Vitamin A Positively Correlates with Secretory Immunoglobulin A: A Cross-Sectional Study in Omicron COVID-19 Outpatients

 , , ,
, , ,  , ,
, ,  ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design and Patients
2.2. Data Collection
2.3. Analysis of Serum Samples
2.4. Analysis of Salivary Samples
2.5. Assessment of Nutrient Intake
2.6. Statistical Analysis
3. Results
3.1. Clinical, Biochemical, and Nutritional Characteristics
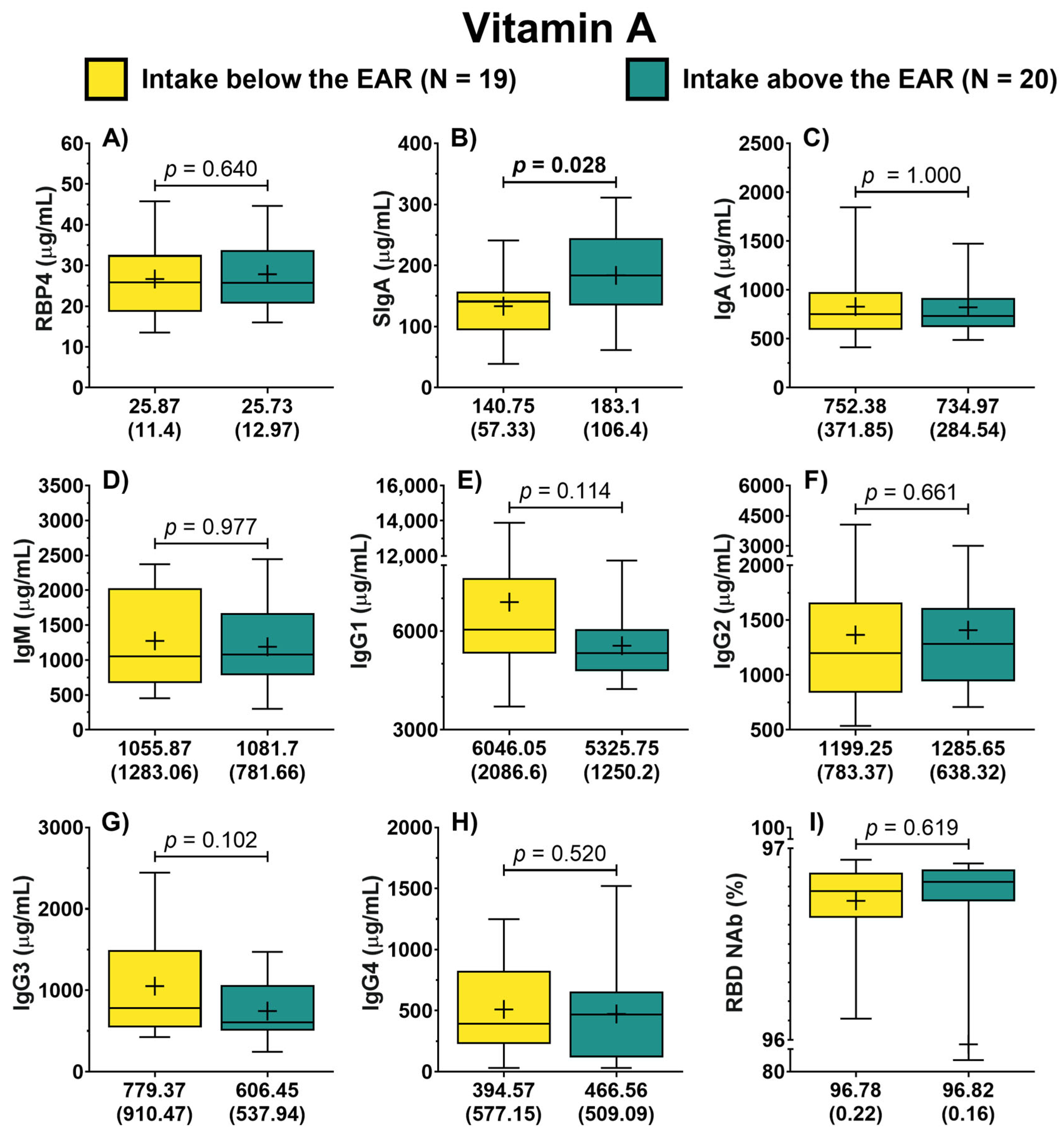
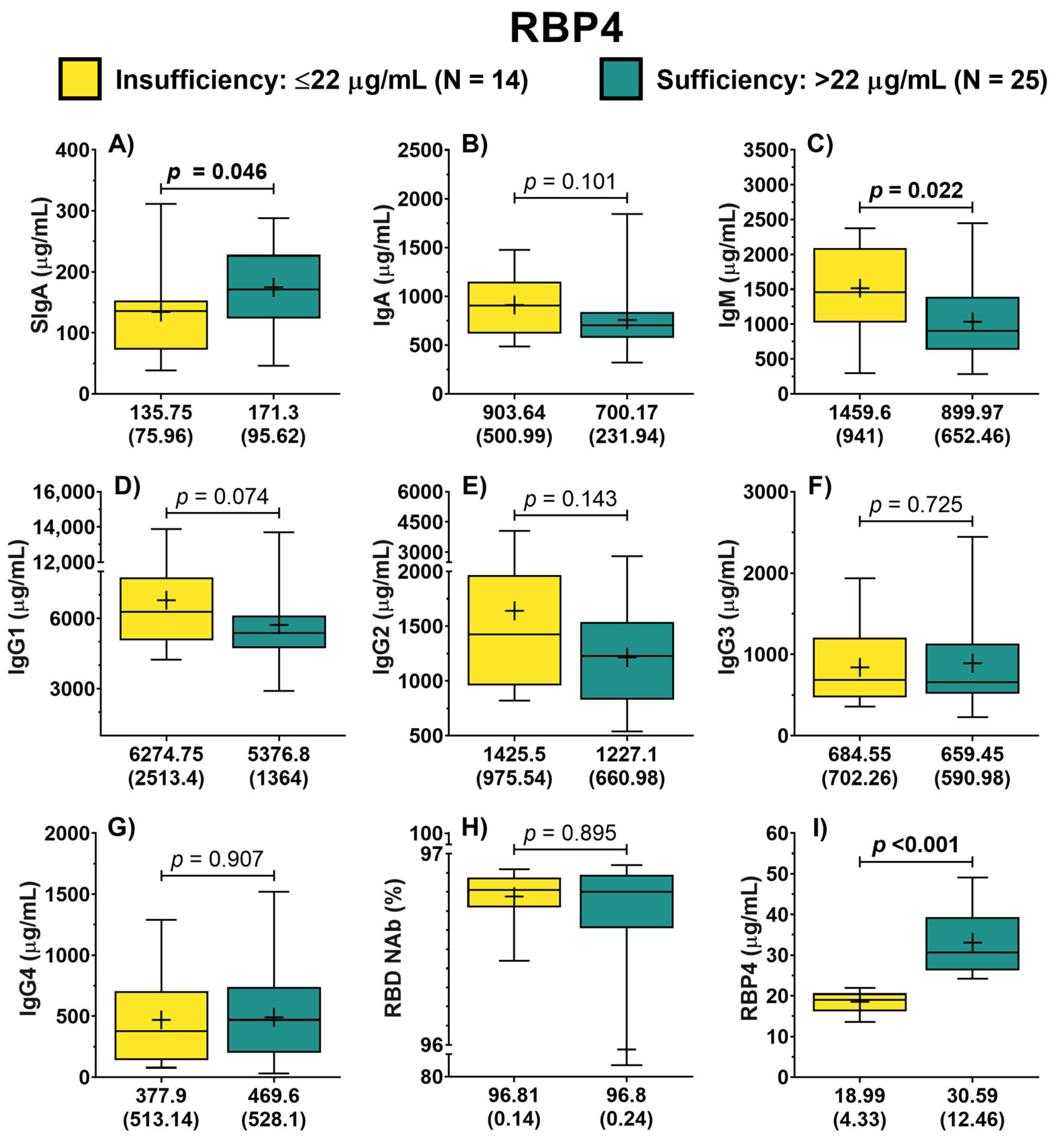
3.2. Antibodies by Vitamin A Nutritional Status
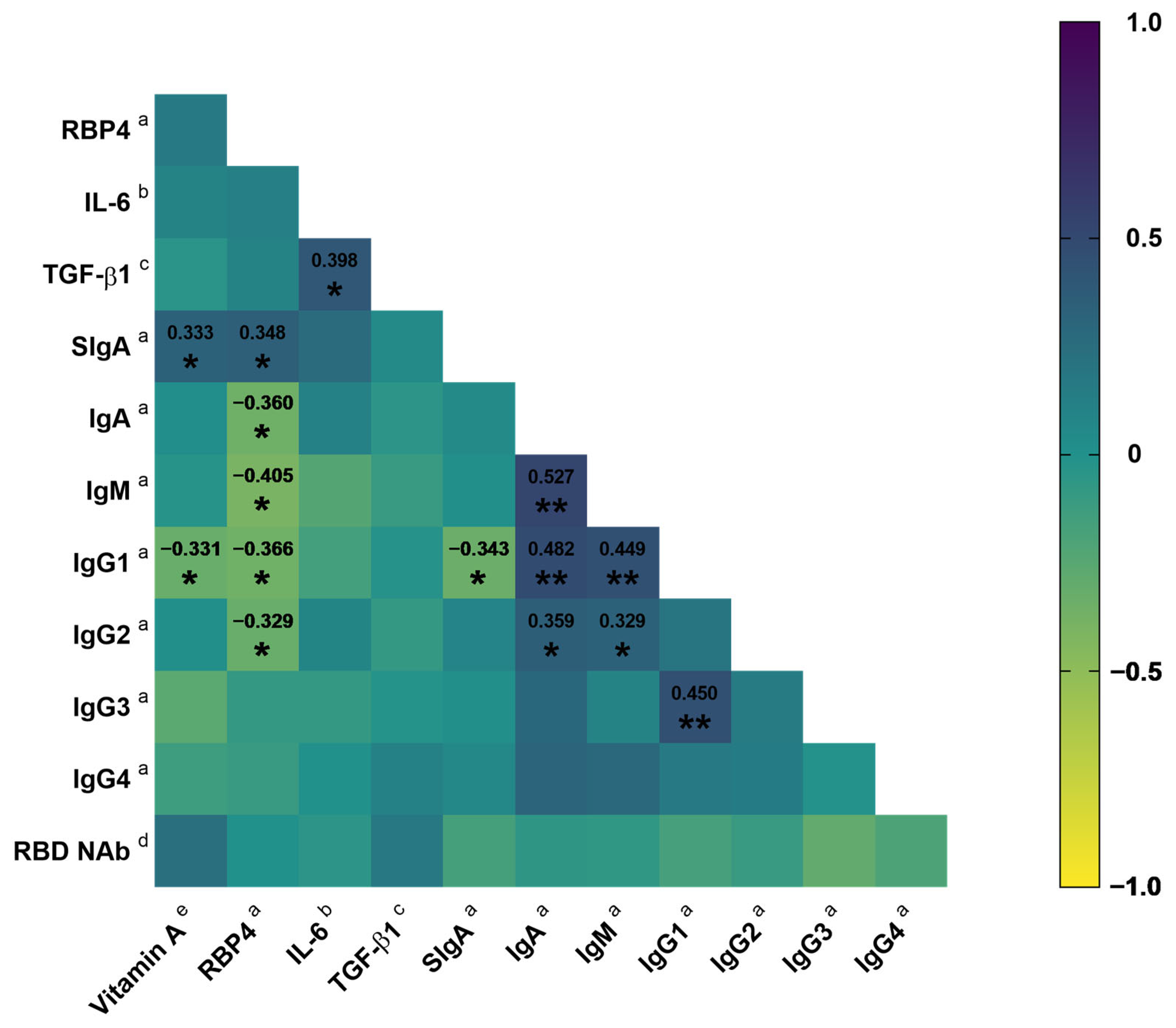
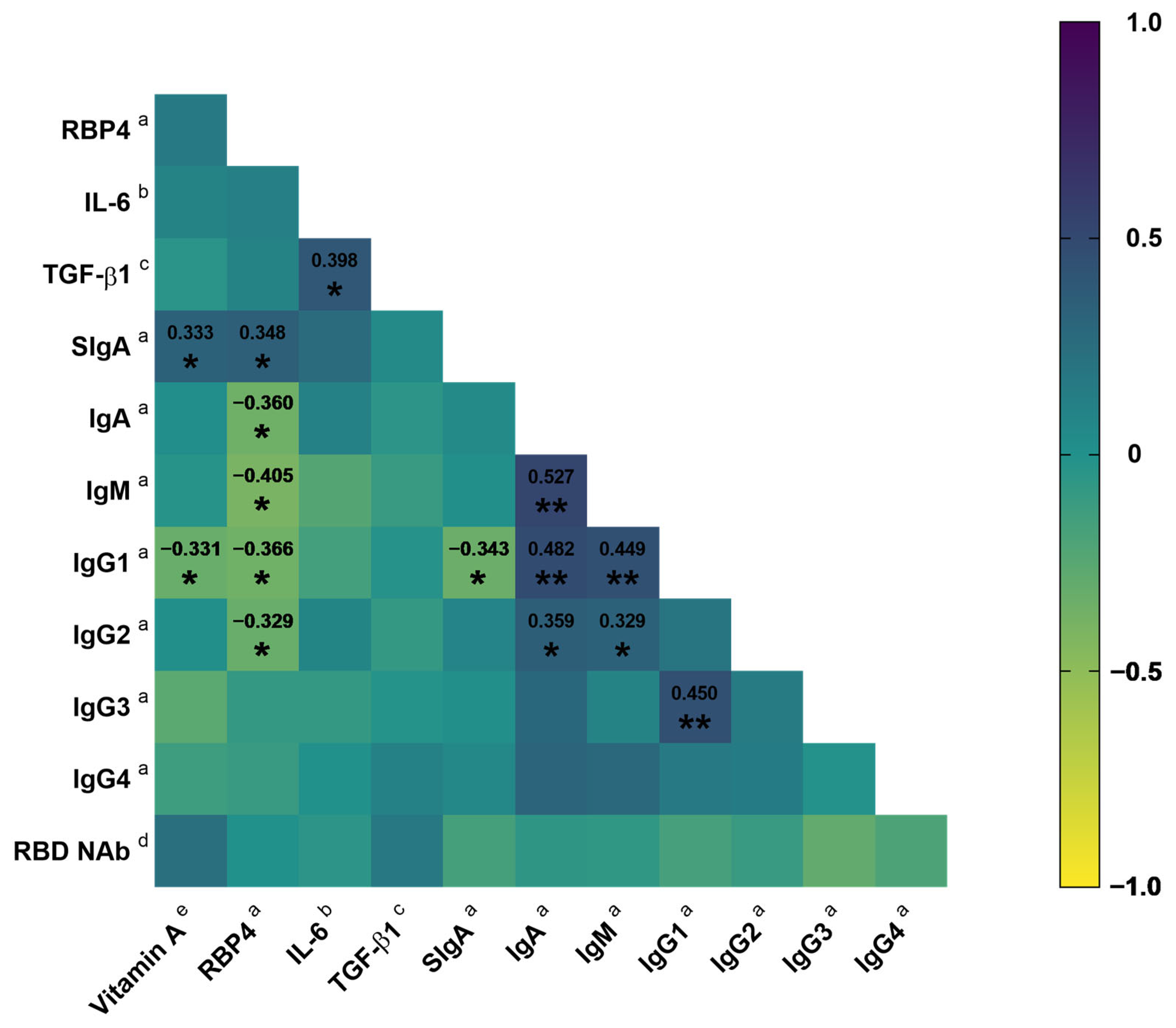
3.3. Association of Vitamin A Nutritional Status with SIgA
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jin, X.; Ren, J.; Li, R.; Gao, Y.; Zhang, H.; Li, J.; Zhang, J.; Wang, X.; Wang, G. Global Burden of Upper Respiratory Infections in 204 Countries and Territories, from 1990 to 2019. eClinicalMedicine 2021, 37, 100986. [Google Scholar] [CrossRef] [PubMed]
- Safiri, S.; Mahmoodpoor, A.; Kolahi, A.-A.; Nejadghaderi, S.A.; Sullman, M.J.M.; Mansournia, M.A.; Ansarin, K.; Collins, G.S.; Kaufman, J.S.; Abdollahi, M. Global Burden of Lower Respiratory Infections during the Last Three Decades. Front. Public Health 2023, 10, 1028525. [Google Scholar] [CrossRef]
- Calderaro, A.; Buttrini, M.; Farina, B.; Montecchini, S.; De Conto, F.; Chezzi, C. Respiratory Tract Infections and Laboratory Diagnostic Methods: A Review with A Focus on Syndromic Panel-Based Assays. Microorganisms 2022, 10, 1856. [Google Scholar] [CrossRef] [PubMed]
- Polatoğlu, I.; Oncu-Oner, T.; Dalman, I.; Ozdogan, S. COVID-19 in Early 2023: Structure, Replication Mechanism, Variants of SARS-CoV-2, Diagnostic Tests, and Vaccine & Drug Development Studies. MedComm 2023, 4, e228. [Google Scholar] [CrossRef] [PubMed]
- Bonanni, P.; Ceddia, F.; Dawson, R. A Call to Action: Current Challenges and Considerations for COVID-19 Vaccination in Immunocompromised Populations. J. Infect. Dis. 2023, 228, S70–S76. [Google Scholar] [CrossRef] [PubMed]
- Velikova, T.; Snegarova, V.; Kukov, A.; Batselova, H.; Mihova, A.; Nakov, R. Gastrointestinal Mucosal Immunity and COVID-19. World J. Gastroenterol. 2021, 27, 5047–5059. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Jin, L.; Chen, T. The Effects of Secretory IgA in the Mucosal Immune System. BioMed Res. Int. 2020, 2020, 2032057. [Google Scholar] [CrossRef] [PubMed]
