Altered Bioenergetics of Blood Cell Sub-Populations in Acute Pancreatitis Patients


Abstract
1. Introduction
2. Experimental Section
2.1. Blood Collection and Cell Isolation
2.2. Assessment of Monocyte, Lymphocyte and Neutrophil Bioenergetics
2.3. Statistics
3. Results
3.1. Characteristics of Patients and Healthy Controls Included in the Analysis
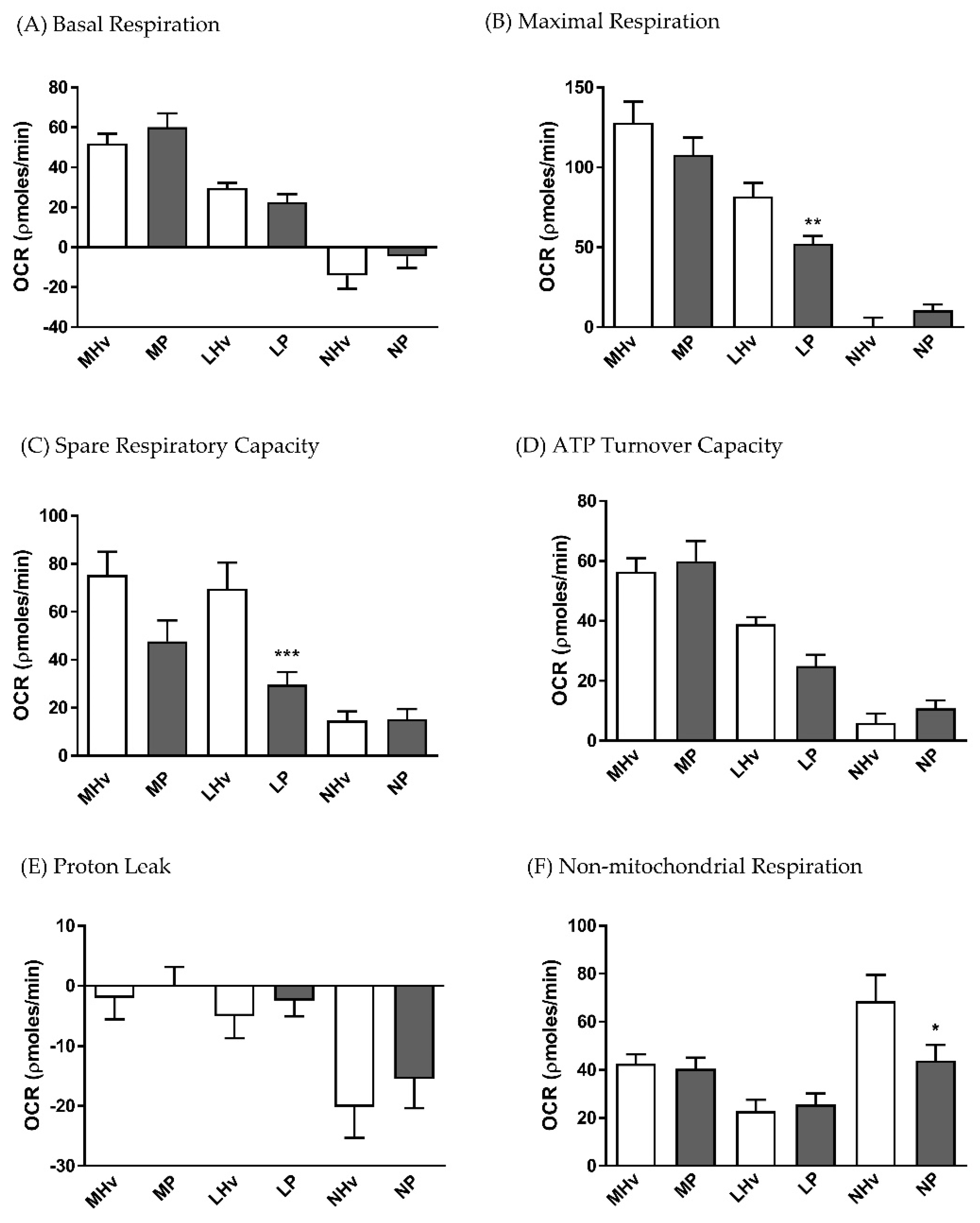
3.2. Bioenergetics Differences in OCR Between Healthy Volunteers and AP Patients
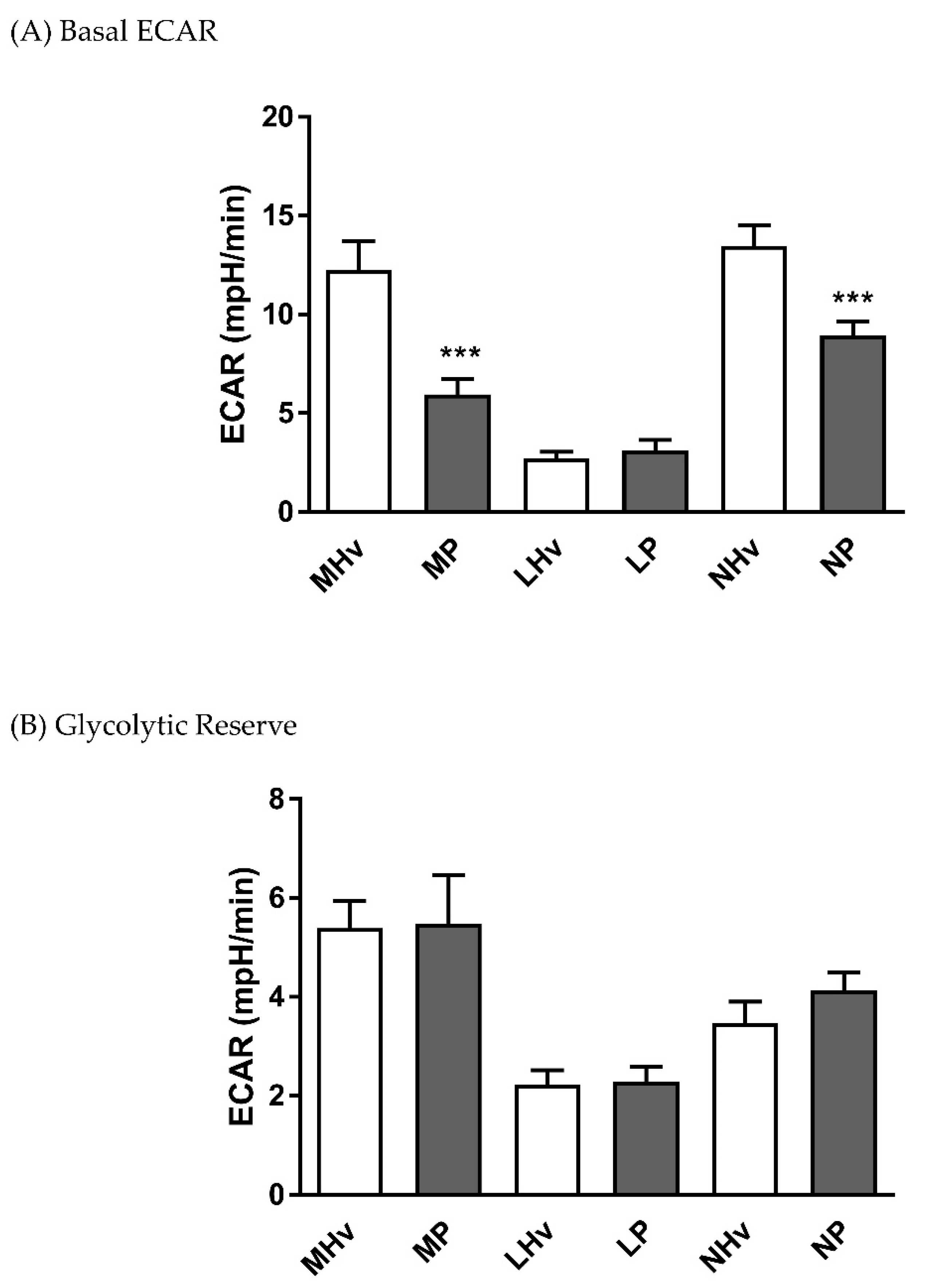
3.3. Analysis of Mitochondrial Bioenergetics Differences in ECAR Between Healthy Volunteers and AP Patients
3.4. Analysis of the Oxidative Burst in Healthy Volunteers and AP Patients
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Peery, A.F.; Crockett, S.D.; Murphy, C.C.; Lund, J.L.; Dellon, E.S.; Williams, J.L.; Jensen, E.T.; Shaheen, N.J.; Barritt, A.S.; Lieber, S.R.; et al. Burden and cost of gastrointestinal, liver, and pancreatic diseases in the united states: Update 2018. Gastroenterology 2019, 156, 254–272. [Google Scholar] [CrossRef] [PubMed]
- Yadav, D.; Lowenfels, A.B. The epidemiology of pancreatitis and pancreatic cancer. Gastroenterology 2013, 144, 1252–1261. [Google Scholar] [CrossRef] [PubMed]
- Kruger, B.; Albrecht, E.; Lerch, M.M. The role of intracellular calcium signaling in premature protease activation and the onset of pancreatitis. Am. J. Pathol. 2000, 157, 43–50. [Google Scholar] [CrossRef]
- Raraty, M.; Ward, J.; Erdemli, G.; Vaillant, C.; Neoptolemos, J.P.; Sutton, R.; Petersen, O.H. Calcium-dependent enzyme activation and vacuole formation in the apical granular region of pancreatic acinar cells. Proc. Natl. Acad. Sci. USA 2000, 97, 13126–13131. [Google Scholar] [CrossRef] [PubMed]
- Voronina, S.G.; Barrow, S.L.; Simpson, A.W.; Gerasimenko, O.V.; da Silva Xavier, G.; Rutter, G.A.; Petersen, O.H.; Tepikin, A.V. Dynamic changes in cytosolic and mitochondrial atp levels in pancreatic acinar cells. Gastroenterology 2010, 138, 1976–1987. [Google Scholar] [CrossRef] [PubMed]
