
A β-Thalassemia Cell Biobank: Updates, Further Validation in Genetic and Therapeutic Research and Opportunities During (and After) the COVID-19 Pandemic

 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Isolation and Culture of Peripheral Blood Cells
2.3. Freezing, Cryopreservation, and Thawing
2.4. Quality Control Procedures
2.5. Treatment of Long-Storage Biobanked Cells with HbF Inducers
2.6. Treatment of Cells with the β039 CRISPR-Cas9 System
3. Results
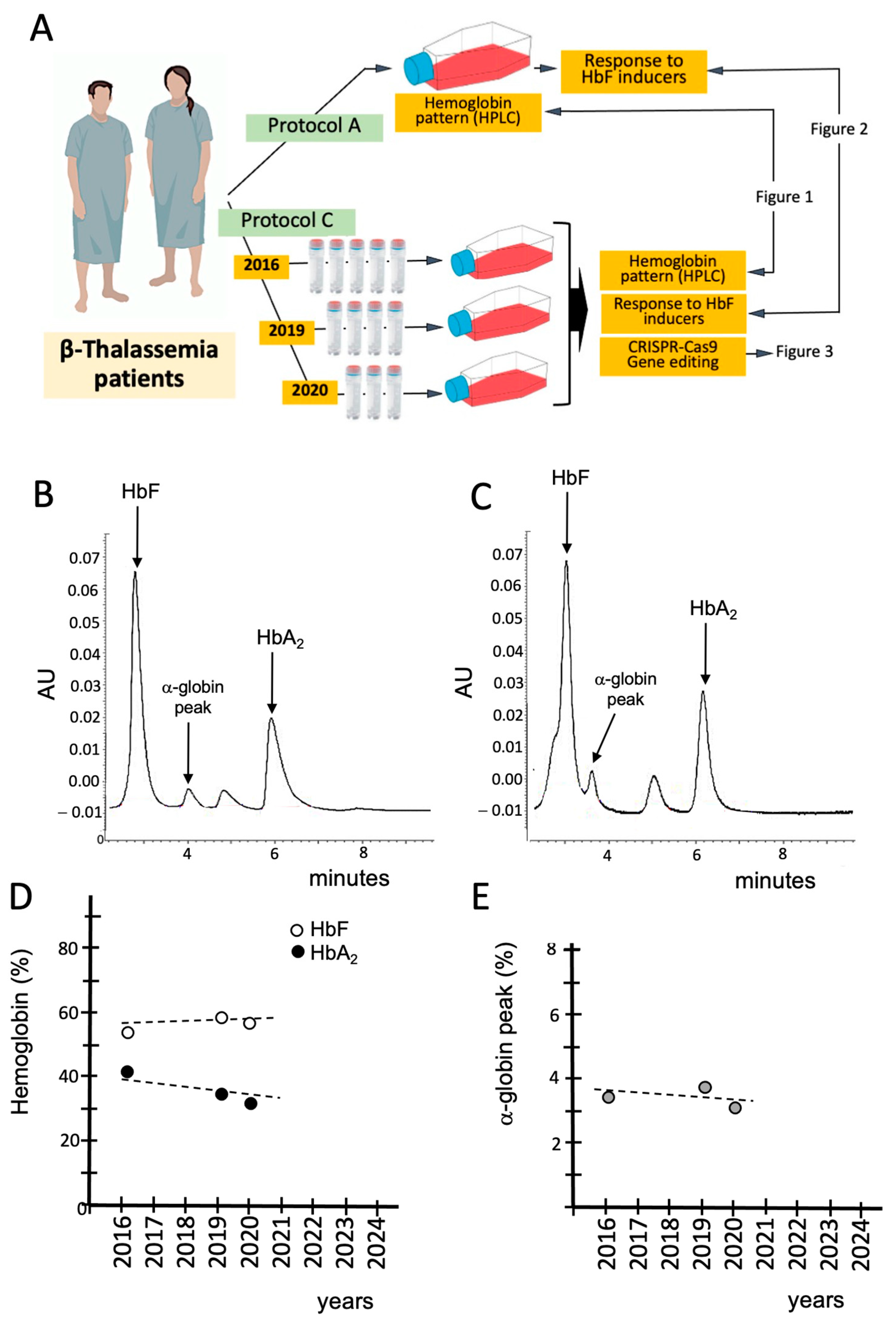
3.1. Characterization and Validation of the β-Thal Cell Biobank: Biobanked Samples of the Same β-Thalassemia Patient Maintain the Hemoglobin Pattern After Long-Time (6 Years) Storage
3.2. Validation of the β-Thal Cell Biobank: Use of Biobanked Samples in Pre-Clinical Studies
3.3. Validation of the β-Thal Cell Biobank: An Update on the Induction of Fetal Hemoglobin (HbF) Using Biobanked Samples
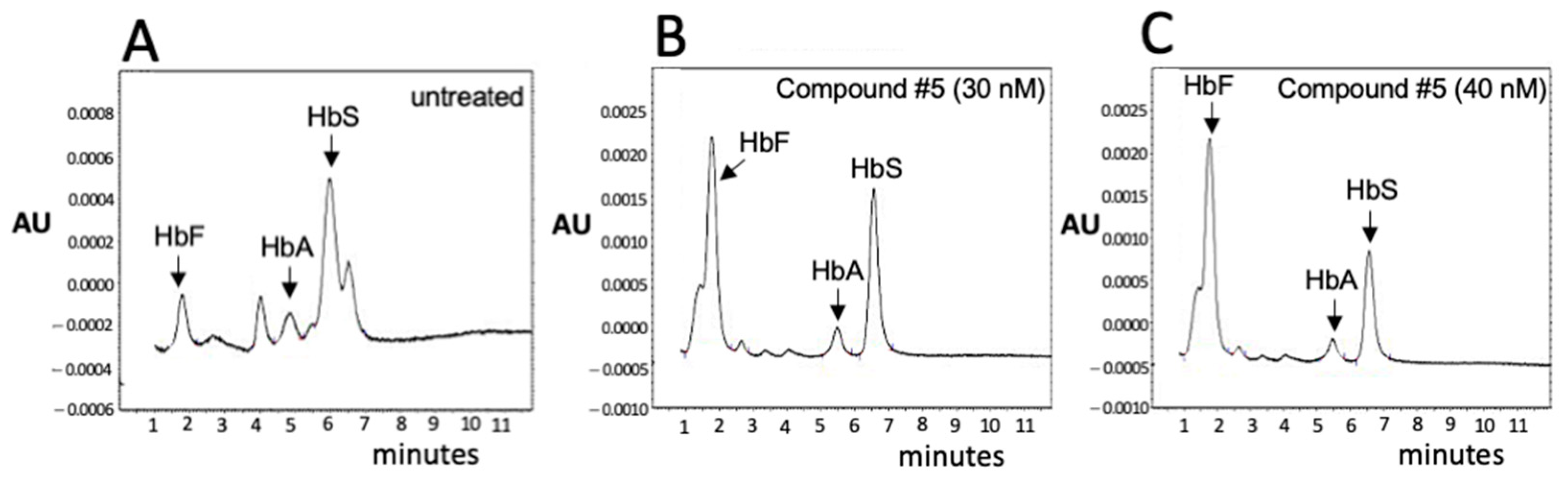
3.4. Validation of the β-Thal Cell Biobank: CRISPR-Cas9-Based Gene Editing Using Biobanked Samples
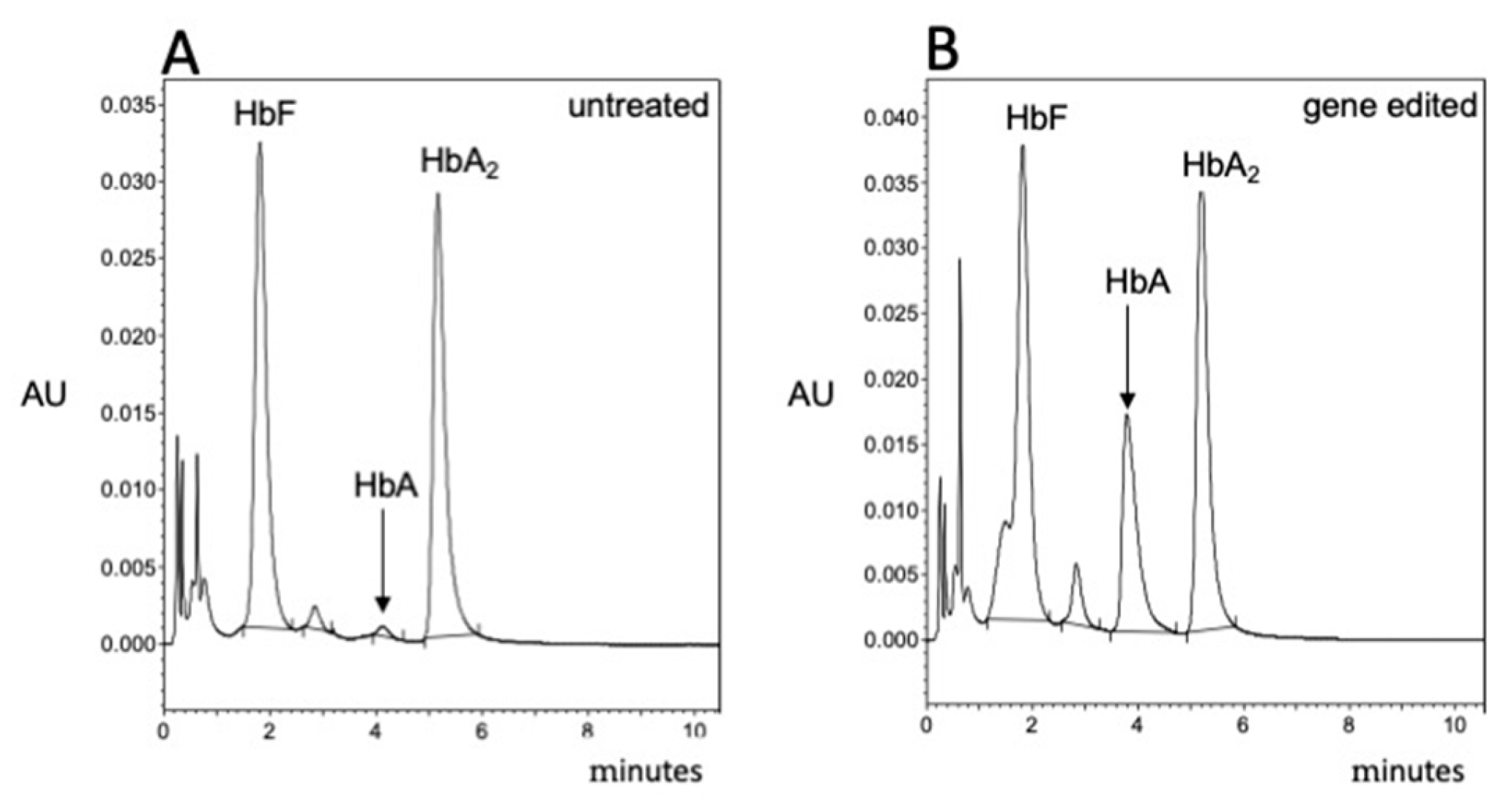
3.5. Updates on the Composition of the Cell β-Thal Cell Biobank
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Al Diffalha, S.; Sexton, K.C.; Watson, P.H.; Grizzle, W.E. The importance of human tissue bioresources in advancing biomedical research. Biopreservation Biobanking 2019, 17, 209–212. [Google Scholar] [CrossRef]
- Paskal, W.; Paskal, A.M.; Dębski, T.; Gryziak, M.; Jaworowski, J. Aspects of modern biobank activity–comprehensive review. Pathol. Oncol. Res. 2018, 24, 771–785. [Google Scholar] [CrossRef]
- Compton, C.; Kelly, A. Biospecimen banking in the post-genome era. Genom. Pers. Med. 2013, 1, 229–236. [Google Scholar]
- Annaratone, L.; De Palma, G.; Bonizzi, G.; Sapino, A.; Botti, G.; Berrino, E.; Mannelli, C.; Arcella, P.; Di Martino, S.; Steffan, A.; et al. Basic principles of biobanking: From biological samples to precision medicine for patients. Virchows Arch. 2021, 479, 233–246. [Google Scholar] [CrossRef] [PubMed]
- Tang, R.; Han, C.; Yin, R.; Zhu, P.; Zhu, L.; Lu, Y.; Zheng, C. Quality Control of DNA Extracted from All-Cell Pellets After Cryopreservation for More Than 10 Years. Biopreservation Biobanking 2022, 20, 211–216. [Google Scholar] [CrossRef]
- Juurikka, K.; Åström, P.; Pekkala, T.; Öhman, H.; Sorsa, T.; Tervahartiala, T.; Salo, T.; Lehenkari, P.; Nyberg, P. Insights into Preservation of Blood Biomarkers in Biobank Samples. Biopreservation Biobanking 2022, 20, 297–301. [Google Scholar] [CrossRef] [PubMed]