- Sano, K.; Bhavsar, D.; Singh, G.; Floda, D.; Srivastava, K.; Gleason, C.; PARIS Study Group; Carreño, J.M.; Simon, V.; Krammer, F.; et al. SARS-CoV-2 Vaccination Induces Mucosal Antibody Responses in Previously Infected Individuals. Nat. Commun. 2022, 13, 5135. [Google Scholar] [CrossRef]
- Wang, Z.; Lorenzi, J.C.C.; Muecksch, F.; Finkin, S.; Viant, C.; Gaebler, C.; Cipolla, M.; Hoffmann, H.-H.; Oliveira, T.Y.; Oren, D.A.; et al. Enhanced SARS-CoV-2 Neutralization by Dimeric IgA. Sci. Transl. Med. 2021, 13, eabf1555. [Google Scholar] [CrossRef]
- Russell, M.W.; Mestecky, J. Mucosal Immunity: The Missing Link in Comprehending SARS-CoV-2 Infection and Transmission. Front. Immunol. 2022, 13, 957107. [Google Scholar] [CrossRef]
- Carazo, A.; Macáková, K.; Matoušová, K.; Krčmová, L.K.; Protti, M.; Mladěnka, P. Vitamin A Update: Forms, Sources, Kinetics, Detection, Function, Deficiency, Therapeutic Use and Toxicity. Nutrients 2021, 13, 1703. [Google Scholar] [CrossRef] [PubMed]
- Bos, A.; Van Egmond, M.; Mebius, R. The Role of Retinoic Acid in the Production of Immunoglobulin A. Mucosal Immunol. 2022, 15, 562–572. [Google Scholar] [CrossRef] [PubMed]
- Huiming, Y.; Chaomin, W.; Meng, M. Vitamin A for Treating Measles in Children. In Cochrane Database of Systematic Reviews; The Cochrane Collaboration, Ed.; John Wiley & Sons, Ltd: Chichester, UK, 2005; p. CD001479. [Google Scholar]
- Amimo, J.O.; Michael, H.; Chepngeno, J.; Raev, S.A.; Saif, L.J.; Vlasova, A.N. Immune Impairment Associated with Vitamin A Deficiency: Insights from Clinical Studies and Animal Model Research. Nutrients 2022, 14, 5038. [Google Scholar] [CrossRef] [PubMed]
- Sirisinha, S.; Darip, M.D.; Moongkarndi, P.; Ongsakul, M.; Lamb, A.J. Impaired Local Immune Response in Vitamin A-Deficient Rats. Clin. Exp. Immunol. 1980, 40, 127–135. [Google Scholar] [PubMed]
- Cui, D.; Stephensen, C.B.; Moldoveanu, Z. High-Level Dietary Vitamin A Enhances T-Helper Type 2 Cytokine Production and Secretory Immunoglobulin A Response to Influenza A Virus Infection in BALB/c Mice. J. Nutr. 2000, 130, 1132–1139. [Google Scholar] [CrossRef] [PubMed]
- Surman, S.L.; Jones, B.G.; Sealy, R.E.; Rudraraju, R.; Hurwitz, J.L. Oral Retinyl Palmitate or Retinoic Acid Corrects Mucosal IgA Responses toward an Intranasal Influenza Virus Vaccine in Vitamin A Deficient Mice. Vaccine 2014, 32, 2521–2524. [Google Scholar] [CrossRef] [PubMed]
