
Pathogenesis of Chronic Kidney Disease Is Closely Bound up with Alzheimer’s Disease, Especially via the Renin-Angiotensin System

Abstract
:1. Introduction
2. Pathophysiology of Chronic Kidney Disease
3. Alzheimer’s Disease Pathogenesis
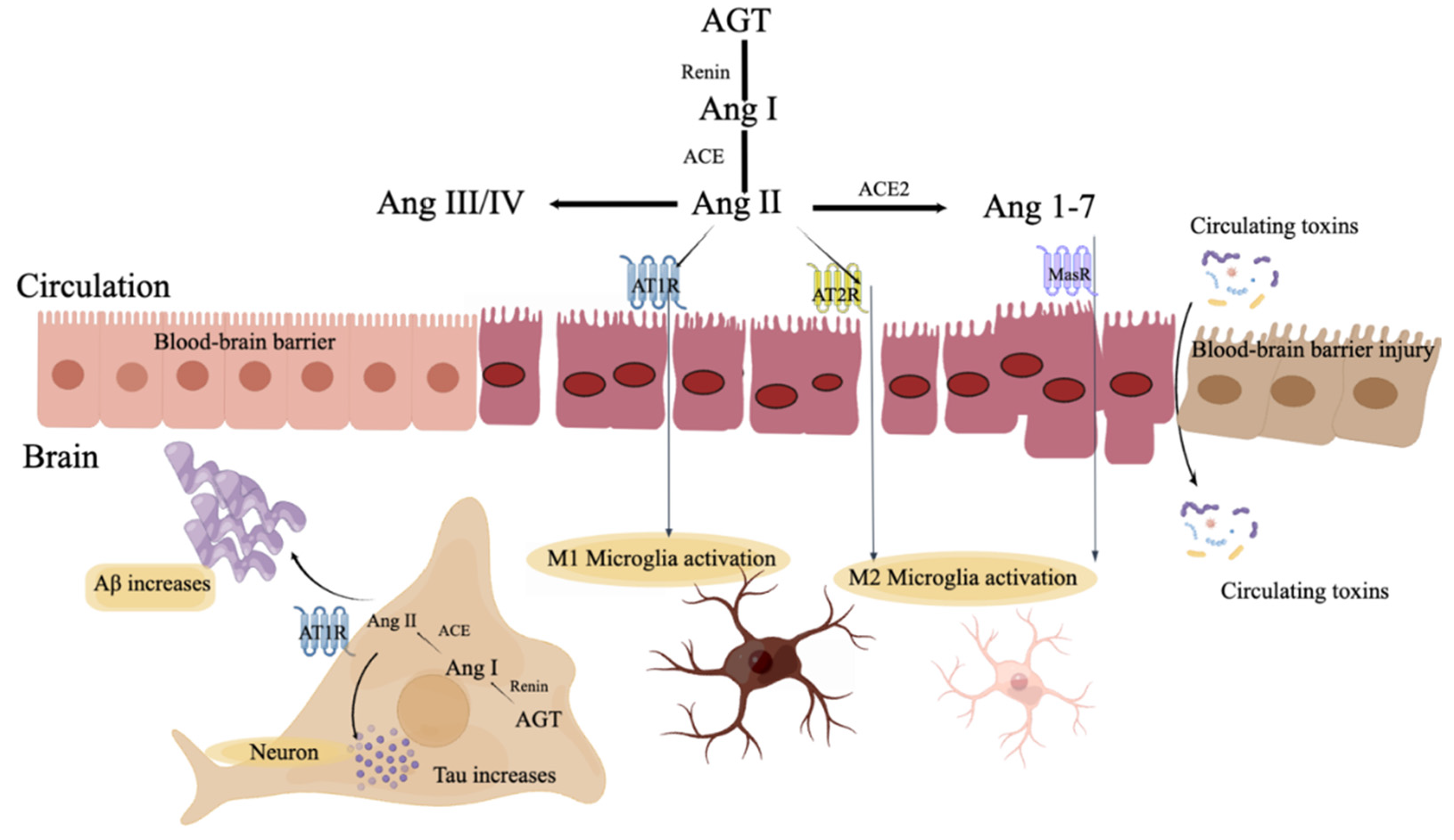
4. Renin-Angiotensin System
4.1. RAS in the Systemic Circulation
4.2. RAS in the Brain
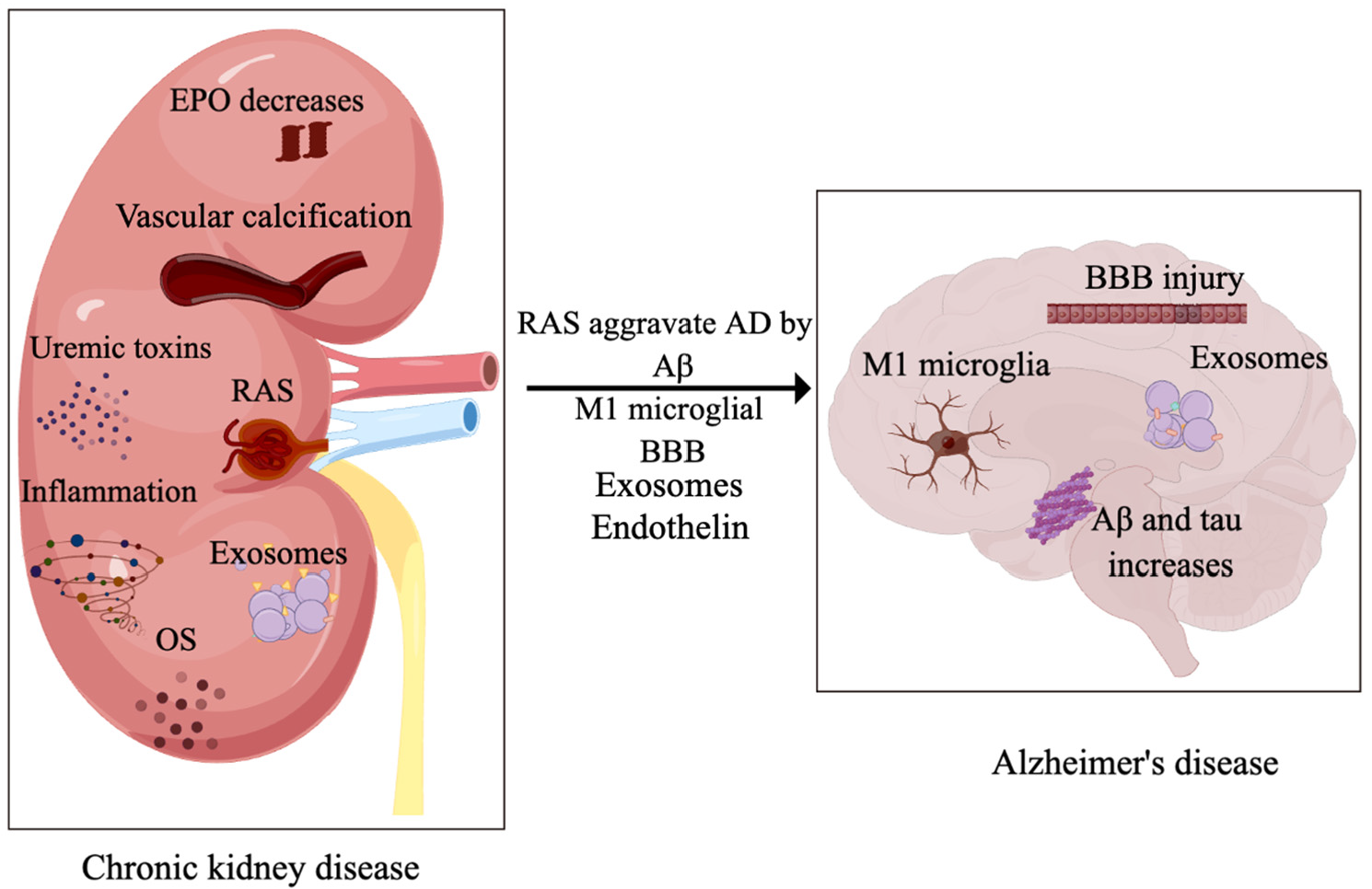
5. Association between CKD and AD
5.1. RAS Linking CKD and AD
5.2. RAS and Aβ and Tau in AD
5.3. RAS and Microglia
5.4. RAS and the Blood–Brain Barrier (BBB)
6. Uremic Toxins
7. Erythropoietin
8. Extracellular Vehicles
9. Endothelin
10. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| CKD | chronic kidney disease |
| AD | Alzheimer’s disease |
| Aβ | β-amyloid |
| NFT | Neurofibrillary tangle |
| RAS | Renin-angiotensin system |
| ACE 1, 2 | Angiotensin-converting enzyme 1, 2 |
| ACEI | ACE inhibitor |
| ApoE4 | Epsilon 4 allele of the apolipoprotein E gene |
| APP | Amyloid precursor protein |
| SorLA | Sorting protein-related receptor |
| GFR | Glomerular filtration rate |
| EPO | Erythropoietin |
| EV | Extracellular vehicle |
| ROS | Reactive oxygen species |
| Ang I, II, III, IV | Angiotensin I, II, III, IV |
| AT1/2/4R | Ang II receptor type 1, 2, 4 |
| AGT | Angiotensinogen |
| AVP | Arginine vasopressin |
| CNS | Central nervous system |
| Atp6ap2 | ATPase H+ transporting accessory protein 2 |
| AGTR | AGT receptor |
| MasR | Mas receptor |
| ARB | Angiotensin-II-receptor blocker |
| TLR4 | Toll-like receptor 4 |
| iNOS | Inducible nitric oxide synthase |