- O’Donoghue, S.; Dee, S.; Byrne, J.A.; Watson, P.H. How Many Health Research Biobanks Are There? Biopreservation Biobanking 2022, 20, 224–228. [Google Scholar] [CrossRef]
- Coppola, L.; Cianflone, A.; Grimaldi, A.M.; Incoronato, M.; Bevilacqua, P.; Messina, F.; Baselice, S.; Soricelli, A.; Mirabelli, P.; Salvatore, M. Biobanking in health care: Evolution and future directions. J. Transl. Med. 2019, 17, 172. [Google Scholar] [CrossRef] [PubMed]
- Rush, A.; Catchpoole, D.R.; Reaiche-Miller, G.; Gilbert, T.; Ng, W.; Watson, P.H.; Byrne, J.A. What Do Biomedical Researchers Want from Biobanks? Results of an Online Survey. Biopreservation Biobanking 2022, 20, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Samuel, G.; Lucassen, A. Access to Biobanks: Responsibilities Within a Research Ecosystem. Biopreservation Biobanking 2023, 21, 275–281. [Google Scholar] [CrossRef]
- Gille, F.; Vayena, E.; Blasimme, A. Future-proofing biobanks’ governance. Eur. J. Hum. Genet. 2020, 28, 989–996. [Google Scholar] [CrossRef] [PubMed]
- Roslan, F.F.; Yu, Y.; Ooi, G.C.; Then, K.L.; Then, K.Y.; Cheong, S.K.; Guo, Z.; Ab Patar, M.N.A.; Tan, J.J. From banked human cord blood to induced pluripotent stem cells: New opportunities and promise in induced pluripotent stem cell banking (Review). Int. J. Mol. Med. 2024, 54, 114. [Google Scholar] [CrossRef]
- Abraham, M.; Goel, S. Comprehensive characterisation and cryopreservation optimisation of buffalo (Bubalus bubalis) adipose tissue-derived mesenchymal stem cells. Cryobiology 2024, 115, 104896. [Google Scholar] [CrossRef] [PubMed]
- Monje, P.V. Human Schwann Cells in vitro II. Passaging, Purification, Banking, and Labeling of Established Cultures. Bio Protoc. 2023, 13, e4882. [Google Scholar] [CrossRef]
- D’Vaz, N.; Kidd, C.; Miller, S.; Amin, M.; Davis, J.A.; Talati, Z.; Silva, D.T.; Prescott, S.L. The ORIGINS Project Biobank: A Collaborative Bio Resource for Investigating the Developmental Origins of Health and Disease. Int. J. Environ. Res. Public Health 2023, 20, 6297. [Google Scholar] [CrossRef]
- Brophy, S.; Amet, R.; Foy-Stones, H.; Gardiner, N.; McElligott, A.M. Isolation and Cryopreservation of Mononuclear Cells from Peripheral Blood and Bone Marrow of Blood Cancer Patients. Methods Mol. Biol. 2023, 2645, 179–187. [Google Scholar]
- Degnin, M.; Agarwal, A.; Tarlock, K.; Meshinchi, S.; Druker, B.J.; Tognon, C.E. Novel Method Enabling the Use of Cryopreserved Primary Acute Myeloid Leukemia Cells in Functional Drug Screens. J. Pediatr. Hematol. Oncol. 2017, 39, e359–e366. [Google Scholar] [CrossRef] [PubMed]
