
Increased Soluble Interleukin 6 Receptors in Fabry Disease

 ,
,  ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Blood and Urine Samples
2.2. Isolation and Characterization of Exosomes from Urine and Blood
2.3. Interleukin 6 and IL-6 Receptors Quantification in Blood and in Exosomes
2.4. Statistical Analysis
3. Results
3.1. Demographic and Clinical Data of FD Patients
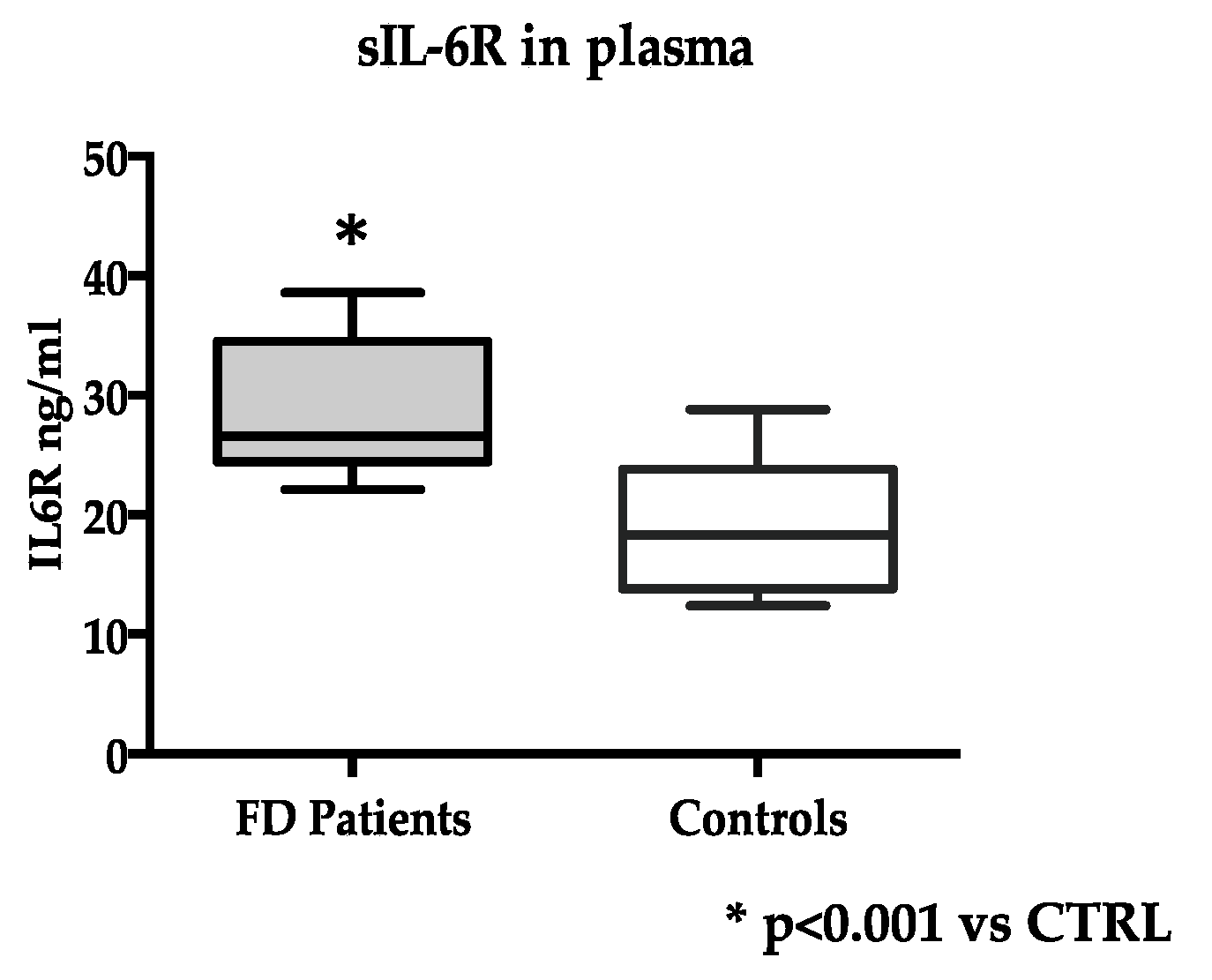
3.2. IL-6 and IL-6R Levels in FD and Healthy Subjects
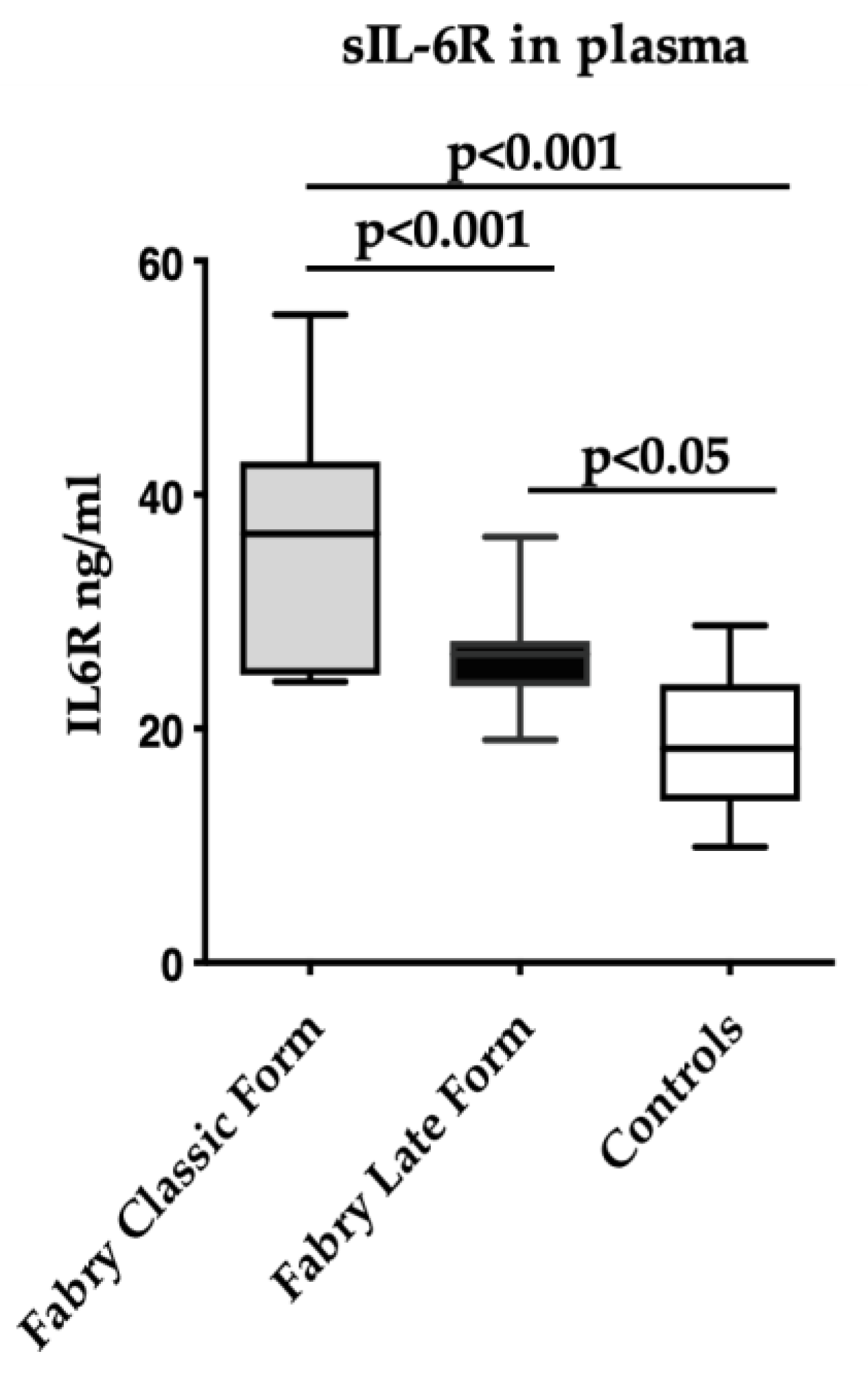
3.3. Soluble IL-6R Levels and FD Patients’ Characteristics
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tøndel, C.; Thurberg, B.L.; DasMahapatra, P.; Lyn, N.; Maski, M.; Batista, J.L.; George, K.; Patel, H.; Hariri, A. Clinical Relevance of Globotriaosylceramide Accumulation in Fabry Disease and the Effect of Agalsidase Beta in Affected Tissues. Mol. Genet. Metab. 2022, 137, 328–341. [Google Scholar] [CrossRef] [PubMed]
- Desnick, R.J.; Brady, R.; Barranger, J.; Collins, A.J.; Germain, D.P.; Goldman, M.; Grabowski, G.; Packman, S.; Wilcox, W.R. Fabry Disease, an under-Recognized Multisystemic Disorder: Expert Recommendations for Diagnosis, Management, and Enzyme Replacement Therapy. Ann. Intern. Med. 2003, 138, 338–346. [Google Scholar] [CrossRef] [PubMed]
- Bertoldi, G.; Carraro, G.; Ravarotto, V.; Di Vico, V.; Baldini Anastasio, P.; Vitturi, N.; Francini, F.; Stefanelli, L.F.; Calò, L.A. The Effect of Green Tea as an Adjuvant to Enzyme Replacement Therapy on Oxidative Stress in Fabry Disease: A Pilot Study. Front. Nutr. 2022, 9, 924710. [Google Scholar] [CrossRef] [PubMed]
- Ravarotto, V.; Carraro, G.; Pagnin, E.; Bertoldi, G.; Simioni, F.; Maiolino, G.; Martinato, M.; Landini, L.; Davis, P.A.; Calò, L.A. Oxidative Stress and the Altered Reaction to It in Fabry Disease: A Possible Target for Cardiovascular-Renal Remodeling? PLoS ONE 2018, 13, e0204618. [Google Scholar] [CrossRef] [PubMed]