| CXCL12 | C-X-C motif chemokine ligand 12 |
| IL-1β, 4, 6, 10 | Interleukin 1β, 4, 6, 10 |
| BBB | Blood-brain barrier |
| TNF-α | Tumor necrosis factor α |
| EPOR | EPO receptor |
| PKB | Protein kinase B |
| MCP-1 | Monocyte chemoattractant protein-1 |
References
- Mayeux, R.; Stern, Y. Epidemiology of Alzheimer Disease. Cold Spring Harb. Perspect. Med. 2012, 2, a006239. [Google Scholar] [CrossRef] [PubMed]
- Fogari, R.; Zoppi, A. Effect of Antihypertensive Agents on Quality of Life in the Elderly. Drugs Aging 2004, 21, 377–393. [Google Scholar] [CrossRef] [PubMed]
- DeTure, M.A.; Dickson, D.W. The Neuropathological Diagnosis of Alzheimer’s Disease. Mol. Neurodegener. 2019, 14, 32. [Google Scholar] [CrossRef]
- Kopp, J.B. Global Glomerulosclerosis in Primary Nephrotic Syndrome: Including Age as a Variable to Predict Renal Outcomes. Kidney Int. 2018, 93, 1043–1044. [Google Scholar] [CrossRef]
- Huang, W.; Qiu, C.; von Strauss, E.; Winblad, B.; Fratiglioni, L. Apoe Genotype, Family History of Dementia, and Alzheimer Disease Risk: A 6-Year Follow-up Study. Arch. Neurol. 2004, 61, 1930–1934. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, Y.J.; Zhang, M.; Xu, Z.Q.; Gao, C.Y.; Fang, C.Q.; Yan, J.C.; Zhou, H.D.; Group Chongqing Ageing Study. Vascular Risk Factors Promote Conversion from Mild Cognitive Impairment to Alzheimer Disease. Neurology 2011, 76, 1485–1491. [Google Scholar] [CrossRef]
- Kivipelto, M.; Ngandu, T.; Fratiglioni, L.; Viitanen, M.; Kåreholt, I.; Winblad, B.; Helkala, E.L.; Tuomilehto, J.; Soininen, H.; Nissinen, A. Obesity and Vascular Risk Factors at Midlife and the Risk of Dementia and Alzheimer Disease. Arch. Neurol. 2005, 62, 1556–1560. [Google Scholar] [CrossRef]