- Harris, J.R.; Burton, P.; Knoppers, B.M.; Lindpaintner, K.; Bledsoe, M.; Brookes, A.J.; Budin-Ljøsne, I.; Chisholm, R.; Cox, D.; Deschênes, M. Toward a roadmap in global biobanking for health. Eur. J. Hum. Genet. 2012, 20, 1105–1111. [Google Scholar] [CrossRef] [PubMed]
- Zawati, M.H.; Tassé, A.M.; Mendy, M.; Caboux, E.; Lang, M.; on behalf of Biobank and Cohort Building Network Members. Barriers and opportunities in consent and access procedures in low- and middle-income country biobanks: Meeting notes from the BCNet training and general assembly. Biopreservation Biobanking 2018, 16, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Vodosin, P.; Jorgensen, A.K.; Mendy, M.; Kozlakidis, Z.; Caboux, E.; Zawati, M.H. A Review of Regulatory Frameworks Governing Biobanking in the Low and Middle Income Member Countries of BCNet. Biopreservation Biobanking 2021, 19, 444–452. [Google Scholar] [CrossRef] [PubMed]
- Baldo, C.; Viotti, V.; Maioli, E.; Mogni, M.; Castagnetta, M.; Cavani, S.; Piombo, G.; Coviello, D. Galliera Genetic Bank: A DNA and cell line biobank from patients affected by genetic diseases. Open J. Bioresour. 2016, 3, e1–e5. [Google Scholar] [CrossRef]
- Bao, E.L.; Cheng, A.N.; Sankaran, V.G. The genetics of human hematopoiesis and its disruption in disease. EMBO Mol. Med. 2019, 11, e10316. [Google Scholar] [CrossRef] [PubMed]
- Weatherall, D.J. Phenotype-genotype relationships in monogenic disease: Lessons from the thalassaemias. Nat. Rev. Genet. 2001, 2, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Bunn, H.F.; Noguchi, C.T.; Hofrichter, J.; Schechter, G.P.; Schechter, A.N.; Eaton, W.A. Molecular and cellular pathogenesis of hemoglobin Sickle Cell disease. Proc. Natl. Acad. Sci. USA 1982, 79, 7527–7531. [Google Scholar] [CrossRef] [PubMed]
- Rao, E.; Kumar Chandraker, S.; Misha Singh, M.; Kumar, R. Global distribution of β-thalassemia mutations: An update. Gene 2024, 896, 148022. [Google Scholar] [CrossRef] [PubMed]
- Piel, F.B.; Patil, A.P.; Howes, R.E.; Nyangiri, O.A.; Gething, P.W.; Williams, T.N.; Weatherall, D.J.; Hay, S.I. Global distribution of the sickle cell gene and geographical confirmation of the malaria hypothesis. Nat. Commun. 2010, 1, 104. [Google Scholar] [CrossRef] [PubMed]