- Criddle, D.N.; Murphy, J.; Fistetto, G.; Barrow, S.; Tepikin, A.V.; Neoptolemos, J.P.; Sutton, R.; Petersen, O.H. Fatty acid ethyl esters cause pancreatic calcium toxicity via inositol trisphosphate receptors and loss of atp synthesis. Gastroenterology 2006, 130, 781–793. [Google Scholar] [CrossRef] [PubMed]
- Booth, D.M.; Murphy, J.A.; Mukherjee, R.; Awais, M.; Neoptolemos, J.P.; Gerasimenko, O.V.; Tepikin, A.V.; Petersen, O.H.; Sutton, R.; Criddle, D.N. Reactive oxygen species induced by bile acid induce apoptosis and protect against necrosis in pancreatic acinar cells. Gastroenterology 2011, 140, 2116–2125. [Google Scholar] [CrossRef]
- Mukherjee, R.; Mareninova, O.A.; Odinokova, I.V.; Huang, W.; Murphy, J.; Chvanov, M.; Javed, M.A.; Wen, L.; Booth, D.M.; Cane, M.C.; et al. Mechanism of mitochondrial permeability transition pore induction and damage in the pancreas: Inhibition prevents acute pancreatitis by protecting production of atp. Gut 2016, 65, 1333–1346. [Google Scholar] [CrossRef]
- Wen, L.; Voronina, S.; Javed, M.A.; Awais, M.; Szatmary, P.; Latawiec, D.; Chvanov, M.; Collier, D.; Huang, W.; Barrett, J.; et al. Inhibitors of orai1 prevent cytosolic calcium-associated injury of human pancreatic acinar cells and acute pancreatitis in 3 mouse models. Gastroenterology 2015, 149, 481–492. [Google Scholar] [CrossRef]
- Criddle, D.N. Reactive oxygen species, ca(2+) stores and acute pancreatitis; a step closer to therapy? Cell Calcium 2016, 60, 180–189. [Google Scholar] [CrossRef]
- Xue, J.; Sharma, V.; Habtezion, A. Immune cells and immune-based therapy in pancreatitis. Immunol. Res. 2014, 58, 378–386. [Google Scholar] [CrossRef]
- Mayerle, J.; Sendler, M.; Hegyi, E.; Beyer, G.; Lerch, M.M.; Sahin-Toth, M. Genetics, cell biology, and pathophysiology of pancreatitis. Gastroenterology 2019, 156, 1951–1968. [Google Scholar] [CrossRef]
- Gukovskaya, A.S.; Vaquero, E.; Zaninovic, V.; Gorelick, F.S.; Lusis, A.J.; Brennan, M.L.; Holland, S.; Pandol, S.J. Neutrophils and nadph oxidase mediate intrapancreatic trypsin activation in murine experimental acute pancreatitis. Gastroenterology 2002, 122, 974–984. [Google Scholar] [CrossRef]