- Simoncini, C.; Torri, S.; Montano, V.; Chico, L.; Gruosso, F.; Tuttolomondo, A.; Pinto, A.; Simonetta, I.; Cianci, V.; Salviati, A.; et al. Oxidative Stress Biomarkers in Fabry Disease: Is There a Room for Them? J. Neurol. 2020, 267, 3741–3752. [Google Scholar] [CrossRef] [PubMed]
- Ravarotto, V.; Simioni, F.; Carraro, G.; Bertoldi, G.; Pagnin, E.; Calò, L.A. Oxidative Stress and Cardiovascular-Renal Damage in Fabry Disease: Is There Room for a Pathophysiological Involvement? J. Clin. Med. 2018, 7, 409. [Google Scholar] [CrossRef] [PubMed]
- Weidemann, F.; Sanchez-Niño, M.D.; Politei, J.; Oliveira, J.-P.; Wanner, C.; Warnock, D.G.; Ortiz, A. Fibrosis: A Key Feature of Fabry Disease with Potential Therapeutic Implications. Orphanet J. Rare Dis. 2013, 8, 116. [Google Scholar] [CrossRef]
- Yogasundaram, H.; Nikhanj, A.; Putko, B.N.; Boutin, M.; Jain-Ghai, S.; Khan, A.; Auray-Blais, C.; West, M.L.; Oudit, G.Y. Elevated Inflammatory Plasma Biomarkers in Patients with Fabry Disease: A Critical Link to Heart Failure with Preserved Ejection Fraction. J. Am. Heart Assoc. 2018, 7, e009098. [Google Scholar] [CrossRef]
- Vujkovac, A.C.; Vujkovac, B.; Novaković, S.; Števanec, M.; Šabovič, M. Characteristics of Vascular Phenotype in Fabry Patients. Angiology 2021, 72, 426–433. [Google Scholar] [CrossRef]
- Biancini, G.B.; Vanzin, C.S.; Rodrigues, D.B.; Deon, M.; Ribas, G.S.; Barschak, A.G.; Manfredini, V.; Netto, C.B.O.; Jardim, L.B.; Giugliani, R.; et al. Globotriaosylceramide Is Correlated with Oxidative Stress and Inflammation in Fabry Patients Treated with Enzyme Replacement Therapy. Biochim. Biophys. Acta Mol. Basis Dis. 2012, 1822, 226–232. [Google Scholar] [CrossRef]
- Chen, K.H.; Chien, Y.; Wang, K.L.; Leu, H.B.; Hsiao, C.Y.; Lai, Y.H.; Wang, C.Y.; Chang, Y.L.; Lin, S.J.; Niu, D.M.; et al. Evaluation of Proinflammatory Prognostic Biomarkers for Fabry Cardiomyopathy with Enzyme Replacement Therapy. Can. J. Cardiol. 2016, 32, 1221.e1–1221.e9. [Google Scholar] [CrossRef] [PubMed]
- Rosa Neto, N.S.; de Barros Bento, J.C.; de Falco Caparbo, V.; Pereira, R.M.R. Increased Serum Interleukin-6 and Tumor Necrosis Factor Alpha Levels in Fabry Disease: Correlation with Disease Burden. Clinics 2021, 76, e2643. [Google Scholar] [CrossRef]