- Richardson, T.; Rice, M.; Haumschild, R.; Hoffer, D.; Morana, S.; Watkins, J. Optimizing the management of inherited blood disorders in a changing market: Findings from the AMCP Market Insights Program. J. Manag. Care Spec. Pharm. 2024, 30, S1–S12. [Google Scholar] [CrossRef] [PubMed]
- Musallam, K.M.; Viprakasit, V.; Lombard, L.; Gilroy, K.; Rane, A.; Vinals, L.; Tam, C.; Rizzo, M.; Coates, T.D. Systematic review and evidence gap assessment of the clinical, quality of life, and economic burden of alpha-thalassemia. EJHaem. 2024, 5, 541–547. [Google Scholar] [CrossRef] [PubMed]
- Gamaleldin, M.; Abraham, I.; Meabed, M.; Elberry, A.; Abdelhalim, S.; Hussein, A.; Waggas, D.; Hussein, R. Cost-effectiveness analysis of Manuka honey-Omega-3 combination treatments in treating oxidative stress of pediatric β-thalassemia major. Eur. Rev. Med. Pharmacol. Sci. 2024, 28, 1144–1154. [Google Scholar] [PubMed]
- Cosenza, L.C.; Breda, L.; Breveglieri, G.; Zuccato, C.; Finotti, A.; Lampronti, I.; Borgatti, M.; Chiavilli, F.; Gamberini, M.R.; Satta, S.; et al. A validated cellular biobank for β-thalassemia. J. Transl. Med. 2016, 14, 255. [Google Scholar] [CrossRef]
- Gambari, R.; Waziri, A.D.; Goonasekera, H.; Peprah, E. Pharmacogenomics of Drugs Used in β-Thalassemia and Sickle-Cell Disease: From Basic Research to Clinical Applications. Int. J. Mol. Sci. 2024, 25, 4263. [Google Scholar] [CrossRef]
- Sripichai, O.; Fucharoen, S. Fetal hemoglobin regulation in β-thalassemia: Heterogeneity, modifiers and therapeutic approaches. Expert. Rev. Hematol. 2016, 9, 1129–1137. [Google Scholar] [CrossRef] [PubMed]
- Hashemi, Z.; Ebrahimzadeh, M.A. Hemoglobin F (HbF) Inducers; History, Structure and Efficacies. Mini Rev. Med. Chem. 2022, 22, 52–68. [Google Scholar] [CrossRef]
- Langer, A.L.; Esrick, E.B. β-Thalassemia: Evolving treatment options beyond transfusion and iron chelation. Hematol. Am. Soc. Hematol. Educ. Program 2021, 2021, 600–606. [Google Scholar] [CrossRef]
- Bou-Fakhredin, R.; De Franceschi, L.; Motta, I.; Cappellini, M.D.; Taher, A.T. Pharmacological Induction of Fetal Hemoglobin in β-Thalassemia and Sickle Cell Disease: An Updated Perspective. Pharmaceuticals 2022, 15, 753. [Google Scholar] [CrossRef] [PubMed]
- Foong, W.C.; Loh, C.K.; Ho, J.J.; Lau, D.S. Foetal haemoglobin inducers for reducing blood transfusion in non-transfusion-dependent beta-thalassaemias. Cochrane Database Syst. Rev. 2023, 1, CD013767. [Google Scholar] [PubMed]
- CA22119 - Haemoglobinopathies in European Liaison of Medicine and Science (HELIOS). Available online: https://www.cost.eu/actions/CA22119/ (accessed on 9 December 2024).