- Forbes, J.M.; Cooper, M.E. Mechanisms of Diabetic Complications. Physiol. Rev. 2013, 93, 137–188. [Google Scholar] [CrossRef]
- Stanciu, G.D.; Bild, V.; Ababei, D.C.; Rusu, R.N.; Cobzaru, A.; Paduraru, L.; Bulea, D. Link between Diabetes and Alzheimer’s Disease Due to the Shared Amyloid Aggregation and Deposition Involving Both Neurodegenerative Changes and Neurovascular Damages. J. Clin. Med. 2020, 9, 1713. [Google Scholar] [CrossRef]
- Wan, J.; Wang, S.; Haynes, K.; Denburg, M.R.; Shin, D.B.; Gelfand, J.M. Risk of Moderate to Advanced Kidney Disease in Patients with Psoriasis: Population Based Cohort Study. BMJ 2013, 347, f5961. [Google Scholar] [CrossRef]
- Willnow, T.E.; Andersen, O.M. Sorting Receptor Sorla—A Trafficking Path to Avoid Alzheimer Disease. J. Cell Sci. 2013, 126, 2751–2760. [Google Scholar] [CrossRef]
- Scherzer, C.R.; Offe, K.; Gearing, M.; Rees, H.D.; Fang, G.; Heilman, C.J.; Schaller, C.; Bujo, H.; Levey, A.I.; Lah, J.J. Loss of Apolipoprotein E Receptor Lr11 in Alzheimer Disease. Arch. Neurol. 2004, 61, 1200–1205. [Google Scholar] [CrossRef]
- Nielsen, M.S.; Gustafsen, C.; Madsen, P.; Nyengaard, J.R.; Hermey, G.; Bakke, O.; Mari, M.; Schu, P.; Pohlmann, R.; Dennes, A.; et al. Sorting by the Cytoplasmic Domain of the Amyloid Precursor Protein Binding Receptor Sorla. Mol. Cell. Biol. 2007, 27, 6842–6851. [Google Scholar] [CrossRef]