- Surman, S.L.; Penkert, R.R.; Jones, B.G.; Sealy, R.E.; Hurwitz, J.L. Vitamin Supplementation at the Time of Immunization with a Cold-Adapted Influenza Virus Vaccine Corrects Poor Mucosal Antibody Responses in Mice Deficient for Vitamins A and D. Clin. Vaccine Immunol. 2016, 23, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Patel, N.; Penkert, R.R.; Sealy, R.E.; Surman, S.L.; Jones, B.G.; Ringwald-Smith, K.; Ross, A.C.; Hurwitz, J.L. Retinol Binding Protein, Sunlight Hours, and the Influenza Virus-Specific Immune Response. Biomedicines 2022, 10, 2322. [Google Scholar] [CrossRef]
- Jones, B.G.; Oshansky, C.M.; Bajracharya, R.; Tang, L.; Sun, Y.; Wong, S.S.; Webby, R.; Thomas, P.G.; Hurwitz, J.L. Retinol Binding Protein and Vitamin D Associations with Serum Antibody Isotypes, Serum Influenza Virus-Specific Neutralizing Activities and Airway Cytokine Profiles. Clin. Exp. Immunol. 2016, 183, 239–247. [Google Scholar] [CrossRef]
- Muñoz-Valle, J.F.; Venancio-Landeros, A.A.; Sánchez-Sánchez, R.; Reyes-Díaz, K.; Galindo-Ornelas, B.; Hérnandez-Monjaraz, W.S.; García-Ríos, A.; García-Ortega, L.F.; Hernández-Bello, J.; Peña-Rodríguez, M.; et al. An Upgrade on the Surveillance System of SARS-CoV-2: Deployment of New Methods for Genetic Inspection. Int. J. Mol. Sci. 2022, 23, 3143. [Google Scholar] [CrossRef]
- González-Estevez, G.; Turrubiates-Hernández, F.J.; Herrera-Jiménez, L.E.; Sánchez-Zuno, G.A.; Herrera-Godina, M.G.; Muñoz-Valle, J.F. Association of Food Intake Quality with Vitamin D in SARS-CoV-2 Positive Patients from Mexico: A Cross-Sectional Study. Int. J. Environ. Res. Public Health 2021, 18, 7266. [Google Scholar] [CrossRef] [PubMed]
- Macedo-Ojeda, G.; Muñoz-Valle, J.F.; Yokogawa-Teraoka, P.; Machado-Sulbarán, A.C.; Loza-Rojas, M.G.; García-Arredondo, A.C.; Tejeda-Constantini, R.; Vega-Magaña, A.N.; González-Estevez, G.; García-Chagollán, M.; et al. COVID-19 Screening by Anti-SARS-CoV-2 Antibody Seropositivity: Clinical and Epidemiological Characteristics, Comorbidities, and Food Intake Quality. Int. J. Environ. Res. Public Health 2021, 18, 8995. [Google Scholar] [CrossRef] [PubMed]
- Machin, D.; Campbell, M.J.; Tan, S.B.; Tan, S.H. Sample Size Tables for Clinical Studies, 1st ed.; Wiley: Hoboken, NJ, USA, 2008; ISBN 978-1-4051-4650-0. [Google Scholar]
- De Pee, S.; Dary, O. Biochemical Indicators of Vitamin A Deficiency: Serum Retinol and Serum Retinol Binding Protein. J. Nutr. 2002, 132, 2895S–2901S. [Google Scholar] [CrossRef] [PubMed]