- Schumertl, T.; Lokau, J.; Rose-John, S.; Garbers, C. Function and Proteolytic Generation of the Soluble Interleukin-6 Receptor in Health and Disease. Biochim. Biophys. Acta Mol. Cell Res. 2022, 1869, 119143. [Google Scholar] [CrossRef]
- Rose-John, S.; Jenkins, B.J.; Garbers, C.; Moll, J.M.; Scheller, J. Targeting IL-6 Trans-Signalling: Past, Present and Future Prospects. Nat. Rev. Immunol. 2023, 23, 666–681. [Google Scholar] [CrossRef]
- Lust, J.A.; Donovan, K.A.; Kline, M.P.; Greipp, P.R.; Kyle, R.A.; Maihle, N.J. Isolation of an MRNA Encoding a Soluble Form of the Human Interleukin-6 Receptor. Cytokine 1992, 4, 96–100. [Google Scholar] [CrossRef]
- Moss, M.L.; Jin, S.L.C.; Milla, M.E.; Bickett, D.M.; Burkhart, W.; Carter, H.L.; Chen, W.J.; Clay, W.C.; Didsbury, J.R.; Hassler, D.; et al. Erratum: Cloning of a Disintegrin Metalloproteinase That Processes Precursor Tumour-Necrosis Factor-α (Nature (1997) 385 (733–736)). Nature 1997, 386, 738. [Google Scholar] [CrossRef]
- Black, R.A.; Rauch, C.T.; Kozlosky, C.J.; Peschon, J.J.; Slack, J.L.; Wolfson, M.F.; Castner, B.J.; Stocking, K.L.; Reddy, P.; Srinivasan, S.; et al. A Metalloproteinase Disintegrin That Releases from Cells. Nature 1997, 385, 729–733. [Google Scholar] [CrossRef]
- Arnold, P.; Lückstädt, W.; Li, W.; Boll, I.; Lokau, J.; Garbers, C.; Lucius, R.; Rose-John, S.; Becker-Pauly, C. Joint Reconstituted Signaling of the IL-6 Receptor via Extracellular Vesicles. Cells 2020, 9, 1307. [Google Scholar] [CrossRef]
- Gheinani, A.H.; Vögeli, M.; Baumgartner, U.; Vassella, E.; Draeger, A.; Burkhard, F.C.; Monastyrskaya, K. Improved Isolation Strategies to Increase the Yield and Purity of Human Urinary Exosomes for Biomarker Discovery. Sci. Rep. 2018, 8, 3945. [Google Scholar] [CrossRef]
- Anderson, D.R.; Poterucha, J.T.; Mikuls, T.R.; Duryee, M.J.; Garvin, R.P.; Klassen, L.W.; Shurmur, S.W.; Thiele, G.M. IL-6 and Its Receptors in Coronary Artery Disease and Acute Myocardial Infarction. Cytokine 2013, 62, 395–400. [Google Scholar] [CrossRef]