- Chakraborty, M.; Hickey, A.J.; Petrov, M.S.; Macdonald, J.R.; Thompson, N.; Newby, L.; Sim, D.; Windsor, J.A.; Phillips, A.R. Mitochondrial dysfunction in peripheral blood mononuclear cells in early experimental and clinical acute pancreatitis. Pancreatology 2016, 16, 739–747. [Google Scholar] [CrossRef]
- Brand, M.D.; Nicholls, D.G. Assessing mitochondrial dysfunction in cells. Biochem. J. 2011, 435, 297–312. [Google Scholar] [CrossRef]
- Kramer, P.A.; Chacko, B.K.; George, D.J.; Zhi, D.; Wei, C.C.; Dell’Italia, L.J.; Melby, S.J.; George, J.F.; Darley-Usmar, V.M. Decreased bioenergetic health index in monocytes isolated from the pericardial fluid and blood of post-operative cardiac surgery patients. Biosci. Rep. 2015, 35, e00237. [Google Scholar] [CrossRef]
- Tsai, K.; Wang, S.S.; Chen, T.S.; Kong, C.W.; Chang, F.Y.; Lee, S.D.; Lu, F.J. Oxidative stress: An important phenomenon with pathogenetic significance in the progression of acute pancreatitis. Gut 1998, 42, 850–855. [Google Scholar] [CrossRef]
- Armstrong, J.A.; Cash, N.J.; Ouyang, Y.; Morton, J.C.; Chvanov, M.; Latawiec, D.; Awais, M.; Tepikin, A.V.; Sutton, R.; Criddle, D.N. Oxidative stress alters mitochondrial bioenergetics and modifies pancreatic cell death independently of cyclophilin d, resulting in an apoptosis-to-necrosis shift. J. Biol. Chem. 2018, 293, 8032–8047. [Google Scholar] [CrossRef]
- Kramer, P.A.; Chacko, B.K.; Ravi, S.; Johnson, M.S.; Mitchell, T.; Darley-Usmar, V.M. Bioenergetics and the oxidative burst: Protocols for the isolation and evaluation of human leukocytes and platelets. J. Vis. Exp. 2014, 85, E51301. [Google Scholar] [CrossRef]
- Armstrong, J.A.; Cash, N.J.; Morton, J.C.; Tepikin, A.V.; Sutton, R.; Criddle, D.N. Mitochondrial targeting of antioxidants alters pancreatic acinar cell bioenergetics and determines cell fate. Int. J. Mol. Sci. 2019, 20, 1700. [Google Scholar] [CrossRef]