- Isho, B.; Abe, K.T.; Zuo, M.; Jamal, A.J.; Rathod, B.; Wang, J.H.; Li, Z.; Chao, G.; Rojas, O.L.; Bang, Y.M.; et al. Persistence of Serum and Saliva Antibody Responses to SARS-CoV-2 Spike Antigens in COVID-19 Patients. Sci. Immunol. 2020, 5, eabe5511. [Google Scholar] [CrossRef] [PubMed]
- Vizmanos-Lamotte, B.; López-Uriarte, P.J.; Hunot-Alexander, C.; Bernal-Orozco, M.F.; Rodríguez-Rocha, N.P.; Macedo-Ojeda, G.; Martínez-Lomelí, L.; Rovillé-Sausse, F. Álbum Fotográfico de Alimentos Mexicanos; Editorial Pandora: Guadalajara, Mexico, 2015; ISBN 978-607-96709-1-7. [Google Scholar]
- Willett, W.; Howe, G.; Kushi, L. Adjustment for Total Energy Intake in Epidemiologic Studies. Am. J. Clin. Nutr. 1997, 65, 1220S–1228S. [Google Scholar] [CrossRef]
- Institute of Medicine (U.S.) Committee to Review Dietary Reference Intakes for Vitamin D and Calcium. Dietary Reference Intakes for Vitamin D and Calcium; National Academies Press: Washington, DC, USA, 2011; ISBN 978-0-309-16394-1. [Google Scholar]
- Zhao, T.; Liu, S.; Zhang, R.; Zhao, Z.; Yu, H.; Pu, L.; Wang, L.; Han, L. Global Burden of Vitamin A Deficiency in 204 Countries and Territories from 1990–2019. Nutrients 2022, 14, 950. [Google Scholar] [CrossRef]
- Popkin, B.M.; Ng, S.W. The Nutrition Transition to a Stage of High Obesity and Noncommunicable Disease Prevalence Dominated by Ultra-processed Foods Is Not Inevitable. Obes. Rev. 2022, 23, e13366. [Google Scholar] [CrossRef]
- Martini, D.; Godos, J.; Bonaccio, M.; Vitaglione, P.; Grosso, G. Ultra-Processed Foods and Nutritional Dietary Profile: A Meta-Analysis of Nationally Representative Samples. Nutrients 2021, 13, 3390. [Google Scholar] [CrossRef]
- Marrón-Ponce, J.A.; Sánchez-Pimienta, T.G.; Rodríguez-Ramírez, S.; Batis, C.; Cediel, G. Ultra-processed Foods Consumption Reduces Dietary Diversity and Micronutrient Intake in the Mexican Population. J. Hum. Nutr. Diet. 2023, 36, 241–251. [Google Scholar] [CrossRef]
- Pedroza-Tobías, A.; Hernández-Barrera, L.; López-Olmedo, N.; García-Guerra, A.; Rodríguez-Ramírez, S.; Ramírez-Silva, I.; Villalpando, S.; Carriquiry, A.; Rivera, J.A. Usual Vitamin Intakes by Mexican Populations. J. Nutr. 2016, 146, 1866S–1873S. [Google Scholar] [CrossRef]
- Ramírez-Silva, I.; Rodríguez-Ramírez, S.; Barragán-Vázquez, S.; Castellanos-Gutiérrez, A.; Reyes-García, A.; Martínez-Piña, A.; Pedroza-Tobías, A. Prevalence of Inadequate Intake of Vitamins and Minerals in the Mexican Population Correcting by Nutrient Retention Factors, Ensanut 2016. Salud Pública México 2020, 62, 521–531. [Google Scholar] [CrossRef] [PubMed]