- Kountouris, P.; Stephanou, C.; Archer, N.; Bonifazi, F.; Giannuzzi, V.; Kuo, K.H.M.; Maggio, A.; Makani, J.; Mañú-Pereira, M.D.M.; Michailidou, K.; et al. The International Hemoglobinopathy Research Network (INHERENT): An international initiative to study the role of genetic modifiers in hemoglobinopathies. Am. J. Hematol. 2021, 96, E416–E420. [Google Scholar] [CrossRef] [PubMed]
- Petelina, T.I.; Musikhina, N.A.; Avdeeva, K.S.; Shcherbinina, A.E.; Leonovich, S.V.; Zueva, E.V.; Garanina, V.D.; Gultiaeva, E.P.; Yaroslavskaya, E.I.; Kalyuzhnaya, E.N.; et al. Estimation of erythrocyte parameters of general blood analysis in patients with SARS-CoV-2 -associated pneumonia. Clin. Lab. Diagn. 2022, 67, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Delaye, J.B.; Alarcan, H.; Vallet, N.; Veyrat-Durebex, C.; Bernard, L.; Hérault, O.; Ropert, M.; Marlet, J.; Gyan, E.; Andres, C.; et al. Specific changes of erythroid regulators and hepcidin in patients infected by SARS-COV-2. J. Investig. Med. 2022, 70, 934–938. [Google Scholar] [CrossRef] [PubMed]
- Elahi, S. Hematopoietic responses to SARS-CoV-2 infection. Cell Mol. Life Sci. 2022, 79, 187. [Google Scholar] [CrossRef] [PubMed]
- Allahyani, M.A.; Aljuaid, A.A.; Almehmadi, M.M.; Alghamdi, A.A.; Halawani, I.F.; Aldairi, A.F.; Alharbi, A.M.; Albshri, M.H.; Mutwalli, A.A.; Alhazmi, A.S. Detection of erythroid progenitors and erythrocytopathies in patients with severe COVID-19 disease. Saudi Med. J. 2022, 43, 899–906. [Google Scholar] [CrossRef]
- Saito, S.; Shahbaz, S.; Sligl, W.; Osman, M.; Tyrrell, D.L.; Elahi, S. Differential Impact of SARS-CoV-2 Isolates, Namely, the Wuhan Strain, Delta, and Omicron Variants on Erythropoiesis. Microbiol. Spectr. 2022, 10, e0173022. [Google Scholar] [CrossRef] [PubMed]
- Girón-Pérez, D.A.; Nava-Piedra, U.N.; Esquivel-Esparza, Z.E.; Benitez-Trinidad, A.B.; Barcelos-Garcia, R.G.; Vázquez-Pulido, E.Y.; Toledo-Ibarra, G.A.; Ventura-Ramón, G.H.; Covantes-Rosales, C.E.; Barajas-Carrillo, V.W.; et al. Hematologic analysis of hospitalized patients and outpatients infected with SARS-CoV-2 and possible use as a prognostic biomarker. Exp. Hematol. 2023, 119–120, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Shahbaz, S.; Xu, L.; Osman, M.; Sligl, W.; Shields, J.; Joyce, M.; Tyrrell, D.L.; Oyegbami, O.; Elahi, S. Erythroid precursors and progenitors suppress adaptive immunity and get invaded by SARS-CoV-2. Stem Cell Rep. 2021, 16, 1165–1181. [Google Scholar] [CrossRef] [PubMed]
- Huerga Encabo, H.; Grey, W.; Garcia-Albornoz, M.; Wood, H.; Ulferts, R.; Aramburu, I.V.; Kulasekararaj, A.G.; Mufti, G.; Papayannopoulos, V.; Beale, R.; et al. Human Erythroid Progenitors Are Directly Infected by SARS-CoV-2: Implications for Emerging Erythropoiesis in Severe COVID-19 Patients. Stem Cell Rep. 2021, 16, 428–436. [Google Scholar] [CrossRef] [PubMed]
- Balzanelli, M.G.; Distratis, P.; Dipalma, G.; Vimercati, L.; Inchingolo, A.D.; Lazzaro, R.; Aityan, S.K.; Maggiore, M.E.; Mancini, A.; Laforgia, R.; et al. Sars-CoV-2 Virus Infection May Interfere CD34+ Hematopoietic Stem Cells and Megakaryocyte-Erythroid Progenitors Differentiation Contributing to Platelet Defection towards Insurgence of Thrombocytopenia and Thrombophilia. Microorganisms 2021, 9, 1632. [Google Scholar] [CrossRef]
- Kronstein-Wiedemann, R.; Stadtmüller, M.; Traikov, S.; Georgi, M.; Teichert, M.; Yosef, H.; Wallenborn, J.; Karl, A.; Schütze, K.; Wagner, M.; et al. SARS-CoV-2 Infects Red Blood Cell Progenitors and Dysregulates Hemoglobin and Iron Metabolism. Stem Cell Rev. Rep. 2022, 18, 1809–1821. [Google Scholar] [CrossRef] [PubMed]