- Kanda, T.; Inoue, M.; Kotajima, N.; Fujimaki, S.; Hoshino, Y.; Kurabayashi, M.; Kobayashi, I.; Tamura, J. Circulating Interleukin-6 and Interleukin-6 Receptors in Patients with Acute and Recent Myocardial Infarction. Cardiology 2000, 93, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Keul, R.; Heinrich, P.C.; Müller-newen, G.; Muller, K.; Woo, P. A Possible Role for Soluble IL-6 Receptor in the Pathogenesis of Systemic Onset Juvenile Chronic Arthritis. Cytokine 1998, 10, 729–734. [Google Scholar] [CrossRef] [PubMed]
- Lenders, M.; Brand, E. Fabry Disease—A Multisystemic Disease with Gastrointestinal Manifestations. Gut Microbes 2022, 14, 2027852. [Google Scholar] [CrossRef] [PubMed]
- Gugelmo, G.; Vitturi, N.; Francini-Pesenti, F.; Fasan, I.; Lenzini, L.; Valentini, R.; Carraro, G.; Avogaro, A.; Spinella, P. Gastrointestinal Manifestations and Low-FODMAP Protocol in a Cohort of Fabry Disease Adult Patients. Nutrients 2023, 15, 658. [Google Scholar] [CrossRef] [PubMed]
- Germain, D.P.; Benistan, K.; Boutouyrie, P.; Mutschler, C. Osteopenia and Osteoporosis: Previously Unrecognized Manifestations of Fabry Disease. Clin. Genet. 2005, 68, 93–95. [Google Scholar] [PubMed]
- Nose, Y.; Fujii, H.; Goto, S.; Kono, K.; Okamoto, H.; Watanabe, K.; Nishi, S. Investigation of Bone Mineral Density and the Changes by Enzyme Replacement Therapy in Patients with Fabry Disease. Mol. Genet. Metab. 2023, 139, 107634. [Google Scholar] [CrossRef] [PubMed]
- Aliyu, M.; Zohora, F.T.; Anka, A.U.; Ali, K.; Maleknia, S.; Saffarioun, M.; Azizi, G. Interleukin-6 Cytokine: An Overview of the Immune Regulation, Immune Dysregulation, and Therapeutic Approach. Int. Immunopharmacol. 2022, 111, 109130. [Google Scholar] [CrossRef]
- Rose-John, S. Interleukin-6 Family Cytokines. Cold Spring Harb. Perspect. Biol. 2018, 10, a028415. [Google Scholar] [CrossRef]
- Parolo, S.; Tomasoni, D.; Bora, P.; Ramponi, A.; Kaddi, C.; Azer, K.; Domenici, E.; Neves-Zaph, S.; Lombardo, R. Reconstruction of the Cytokine Signaling in Lysosomal Storage Diseases by Literature Mining and Network Analysis. Front. Cell Dev. Biol. 2021, 9, 703489. [Google Scholar] [CrossRef]