- Tanumihardjo, S.A.; Russell, R.M.; Stephensen, C.B.; Gannon, B.M.; Craft, N.E.; Haskell, M.J.; Lietz, G.; Schulze, K.; Raiten, D.J. Biomarkers of Nutrition for Development (BOND)—Vitamin A Review. J. Nutr. 2016, 146, 1816S–1848S. [Google Scholar] [CrossRef] [PubMed]
- Mohd, M.A.; Ahmad Norudin, N.A.; Muhammad, T.S.T. Transcriptional Regulation of Retinol Binding Protein 4 by Interleukin-6 via Peroxisome Proliferator-Activated Receptor α and CCAAT/Enhancer Binding Proteins. Mol. Cell. Endocrinol. 2020, 505, 110702. [Google Scholar] [CrossRef] [PubMed]
- Tepasse, P.-R.; Vollenberg, R.; Fobker, M.; Kabar, I.; Schmidt, H.; Meier, J.A.; Nowacki, T.; Hüsing-Kabar, A. Vitamin A Plasma Levels in COVID-19 Patients: A Prospective Multicenter Study and Hypothesis. Nutrients 2021, 13, 2173. [Google Scholar] [CrossRef]
- Vollenberg, R.; Tepasse, P.-R.; Fobker, M.; Hüsing-Kabar, A. Significantly Reduced Retinol Binding Protein 4 (RBP4) Levels in Critically Ill COVID-19 Patients. Nutrients 2022, 14, 2007. [Google Scholar] [CrossRef] [PubMed]
- Kirtana, J.; Kumar, A.; Kumar, S.; Singh, A.; Shankar, S.; Sharma, A.; Kumar, A.; Kaur, R.; Khan, M.; Ranjan, P.; et al. Mild COVID-19 Infection-Predicting Symptomatic Phase and Outcome: A Study from AIIMS, New Delhi. J. Fam. Med. Prim. Care 2020, 9, 5360. [Google Scholar] [CrossRef]
- Liu, J.; Li, S.; Liu, J.; Liang, B.; Wang, X.; Wang, H.; Li, W.; Tong, Q.; Yi, J.; Zhao, L.; et al. Longitudinal Characteristics of Lymphocyte Responses and Cytokine Profiles in the Peripheral Blood of SARS-CoV-2 Infected Patients. eBioMedicine 2020, 55, 102763. [Google Scholar] [CrossRef]
- Abdelkader, A.; Wahba, A.A.; El-tonsy, M.; Zewail, A.A.; Shams Eldin, M. Recurrent Respiratory Infections and Vitamin A Levels: A Link? It Is Cross-Sectional. Medicine 2022, 101, e30108. [Google Scholar] [CrossRef]
- Almoosawi, S.; Palla, L. Association between Vitamin Intake and Respiratory Complaints in Adults from the UK National Diet and Nutrition Survey Years 1–8. BMJ Nutr. Prev. Health 2020, 3, 403–408. [Google Scholar] [CrossRef]
- Ross, A.C.; Chen, Q.; Ma, Y. Vitamin A and Retinoic Acid in the Regulation of B-Cell Development and Antibody Production. In Vitamins & Hormones; Elsevier: Amsterdam, The Netherlands, 2011; Volume 86, pp. 103–126. ISBN 978-0-12-386960-9. [Google Scholar]