- Bernardes, J.P.; Mishra, N.; Tran, F.; Bahmer, T.; Best, L.; Blase, J.I.; Bordoni, D.; Franzenburg, J.; Geisen, U.; Josephs-Spaulding, J.; et al. Longitudinal Multi-omics Analyses Identify Responses of Megakaryocytes, Erythroid Cells, and Plasmablasts as Hallmarks of Severe COVID-19. Immunity 2020, 53, 1296–1314.e9. [Google Scholar] [CrossRef]
- Eltobgy, M.; Johns, F.; Farkas, D.; Leuenberger, L.; Cohen, S.P.; Ho, K.; Karow, S.; Swoope, G.; Pannu, S.; Horowitz, J.C.; et al. Longitudinal transcriptomic analysis reveals persistent enrichment of iron homeostasis and erythrocyte function pathways in severe COVID-19 ARDS. Front. Immunol. 2024, 15, 1397629. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Shu, X.; Deng, R.; Yang, Z.; Shu, M.; Ou, X.; Zhang, X.; Wu, Z.; Zeng, H.; Shao, L. Transcriptome Changes of Hematopoietic Stem and Progenitor Cells in the Peripheral Blood of COVID-19 Patients by scRNA-seq. Int. J. Mol. Sci. 2023, 24, 10878. [Google Scholar] [CrossRef]
- Simeon-Dubach, D.; Zeisberger, S.M.; Hoerstrup, S.P. Quality Assurance in Biobanking for Pre-Clinical Research. Transfus. Med. Hemother. 2016, 43, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Coppola, L.; Grimaldi, A.M.; Sarnacchiaro, G.; di Fasano, M.S.; Smaldone, G.; Salvatore, M. An overview of Synlab SDN Biobank’s quality control system. Sci. Rep. 2024, 14, 19303. [Google Scholar] [CrossRef]
- Mouttham, L.; Garrison, S.J.; Archer, D.L.; Castelhano, M.G. A Biobank’s Journey: Implementation of a Quality Management System and Accreditation to ISO 20387. Biopreservation Biobanking 2021, 19, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Caixeiro, N.J.; Lai, K.; Lee, C.S. Quality assessment and preservation of RNA from biobank tissue specimens: A systematic review. J. Clin. Pathol. 2016, 69, 260–265. [Google Scholar] [CrossRef] [PubMed]
- Zuccato, C.; Cosenza, L.C.; Tupini, C.; Finotti, A.; Sacchetti, G.; Simoni, D.; Gambari, R.; Lampronti, I. New Synthetic Isoxazole Derivatives Acting as Potent Inducers of Fetal Hemoglobin in Erythroid Precursor Cells Isolated from β-Thalassemic Patients. Molecules 2023, 29, 8. [Google Scholar] [CrossRef] [PubMed]
- Szuberski, J.; Oliveira, J.L.; Hoyer, J.D. A comprehensive analysis of hemoglobin variants by high-performance liquid chromatography (HPLC). Int. J. Lab. Hematol. 2012, 34, 594–604. [Google Scholar] [CrossRef] [PubMed]
- Joutovsky, A.; Hadzi-Nesic, J.; Nardi, M.A. HPLC retention time as a diagnostic tool for hemoglobin variants and hemoglobinopathies: A study of 60000 samples in a clinical diagnostic laboratory. Clin Chem. 2004, 50, 1736–1747. [Google Scholar] [CrossRef] [PubMed]
- Zuccato, C.; Cosenza, L.C.; Zurlo, M.; Gasparello, J.; Papi, C.; D’Aversa, E.; Breveglieri, G.; Lampronti, I.; Finotti, A.; Borgatti, M.; et al. Expression of γ-globin genes in β-thalassemia patients treated with sirolimus: Results from a pilot clinical trial (Sirthalaclin). Ther. Adv. Hematol. 2022, 13, 20406207221100648. [Google Scholar] [CrossRef]
- Cosenza, L.C.; Gasparello, J.; Romanini, N.; Zurlo, M.; Zuccato, C.; Gambari, R.; Finotti, A. Efficient CRISPR-Cas9-based genome editing of β-globin gene on erythroid cells from homozygous β039-thalassemia patients. Mol. Ther. Methods Clin. Dev. 2021, 21, 507–523. [Google Scholar] [CrossRef] [PubMed]
- Cosenza, L.C.; Zuccato, C.; Zurlo, M.; Gambari, R.; Finotti, A. Co-Treatment of Erythroid Cells from β-Thalassemia Patients with CRISPR-Cas9-Based β039-Globin Gene Editing and Induction of Fetal Hemoglobin. Genes 2022, 13, 1727. [Google Scholar] [CrossRef] [PubMed]
- Zuccato, C.; Cosenza, L.C.; Zurlo, M.; Lampronti, I.; Borgatti, M.; Scapoli, C.; Gambari, R.; Finotti, A. Treatment of Erythroid Precursor Cells from β-Thalassemia Patients with Cinchona Alkaloids: Induction of Fetal Hemoglobin Production. Int. J. Mol. Sci. 2021, 22, 13433. [Google Scholar] [CrossRef] [PubMed]
