Pucci-Minafra, I. Differential occurrence of S100A7 in breast cancer tissues: A proteomic-based investigation. Prot. Clin. Appl. 2012, 6, 364–373. [Google Scholar] [CrossRef] [Green Version]
- Pitrone, M.; Pizzolanti, G.; Tomasello, L.; Coppola, A.; Morini, L.; Pantuso, G.; Ficarella, R.; Guarnotta, V.; Perrini, S.; Giorgino, F.; et al. NANOG Plays a Hierarchical Role in the Transcription Network Regulating the Pluripotency and Plasticity of Adipose Tissue-Derived Stem Cells. Int. J. Mol. Sci. 2017, 18, 1107. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Burrows, G.G.; van’t Hof, W.; Reddy, A.P.; Wilmarth, P.A.; David, L.L.; Raber, A.; Bogaerts, A.; Timmerman, L.; Pinxteren, J.; Roobrouck, V.D.; et al. Solution-Phase Crosstalk and Regulatory Interactions Between Multipotent Adult Progenitor Cells and Peripheral Blood Mononuclear Cells: Anti-CD3/Anti-CD28-Activated PBMCs Plus MAPCs. Stem Cells Transl. Med. 2015, 4, 1436–1449. [Google Scholar] [CrossRef] [Green Version]
- Krampera, M. Mesenchymal stromal cell ‘licensing’: A multistep process. Leukemia 2011, 25, 1408–1414. [Google Scholar] [CrossRef] [Green Version]
- Uccelli, A.; Moretta, L.; Pistoia, V. Immunoregulatory function of mesenchymal stem cells. Eur. J. Immunol. 2006, 36, 2566–2573. [Google Scholar] [CrossRef]
- Ghannam, S.; Bouffi, C.; Djouad, F.; Jorgensen, C.; Noël, D. Immunosuppression by mesenchymal stem cells: Mechanisms and clinical applications. Stem Cell Res. Ther. 2010, 1, 2. [Google Scholar] [CrossRef] [Green Version]
- Anderson, M.S. Autoimmune endocrine disease. Curr. Opin. Immunol. 2002, 14, 760–764. [Google Scholar] [CrossRef]
- He, Y. Roles of heterogeneous nuclear ribonucleoproteins A and B in cell proliferation. J. Cell Sci. 2005, 118, 3173–3183. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.S.; Lee, H.M.; Jang, Y.-J.; Kim, C.-H.; Ryu, C.J. Heterogeneous Nuclear Ribonucleoprotein A2/B1 Regulates the Self-Renewal and Pluripotency of Human Embryonic Stem Cells Via the Control of the G1/S Transition: hnRNP A2/B1 Regulates G1/S Transition of hESCs. Stem Cells 2013, 31, 2647–2658. [Google Scholar] [CrossRef]
- Tauler, J.; Zudaire, E.; Liu, H.; Shih, J.; Mulshine, J.L. hnRNP A2/B1 Modulates Epithelial-Mesenchymal Transition in Lung Cancer Cell Lines. Cancer Res. 2010, 70, 7137–7147. [Google Scholar] [CrossRef] [Green Version]
- Glennie, S.; Soeiro, I.; Dyson, P.J.; Lam, E.W.-F.; Dazzi, F. Bone marrow mesenchymal stem cells induce division arrest anergy of activated T cells. Blood 2005, 105, 2821–2827. [Google Scholar] [CrossRef]
- Reddy, M.; Eirikis, E.; Davis, C.; Davis, H.M.; Prabhakar, U. Comparative analysis of lymphocyte activation marker expression and cytokine secretion profile in stimulated human peripheral blood mononuclear cell cultures: An in vitro model to monitor cellular immune function. J. Immunol. Methods 2004, 293, 127–142. [Google Scholar] [CrossRef]
- Jackson, A.L.; Matsumoto, H.; Janszen, M.; Maino, V.; Blidy, A.; Shye, S. Restricted expression of p55 interleukin 2 receptor (CD25) on normal T cells. Clin. Immunol. Immunopathol. 1990, 54, 126–133. [Google Scholar] [CrossRef]
- Keir, M.E.; Butte, M.J.; Freeman, G.J.; Sharpe, A.H. PD-1 and Its Ligands in Tolerance and Immunity. Annu. Rev. Immunol. 2008, 26, 677–704. [Google Scholar] [CrossRef] [Green Version]
- Agata, Y.; Kawasaki, A.; Nishimura, H.; Ishida, Y.; Tsubat, T.; Yagita, H.; Honjo, T. Expression of the PD-1 antigen on the surface of stimulated mouse T and B lymphocytes. Int. Immunol. 1996, 8, 765–772. [Google Scholar] [CrossRef] [Green Version]