- Surman, S.L.; Jones, B.G.; Rudraraju, R.; Sealy, R.E.; Hurwitz, J.L. Intranasal Administration of Retinyl Palmitate with a Respiratory Virus Vaccine Corrects Impaired Mucosal IgA Response in the Vitamin A-Deficient Host. Clin. Vaccine Immunol. 2014, 21, 598–601. [Google Scholar] [CrossRef]
- Rudraraju, R.; Jones, B.G.; Surman, S.L.; Sealy, R.E.; Thomas, P.G.; Hurwitz, J.L. Respiratory Tract Epithelial Cells Express Retinaldehyde Dehydrogenase ALDH1A and Enhance IgA Production by Stimulated B Cells in the Presence of Vitamin A. PLoS ONE 2014, 9, e86554. [Google Scholar] [CrossRef]
- Bemark, M.; Angeletti, D. Know Your Enemy or Find Your Friend?—Induction of IgA at Mucosal Surfaces. Immunol. Rev. 2021, 303, 83–102. [Google Scholar] [CrossRef] [PubMed]
- Grewal, J.S.; Pilgrim, M.J.; Grewal, S.; Kasman, L.; Werner, P.; Bruorton, M.E.; London, S.D.; London, L. Salivary Glands Act as Mucosal Inductive Sites via the Formation of Ectopic Germinal Centers after Site-restricted MCMV Infection. FASEB J. 2011, 25, 1680–1696. [Google Scholar] [CrossRef] [PubMed]
- Jackson, D.E.; Lally, E.T.; Nakamura, M.C.; Montgomery, P.C. Migration of IgA-Bearing Lymphocytes into Salivary Glands. Cell. Immunol. 1981, 63, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Czerkinsky, C.; Svennerholm, A.M.; Quiding, M.; Jonsson, R.; Holmgren, J. Antibody-Producing Cells in Peripheral Blood and Salivary Glands after Oral Cholera Vaccination of Humans. Infect. Immun. 1991, 59, 996–1001. [Google Scholar] [CrossRef] [PubMed]
- Kunkel, E.J.; Butcher, E.C. Plasma-Cell Homing. Nat. Rev. Immunol. 2003, 3, 822–829. [Google Scholar] [CrossRef] [PubMed]
- Sheikh-Mohamed, S.; Isho, B.; Chao, G.Y.C.; Zuo, M.; Cohen, C.; Lustig, Y.; Nahass, G.R.; Salomon-Shulman, R.E.; Blacker, G.; Fazel-Zarandi, M.; et al. Systemic and Mucosal IgA Responses Are Variably Induced in Response to SARS-CoV-2 mRNA Vaccination and Are Associated with Protection against Subsequent Infection. Mucosal Immunol. 2022, 15, 799–808. [Google Scholar] [CrossRef]
- Su, F.; Patel, G.B.; Hu, S.; Chen, W. Induction of Mucosal Immunity through Systemic Immunization: Phantom or Reality? Hum. Vaccines Immunother. 2016, 12, 1070–1079. [Google Scholar] [CrossRef]
- Midha, I.K.; Kumar, N.; Kumar, A.; Madan, T. Mega Doses of Retinol: A Possible Immunomodulation in COVID-19 Illness in Resource-limited Settings. Rev. Med. Virol. 2021, 31, 1–14. [Google Scholar] [CrossRef]