- Chacko, B.K.; Kramer, P.A.; Ravi, S.; Johnson, M.S.; Hardy, R.W.; Ballinger, S.W.; Darley-Usmar, V.M. Methods for defining distinct bioenergetic profiles in platelets, lymphocytes, monocytes, and neutrophils, and the oxidative burst from human blood. Lab. Investig. 2013, 93, 690–700. [Google Scholar] [CrossRef]
- Banks, P.A.; Bollen, T.L.; Dervenis, C.; Gooszen, H.G.; Johnson, C.D.; Sarr, M.G.; Tsiotos, G.G.; Vege, S.S.; Acute Pancreatitis Classification Working Group. Classification of acute pancreatitis—2012: Revision of the atlanta classification and definitions by international consensus. Gut 2013, 62, 102–111. [Google Scholar] [CrossRef]
- Oiva, J.; Mustonen, H.; Kylanpaa, M.L.; Kyhala, L.; Kuuliala, K.; Siitonen, S.; Kemppainen, E.; Puolakkainen, P.; Repo, H. Acute pancreatitis with organ dysfunction associates with abnormal blood lymphocyte signaling: Controlled laboratory study. Crit. Care 2010, 14, R207. [Google Scholar] [CrossRef]
- Sansbury, B.E.; Jones, S.P.; Riggs, D.W.; Darley-Usmar, V.M.; Hill, B.G. Bioenergetic function in cardiovascular cells: The importance of the reserve capacity and its biological regulation. Chem. Biol. Interact. 2011, 191, 288–295. [Google Scholar] [CrossRef]
- Yadava, N.; Nicholls, D.G. Spare respiratory capacity rather than oxidative stress regulates glutamate excitotoxicity after partial respiratory inhibition of mitochondrial complex i with rotenone. J. Neurosci. 2007, 27, 7310–7317. [Google Scholar] [CrossRef]
- Kramer, P.A.; Darley-Usmar, V.M. The emerging theme of redox bioenergetics in health and disease. Biomed. J. 2015, 38, 294–300. [Google Scholar]
- Kylanpaa, M.L.; Repo, H.; Puolakkainen, P.A. Inflammation and immunosuppression in severe acute pancreatitis. World J. Gastroenterol. 2010, 16, 2867–2872. [Google Scholar] [CrossRef]
- Cheng, S.C.; Scicluna, B.P.; Arts, R.J.; Gresnigt, M.S.; Lachmandas, E.; Giamarellos-Bourboulis, E.J.; Kox, M.; Manjeri, G.R.; Wagenaars, J.A.; Cremer, O.L.; et al. Broad defects in the energy metabolism of leukocytes underlie immunoparalysis in sepsis. Nat. Immunol. 2016, 17, 406–413. [Google Scholar] [CrossRef]