- Freeman, G.J.; Long, A.J.; Iwai, Y.; Bourque, K.; Chernova, T.; Nishimura, H.; Fitz, L.J.; Malenkovich, N.; Okazaki, T.; Byrne, M.C.; et al. Engagement of the Pd-1 Immunoinhibitory Receptor by a Novel B7 Family Member Leads to Negative Regulation of Lymphocyte Activation. J. Exp. Med. 2000, 192, 1027–1034. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Thudium, K.B.; Han, M.; Wang, X.-T.; Huang, H.; Feingersh, D.; Garcia, C.; Wu, Y.; Kuhne, M.; Srinivasan, M.; et al. In Vitro Characterization of the Anti-PD-1 Antibody Nivolumab, BMS-936558, and In Vivo Toxicology in Non-Human Primates. Cancer Immunol. Res. 2014, 2, 846–856. [Google Scholar] [CrossRef] [Green Version]
- Akiyama, Y.; Nonomura, C.; Kondou, R.; Miyata, H.; Ashizawa, T.; Maeda, C.; Mitsuya, K.; Hayashi, N.; Nakasu, Y.; Yamaguchi, K. Immunological effects of the anti-programmed death-1 antibody on human peripheral blood mononuclear cells. Int. J. Oncol. 2016, 49, 1099–1107. [Google Scholar] [CrossRef] [Green Version]
- Gordon, S.; Martinez, F.O. Alternative Activation of Macrophages: Mechanism and Functions. Immunity 2010, 32, 593–604. [Google Scholar] [CrossRef] [Green Version]
- Mantovani, A.; Sica, A.; Locati, M. New vistas on macrophage differentiation and activation. Eur. J. Immunol. 2007, 37, 14–16. [Google Scholar] [CrossRef]
- Kidd, P. Th1/Th2 balance: The hypothesis, its limitations, and implications for health and disease. Altern. Med. Rev. 2003, 8, 223–246. [Google Scholar]
- Beyer, M.; Mallmann, M.R.; Xue, J.; Staratschek-Jox, A.; Vorholt, D.; Krebs, W.; Sommer, D.; Sander, J.; Mertens, C.; Nino-Castro, A.; et al. High-Resolution Transcriptome of Human Macrophages. PLoS ONE 2012, 7, e45466. [Google Scholar] [CrossRef]
- Bonfield, T.L.; Nolan (Koloze), M.T.; Lennon, D.P.; Caplan, A.I. Defining human mesenchymal stem cell efficacy in vivo. J. Inflamm. 2010, 7, 51. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Shi, S.; Liu, Y.; Uyanne, J.; Shi, Y.; Shi, S.; Le, A.D. Mesenchymal Stem Cells Derived from Human Gingiva Are Capable of Immunomodulatory Functions and Ameliorate Inflammation-Related Tissue Destruction in Experimental Colitis. J. Immunol. 2009, 183, 7787–7798. [Google Scholar] [CrossRef] [Green Version]
- Xiao, J.; Zhang, C.; Zhang, Y.; Zhang, X.; Zhao, J.; Liang, J.; Zhong, X.; Chen, Y. Transplantation of adipose-derived mesenchymal stem cells into a murine model of passive chronic immune thrombocytopenia: Adsc Transplantation and ITP. Transfusion 2012, 52, 2551–2558. [Google Scholar] [CrossRef]
- Hehner, S.P.; Hofmann, T.G.; Dröge, W.; Schmitz, M.L. The antiinflammatory sesquiterpene lactone parthenolide inhibits NF-kappa B by targeting the I kappa B kinase complex. J. Immunol. 1999, 163, 5617–5623. [Google Scholar]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coppola, A.; Cancemi, P.; Tomasello, L.; Guarnotta, V.; Pitrone, M.; Failla, V.; Cillino, S.; Feo, S.; Pizzolanti, G.; Giordano, C. Anti-Inflammatory Action of Heterogeneous Nuclear Ribonucleoprotein A2/B1 in Patients with Autoimmune Endocrine Disorders. J. Clin. Med. 2020, 9, 9. https://doi.org/10.3390/jcm9010009
Coppola A, Cancemi P, Tomasello L, Guarnotta V, Pitrone M, Failla V, Cillino S, Feo S, Pizzolanti G, Giordano C. Anti-Inflammatory Action of Heterogeneous Nuclear Ribonucleoprotein A2/B1 in Patients with Autoimmune Endocrine Disorders. Journal of Clinical Medicine. 2020; 9(1):9. https://doi.org/10.3390/jcm9010009
Chicago/Turabian StyleCoppola, Antonina, Patrizia Cancemi, Laura Tomasello, Valentina Guarnotta, Maria Pitrone, Valentina Failla, Salvatore Cillino, Salvatore Feo, Giuseppe Pizzolanti, and Carla Giordano. 2020. "Anti-Inflammatory Action of Heterogeneous Nuclear Ribonucleoprotein A2/B1 in Patients with Autoimmune Endocrine Disorders" Journal of Clinical Medicine 9, no. 1: 9. https://doi.org/10.3390/jcm9010009
APA StyleCoppola, A., Cancemi, P., Tomasello, L., Guarnotta, V., Pitrone, M., Failla, V., Cillino, S., Feo, S., Pizzolanti, G., & Giordano, C. (2020). Anti-Inflammatory Action of Heterogeneous Nuclear Ribonucleoprotein A2/B1 in Patients with Autoimmune Endocrine Disorders. Journal of Clinical Medicine, 9(1), 9. https://doi.org/10.3390/jcm9010009