- Ma, Y.; Ross, A.C. The Anti-Tetanus Immune Response of Neonatal Mice Is Augmented by Retinoic Acid Combined with Polyriboinosinic:Polyribocytidylic Acid. Proc. Natl. Acad. Sci. USA 2005, 102, 13556–13561. [Google Scholar] [CrossRef]
- Lisulo, M.M.; Kapulu, M.C.; Banda, R.; Sinkala, E.; Kayamba, V.; Sianongo, S.; Kelly, P. Adjuvant Potential of Low Dose All- Trans Retinoic Acid during Oral Typhoid Vaccination in Zambian Men. Clin. Exp. Immunol. 2014, 175, 468–475. [Google Scholar] [CrossRef]
- Said, D.E.; Amer, E.I.; Sheta, E.; Makled, S.; Arafa, F.M.; Diab, H.E. Nano-Encapsulated Antioxidant: Retinoic Acid as a Natural Mucosal Adjuvant for Intranasal Immunization against Chronic Experimental Toxoplasmosis. Trop. Med. Infect. Dis. 2023, 8, 106. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Variable | N = 39 |
|---|---|
| Age (years) | 35 (29–52) |
| Sex, n (%) | |
| Female | 23 (59) |
| Male | 16 (41) |
| Time since onset of symptoms (days) | 19 (17–20) |
| Vaccine administered, n (%) | |
| Pfizer/BioNTech | 12 (31.6) |
| AstraZeneca/Oxford | 8 (21.1) |
| Sputnik V | 1 (2.6) |
| Sinovac | 2 (5.3) |
| CanSino | 12 (31.6) |
| Moderna | 3 (7.9) |
| Full vaccination schedule, n (%) | 38 (100) |
| Time since last vaccination (days) | 134 (59–194) |
| Symptoms (score) | 14 (5.5–24) |
| Temperature (°C) | 36.4 (36.35–36.6) |
| Respiratory rate (rpm) | 14 (14–17) |
| Oxygen saturation (SpO2) | 96 (95–98) |
| Heart rate (ppm) | 74 (68–81) |
| SBP (mmHg) | 115 (110–142) |
| DBP (mmHg) | 79 (73–86) |
| AST/GOT (U/L) | 23.2 (19.39–30.66) |
| ALT/GPT (U/L) | 20.32 (14.45–29.41) |
| hs-CRP (mg/L) | 1.73 (0.39–3.87) |
| IL-6 (pg/mL) | 0.89 (0.44–1.21) |
| TGF-β1 (ng/mL) | 19.75 (15.75–25.52) |
| RBP4 (μg/mL) | 26.77 (19.61–37.57) |
| RBP4 classification, n (%) | |
| Deficiency | 2 (5.1) |
| Insufficiency | 12 (30.8) |
| Sufficiency | 25 (64.1) |
| Variable | N = 39 |
|---|---|
| Lipids (g) | 73.48 (58.89–84.29) |
| Vitamin A (μg RAE) | 511.36 (323.19–677.9) |
| Vitamin D (μg) | 3.42 (1.72–6.01) |
| Vitamin E (mg) | 3.6 (3.01–5.71) |
| Vitamin K (μg) | 25.25 (18.59–45.65) |
| Magnesium (mg) | 275.32 (217.9–350.11) |
| Zinc (mg) | 9.71 (7.75–11.41) |
| Retinol (μg) | 252.28 (172.09–437.56) |
| Carotenoids (μg) | 111.61 (39.98–475.56) |
| β-carotene (μg) | 93.81 (38.21–383.01) |
| α-carotene (μg) | 5.94 (1.91–43.09) |
| β-cryptoxanthin (μg) | 5.42 (0.98–17.84) |
| Variable | N = 39 |
|---|---|