- Tsai, K.; Hsu, T.G.; Lu, F.J.; Hsu, C.F.; Liu, T.Y.; Kong, C.W. Age-related changes in the mitochondrial depolarization induced by oxidative injury in human peripheral blood leukocytes. Free Radic. Res. 2001, 35, 395–403. [Google Scholar] [CrossRef]
- Buck, M.D.; O’Sullivan, D.; Klein Geltink, R.I.; Curtis, J.D.; Chang, C.H.; Sanin, D.E.; Qiu, J.; Kretz, O.; Braas, D.; van der Windt, G.J.; et al. Mitochondrial dynamics controls t cell fate through metabolic programming. Cell 2016, 166, 63–76. [Google Scholar] [CrossRef]
- Weiss, S.L.; Selak, M.A.; Tuluc, F.; Perales Villarroel, J.; Nadkarni, V.M.; Deutschman, C.S.; Becker, L.B. Mitochondrial dysfunction in peripheral blood mononuclear cells in pediatric septic shock. Pediatr. Crit. Care Med. 2015, 16, e4–e12. [Google Scholar] [CrossRef]
- Karlsson, A.; Nixon, J.B.; McPhail, L.C. Phorbol myristate acetate induces neutrophil nadph-oxidase activity by two separate signal transduction pathways: Dependent or independent of phosphatidylinositol 3-kinase. J. Leukoc. Biol. 2000, 67, 396–404. [Google Scholar] [CrossRef]
- Sandoval, D.; Gukovskaya, A.; Reavey, P.; Gukovsky, S.; Sisk, A.; Braquet, P.; Pandol, S.J.; Poucell-Hatton, S. The role of neutrophils and platelet-activating factor in mediating experimental pancreatitis. Gastroenterology 1996, 111, 1081–1091. [Google Scholar] [CrossRef]
- Abdulla, A.; Awla, D.; Thorlacius, H.; Regner, S. Role of neutrophils in the activation of trypsinogen in severe acute pancreatitis. J. Leukoc. Biol. 2011, 90, 975–982. [Google Scholar] [CrossRef]
- Sauce, D.; Dong, Y.; Campillo-Gimenez, L.; Casulli, S.; Bayard, C.; Autran, B.; Boddaert, J.; Appay, V.; Elbim, C. Reduced oxidative burst by primed neutrophils in the elderly individuals is associated with increased levels of the cd16bright/cd62ldim immunosuppressive subset. J. Gerontol. A Biol. Sci. Med. Sci. 2017, 72, 163–172. [Google Scholar] [CrossRef]