- Tebani, A.; Barbey, F.; Dormond, O.; Ducatez, F.; Marret, S.; Nowak, A.; Bekri, S. Deep Next-Generation Proteomics and Network Analysis Reveal Systemic and Tissue-Specific Patterns in Fabry Disease. Transl. Res. 2023, 258, 47–59. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Fabry Disease Patients (n = 20) | |
|---|---|
| Age (years) | 43 ± 16 |
| Sex | 60% females; 40% males |
| GLA mutations (n of subjects): | |
| N215S | 7 |
| W287X | 4 |
| A73V | 3 |
| M290T | 1 |
| R301X | 1 |
| G35R | 1 |
| G212Pfs*18 | 1 |
| Q279K | 1 |
| G360Wfs*15 | 1 |
| Disease form (%) | 30% Classical, 70% Late-onset |
| Therapy (%) | 50% ERT, 20% Chaperonic, 30% none |
| e-GFR CKD EPI (mL/min’/1.73 mq) | 87.0 ± 27.6 |
| Proteinuria (>0.15 g/24 h) | 45% yes, 55% no |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lenzini, L.; Iori, E.; Vettore, M.; Gugelmo, G.; Radu, C.; Padoan, A.; Carraro, G.; Simioni, P.; Calò, L.; Avogaro, A.; et al. Increased Soluble Interleukin 6 Receptors in Fabry Disease. J. Clin. Med. 2024, 13, 218. https://doi.org/10.3390/jcm13010218
Lenzini L, Iori E, Vettore M, Gugelmo G, Radu C, Padoan A, Carraro G, Simioni P, Calò L, Avogaro A, et al. Increased Soluble Interleukin 6 Receptors in Fabry Disease. Journal of Clinical Medicine. 2024; 13(1):218. https://doi.org/10.3390/jcm13010218
Chicago/Turabian StyleLenzini, Livia, Elisabetta Iori, Monica Vettore, Giorgia Gugelmo, Claudia Radu, Andrea Padoan, Gianni Carraro, Paolo Simioni, Lorenzo Calò, Angelo Avogaro, and et al. 2024. "Increased Soluble Interleukin 6 Receptors in Fabry Disease" Journal of Clinical Medicine 13, no. 1: 218. https://doi.org/10.3390/jcm13010218
APA StyleLenzini, L., Iori, E., Vettore, M., Gugelmo, G., Radu, C., Padoan, A., Carraro, G., Simioni, P., Calò, L., Avogaro, A., Rossi, G. P., & Vitturi, N. (2024). Increased Soluble Interleukin 6 Receptors in Fabry Disease. Journal of Clinical Medicine, 13(1), 218. https://doi.org/10.3390/jcm13010218