- Cosenza, L.C.; Marzaro, G.; Zurlo, M.; Gasparello, J.; Zuccato, C.; Finotti, A.; Gambari, R. Inhibitory effects of SARS-CoV-2 spike protein and BNT162b2 vaccine on erythropoietin-induced globin gene expression in erythroid precursor cells from patients with β-thalassemia. Exp. Hematol. 2024, 129, 104128. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Horby, P.W.; Hayden, F.G.; Gao, G.F. A novel coronavirus outbreak of global health concern. Lancet 2020, 395, 470–473. [Google Scholar] [CrossRef] [PubMed]
- Peng, M. Outbreak of COVID-19: An emerging global pandemic threat. Biomed. Pharmacother. 2020, 129, 110499. [Google Scholar] [CrossRef]
- Gatto, M.; Bertuzzo, E.; Mari, L.; Miccoli, S.; Carraro, L.; Casagrandi, R.; Rinaldo, A. Spread and dynamics of the COVID-19 epidemic in Italy: Effects of emergency containment measures. Proc. Natl. Acad. Sci. USA 2020, 117, 10484–10491. [Google Scholar] [CrossRef] [PubMed]
- Sahin, U.; Muik, A.; Vogler, I.; Derhovanessian, E.; Kranz, L.M.; Vormehr, M.; Quandt, J.; Bidmon, N.; Ulges, A.; Baum, A.; et al. BNT162b2 vaccine induces neutralizing antibodies and poly-specific T cells in humans. Nature 2021, 595, 572–577. [Google Scholar] [CrossRef]
- Pegu, A.; O’Connell, S.E.; Schmidt, S.D.; O’Dell, S.; Talana, C.A.; Lai, L.; Albert, J.; Anderson, E.; Bennett, H.; Corbett, K.S.; et al. Durability of mRNA-1273 vaccine-induced antibodies against SARS-CoV-2 variants. Science 2021, 373, 1372–1377. [Google Scholar] [CrossRef] [PubMed]
- Ratajczak, M.Z.; Kucia, M. SARS-CoV-2 infection and overactivation of Nlrp3 inflammasome as a trigger of cytokine "storm" and risk factor for damage of hematopoietic stem cells. Leukemia 2020, 34, 1726–1729. [Google Scholar] [CrossRef] [PubMed]
- Ropa, J.; Cooper, S.; Capitano, M.L.; Van’t Hof, W.; Broxmeyer, H.E. Human Hematopoietic Stem, Progenitor, and Immune Cells Respond Ex Vivo to SARS-CoV-2 Spike Protein. Stem Cell Rev. Rep. 2021, 17, 253–265. [Google Scholar] [CrossRef]
- Kucia, M.; Ratajczak, J.; Bujko, K.; Adamiak, M.; Ciechanowicz, A.; Chumak, V.; Brzezniakiewicz-Janus, K.; Ratajczak, M. An evidence that SARS-CoV-2/COVID-19 spike protein (SP) damages hematopoietic stem/progenitor cells in the mechanism of pyroptosis in Nlrp3 inflammasome-dependent manner. Leukemia 2021, 35, 3026–3029. [Google Scholar] [CrossRef]
- Estep, B.K.; Kuhlmann, C.J.; Osuka, S.; Suryavanshi, G.W.; Nagaoka-Kamata, Y.; Samuel, C.N.; Blucas, M.T.; Jepson, C.E.; Goepfert, P.A.; Kamata, M. Skewed fate and hematopoiesis of CD34+ HSPCs in umbilical cord blood amid the COVID-19 pandemic. iScience 2022, 25, 105544. [Google Scholar] [CrossRef] [PubMed]
- Trougakos, I.P.; Terpos, E.; Alexopoulos, H.; Politou, M.; Paraskevis, D.; Scorilas, A.; Kastritis, E.; Andreakos, E.; Dimopoulos, M.A. Adverse effects of COVID-19 mRNA vaccines: The spike hypothesis. Trends Mol. Med. 2022, 28, 542–554. [Google Scholar] [CrossRef]
- Zika, E.; Paci, D.; Schulte in den Bäumen, T.; Braun, A.; Rijkers-Defrasne, S.; Deschênes, M.; Fortier, I.; Laage-Hellman, J.; A Scerri, C.; Ibarreta Ruiz, D. Biobanks in Europe: Prospects for Harmonisation and Networking; JRC57831; Publications Office of the European Union: Luxembourg, 2010. [Google Scholar]
- Tzortzatou-Nanopoulou, O.; Akyüz, K.; Goisauf, M.; Kozera, Ł.; Mežinska, S.; Th Mayrhofer, M.; Slokenberga, S.; Reichel, J.; Croxton, T.; Ziaka, A.; et al. Ethical, legal, and social implications in research biobanking: A checklist for navigating complexity. Dev. World Bioeth. 2024, 24, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Gottweis, H.; Zatloukal, K. Biobank governance: Trends and perspectives. Pathobiology 2007, 74, 206–211. [Google Scholar] [CrossRef] [PubMed]