- Bhatia, M.; Ramnath, R.D.; Chevali, L.; Guglielmotti, A. Treatment with bindarit, a blocker of mcp-1 synthesis, protects mice against acute pancreatitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2005, 288, G1259–G1265. [Google Scholar] [CrossRef]
- Sakai, Y.; Masamune, A.; Satoh, A.; Nishihira, J.; Yamagiwa, T.; Shimosegawa, T. Macrophage migration inhibitory factor is a critical mediator of severe acute pancreatitis. Gastroenterology 2003, 124, 725–736. [Google Scholar] [CrossRef]
- Yeligar, S.M.; Harris, F.L.; Hart, C.M.; Brown, L.A. Ethanol induces oxidative stress in alveolar macrophages via upregulation of nadph oxidases. J. Immunol. 2012, 188, 3648–3657. [Google Scholar] [CrossRef]
- Sendler, M.; Weiss, F.U.; Golchert, J.; Homuth, G.; van den Brandt, C.; Mahajan, U.M.; Partecke, L.I.; Doring, P.; Gukovsky, I.; Gukovskaya, A.S.; et al. Cathepsin b-mediated activation of trypsinogen in endocytosing macrophages increases severity of pancreatitis in mice. Gastroenterology 2018, 154, 704–718. [Google Scholar] [CrossRef]
- Schmid, D.; Burmester, G.R.; Tripmacher, R.; Kuhnke, A.; Buttgereit, F. Bioenergetics of human peripheral blood mononuclear cell metabolism in quiescent, activated, and glucocorticoid-treated states. Biosci. Rep. 2000, 20, 289–302. [Google Scholar] [CrossRef]
- Sun, N.; Youle, R.J.; Finkel, T. The mitochondrial basis of aging. Mol. Cell 2016, 61, 654–666. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient | Severity (1 = Mild, 2 = Moderate & 3 = Severe) | Aetiology (ERCP = Endoscopic Retrograde Cholangiopancreatography) | Sex (M = Male, F = Female) | Age | Amylase | Platelets (×10 > 9/L, N: 150–400) | White Blood Cells (×10 > 9/L, N: 3.5–11) | Neutrophils (×10 > 9/L, N: 2.0–7.5) | Lymphocytes (×10 > 9/L, N: 1.0–3.5) | Monocytes (×10 > 9/L, N: 0.2–0.8) | Eosinophils (×10 > 9/L, N: 0.0–0.4) | Basophils (×10 >9/L, N: 0.0–0.2) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AP779 | 1 | Biliary | M | 59 | 2634 | 228 | 10.8 | 8.3 | 1.5 | 0.8 | 0.1 | 0.1 |
| AP784 | 3 | Idiopathic | F | 77 | 1240 | 293 | 25.6 | 22.7 | 1.4 | 1.5 | 0 | 0.1 |
| AP785 | 1 | Biliary | M | 74 | 527 | 233 | 19.4 | 17.3 | 0.6 | 1.3 | 0.1 | 0 |