| SIgA (μg/mL) | 152.4 (120.72–210.8) |
| IgA (μg/mL) | 734.96 (613.32–938.63) |
| IgM (μg/mL) | 1065.5 (696.15–1685.1) |
| IgG1 (μg/mL) | 5574.4 (4780.85–6696.25) |
| IgG2 (μg/mL) | 1262.2 (907.87–1586.45) |
| IgG3 (μg/mL) | 659.45 (514.89–1130.55) |
| IgG4 (μg/mL) | 436.26 (191.53–702.08) |
| RBD NAb (%) | 96.8 (96.7–96.88) |
| Variable | Cut-Off | +LR (CI 95%) | Sensitivity (CI 95%) | Specificity (CI 95%) | p-Value |
|---|---|---|---|---|---|
| Vitamin A | >476 μg RAE | 2.22 (1.16–4.24) | 77.8 (54.8–91.0) | 65 (43.3–81.9) | 0.031 |
| RBP4 | >25.05 μg/mL | 2.26 (1.19–4.28) | 78.9 (56.7–91.5) | 65 (43.3–81.9) | 0.005 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Turrubiates-Hernández, F.J.; García-Arellano, S.; Herrera-Jiménez, L.E.; González-Estevez, G.; Márquez-Sandoval, F.; Padilla-Bórquez, D.L.; Ceja-Gálvez, H.R.; Vega-Magaña, N.; Nicoletti, F.; Muñoz-Ríos, G.; et al. Vitamin A Positively Correlates with Secretory Immunoglobulin A: A Cross-Sectional Study in Omicron COVID-19 Outpatients. J. Clin. Med. 2024, 13, 1538. https://doi.org/10.3390/jcm13061538
Turrubiates-Hernández FJ, García-Arellano S, Herrera-Jiménez LE, González-Estevez G, Márquez-Sandoval F, Padilla-Bórquez DL, Ceja-Gálvez HR, Vega-Magaña N, Nicoletti F, Muñoz-Ríos G, et al. Vitamin A Positively Correlates with Secretory Immunoglobulin A: A Cross-Sectional Study in Omicron COVID-19 Outpatients. Journal of Clinical Medicine. 2024; 13(6):1538. https://doi.org/10.3390/jcm13061538
Chicago/Turabian StyleTurrubiates-Hernández, Francisco Javier, Samuel García-Arellano, Laura Elena Herrera-Jiménez, Guillermo González-Estevez, Fabiola Márquez-Sandoval, Diana Lourdes Padilla-Bórquez, Hazael Ramiro Ceja-Gálvez, Natali Vega-Magaña, Ferdinando Nicoletti, Guillermina Muñoz-Ríos, and et al. 2024. "Vitamin A Positively Correlates with Secretory Immunoglobulin A: A Cross-Sectional Study in Omicron COVID-19 Outpatients" Journal of Clinical Medicine 13, no. 6: 1538. https://doi.org/10.3390/jcm13061538
APA StyleTurrubiates-Hernández, F. J., García-Arellano, S., Herrera-Jiménez, L. E., González-Estevez, G., Márquez-Sandoval, F., Padilla-Bórquez, D. L., Ceja-Gálvez, H. R., Vega-Magaña, N., Nicoletti, F., Muñoz-Ríos, G., & Muñoz-Valle, J. F. (2024). Vitamin A Positively Correlates with Secretory Immunoglobulin A: A Cross-Sectional Study in Omicron COVID-19 Outpatients. Journal of Clinical Medicine, 13(6), 1538. https://doi.org/10.3390/jcm13061538