- Ducournau, P.; Strand, R. Trust, Distrust and Co-production: The Relationship Between Research Biobanks and Donors. In The Ethics of Research Biobanking; Solbakk, J., Holm, S., Hofmann, B., Eds.; Springer: Boston, MA, USA, 2009. [Google Scholar] [CrossRef]
- Hoeyer, K.; Olofsson, B.; Mjörndal, T.; Lynöe, N. The Ethics of Research Using Biobanks: Reason to Question the Importance Attributed to Informed Consent. Arch. Intern. Med. 2005, 165, 97–100. [Google Scholar] [CrossRef] [PubMed]
- McMahon, A.M.; Kolawole, O.I. Biobank donation in search of public benefits and the potential impact of intellectual property rights over access to health-technologies developed: A focus on the bioethical implications. Med. Law Rev. 2024, 32, 205–228. [Google Scholar] [CrossRef] [PubMed]
- Jordan, M.; Liddicoat, J.; Liddell, K. An empirical study of large, human biobanks: Intellectual property policies and financial conditions for access. J. Law. Biosci. 2021, 8, lsab018. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| THALAMOSS Project: Induction of HbF with LMW Drugs | Project: Induction of HbA with Readthrough-Based Molecules | Project: β039 CRISPR-Cas9 Genome Editing System | Project: Effects of SARS-CoV-2 Spike Protein on ErPCs | ||||
|---|---|---|---|---|---|---|---|
| Number of patients | Number of Vials | Number of patients | Number of Vials | Number of patients | Number of Vials | Number of patients | Number of Vials |
| 84 | 379 | 5 | 10 | 11 | 27 | 6 | 12 |
| Publications: Zuccato et al., 2021 [62]; Zuccato et al., 2022 [59]; Zuccato et al., 2023 [56] | Unpublished | Publications: Cosenza et al., 2021 [60]; Cosenza et al., 2022 [61] | Publications: Cosenza et al., 2024 [63] | ||||
| Cosenza et al., 2016 [26] | Present Composition of the Biobank | |||
|---|---|---|---|---|
| Genotype/Phenotype | Number of Patients | Number of Vials | Number of Patients | Number of Vials |
| β039/β039 | 29 | 260 | 103 | 421 |
| β+IVSI-110/β+IVSI-110 | 8 | 81 | 9 | 65 |
| β+IVSI-110/β039 | 17 | 191 | 25 | 51 |
| β+IVSI-6/β+IVSI-6 | 2 | 15 | 3 | 28 |
| β+IVSI-6/β039 | 4 | 67 | 5 | 24 |
| β+IVSI-110/β+IVSI-6 | 1 | 11 | 1 | 8 |
| β+IVSI-110/β+IVSI-1 | 1 | 7 | 1 | 7 |
| SCD/SCD | 2 | 13 | 3 | 17 |
| SCD/β039 | 1 | 7 | 2 | 10 |
| SCD/β+IVSI-110 | 1 | 5 | 2 | 8 |
| SCD/β+IVSI-6 | 1 | 7 | 1 | 7 |
| SCD/β+IVSI-1 | 2 | 20 | 2 | 15 |
| Others | 11 | 95 | 64 | 329 |
| Total | 80 | 779 | 221 | 990 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gambari, R.; Gamberini, M.R.; Cosenza, L.C.; Zuccato, C.; Finotti, A. A β-Thalassemia Cell Biobank: Updates, Further Validation in Genetic and Therapeutic Research and Opportunities During (and After) the COVID-19 Pandemic. J. Clin. Med. 2025, 14, 289. https://doi.org/10.3390/jcm14010289
Gambari R, Gamberini MR, Cosenza LC, Zuccato C, Finotti A. A β-Thalassemia Cell Biobank: Updates, Further Validation in Genetic and Therapeutic Research and Opportunities During (and After) the COVID-19 Pandemic. Journal of Clinical Medicine. 2025; 14(1):289. https://doi.org/10.3390/jcm14010289
Chicago/Turabian StyleGambari, Roberto, Maria Rita Gamberini, Lucia Carmela Cosenza, Cristina Zuccato, and Alessia Finotti. 2025. "A β-Thalassemia Cell Biobank: Updates, Further Validation in Genetic and Therapeutic Research and Opportunities During (and After) the COVID-19 Pandemic" Journal of Clinical Medicine 14, no. 1: 289. https://doi.org/10.3390/jcm14010289
APA StyleGambari, R., Gamberini, M. R., Cosenza, L. C., Zuccato, C., & Finotti, A. (2025). A β-Thalassemia Cell Biobank: Updates, Further Validation in Genetic and Therapeutic Research and Opportunities During (and After) the COVID-19 Pandemic. Journal of Clinical Medicine, 14(1), 289. https://doi.org/10.3390/jcm14010289