| AP788 | 1 | Biliary | F | 20 | 1692 | 197 | 12.3 | 8.7 | 2.2 | 1.3 | 0 | 0 |
| AP796 | 1 | Biliary | F | 64 | 1485 | 246 | 13.1 | 10.2 | 1.6 | 1.2 | 0 | 0 |
| AP797 | 1 | Biliary | M | 63 | 2168 | 245 | 11.3 | 10.2 | 0.4 | 0.6 | 0 | 0 |
| AP799 | 1 | Biliary | F | 28 | 1968 | 278 | 8.9 | 7 | 1.4 | 0.5 | 0 | 0.1 |
| AP805 | 2 | Biliary | F | 80 | 1577 | 303 | 10.3 | 9.1 | 0.8 | 0.3 | 0 | 0 |
| AP806 | 1 | Alcohol | M | 65 | 1265 | 188 | 10.7 | 8 | 1.3 | 1.2 | 0.1 | 0.1 |
| AP812 | 1 | Idiopathic | F | 21 | 492 | 194 | 12 | 6.7 | 4.1 | 1 | 0.2 | 0 |
| AP821 | 1 | Biliary | F | 33 | 2333 | 335 | 12.9 | 9.9 | 1.9 | 1 | 0.1 | 0.1 |
| AP828 | 1 | Biliary | F | 77 | 1792 | 237 | 23 | 21.4 | 1.3 | 0.2 | 0.1 | 0.1 |
| AP837 | 3 | Biliary | F | 91 | 1450 | 190 | 12.4 | 11.8 | 0.4 | 0.3 | 0 | 0 |
| AP839 | 1 | Biliary | F | 49 | 2293 | 264 | 9.5 | 6 | 2.5 | 0.9 | 0.1 | 0 |
| AP842 | 1 | ERCP | F | 57 | 2184 | 229 | 10.5 | 9.6 | 0.6 | 0.3 | 0 | 0 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morton, J.C.; Armstrong, J.A.; Sud, A.; Tepikin, A.V.; Sutton, R.; Criddle, D.N. Altered Bioenergetics of Blood Cell Sub-Populations in Acute Pancreatitis Patients. J. Clin. Med. 2019, 8, 2201. https://doi.org/10.3390/jcm8122201
Morton JC, Armstrong JA, Sud A, Tepikin AV, Sutton R, Criddle DN. Altered Bioenergetics of Blood Cell Sub-Populations in Acute Pancreatitis Patients. Journal of Clinical Medicine. 2019; 8(12):2201. https://doi.org/10.3390/jcm8122201
Chicago/Turabian StyleMorton, Jack C., Jane A. Armstrong, Ajay Sud, Alexei V. Tepikin, Robert Sutton, and David N. Criddle. 2019. "Altered Bioenergetics of Blood Cell Sub-Populations in Acute Pancreatitis Patients" Journal of Clinical Medicine 8, no. 12: 2201. https://doi.org/10.3390/jcm8122201
APA StyleMorton, J. C., Armstrong, J. A., Sud, A., Tepikin, A. V., Sutton, R., & Criddle, D. N. (2019). Altered Bioenergetics of Blood Cell Sub-Populations in Acute Pancreatitis Patients. Journal of Clinical Medicine, 8(12), 2201. https://doi.org/10.3390/jcm8122201