- Ştefănescu, R.; Stanciu, G.D.; Luca, A.; Caba, I.C.; Tamba, B.I.; Mihai, C.T. Contributions of Mass Spectrometry to the Identification of Low Molecular Weight Molecules Able to Reduce the Toxicity of Amyloid-Beta Peptide to Cell Cultures and Transgenic Mouse Models of Alzheimer’s Disease. Molecules 2019, 24, 1167. [Google Scholar] [CrossRef]
- Stanciu, G.D.; Luca, A.; Rusu, R.N.; Bild, V.; Chiriac, S.I.B.; Solcan, C.; Bild, W.; Ababei, D.C. Alzheimer’s Disease Pharmacotherapy in Relation to Cholinergic System Involvement. Biomolecules 2019, 10, 40. [Google Scholar] [CrossRef]
- Jha, V.; Garcia-Garcia, G.; Iseki, K.; Li, Z.; Naicker, S.; Plattner, B.; Saran, R.; Wang, A.Y.M.; Yang, C.W. Chronic Kidney Disease: Global Dimension and Perspectives. Lancet 2013, 382, 260–272. [Google Scholar] [CrossRef]
- Krishnan, A.V.; Kiernan, M.C. Neurological Complications of Chronic Kidney Disease. Nat. Rev. Neurol. 2009, 5, 542–551. [Google Scholar] [CrossRef]
- Zhang, C.Y.; He, F.F.; Su, H.; Zhang, C.; Meng, X.F. Association between Chronic Kidney Disease and Alzheimer’s Disease: An Update. Metab. Brain Dis. 2020, 35, 883–894. [Google Scholar] [CrossRef]
- Stanciu, G.D.; Ababei, D.C.; Bild, V.; Bild, W.; Paduraru, L.; Gutu, M.M.; Tamba, B.I. Renal Contributions in the Pathophysiology and Neuropathological Substrates Shared by Chronic Kidney Disease and Alzheimer’s Disease. Brain Sci. 2020, 10, 563. [Google Scholar] [CrossRef]
- Nakagawa, T.; Hasegawa, Y.; Uekawa, K.; Kim-Mitsuyama, S. Chronic Kidney Disease Accelerates Cognitive Impairment in a Mouse Model of Alzheimer’s Disease, through Angiotensin Ii. Exp. Gerontol. 2017, 87, 108–112. [Google Scholar] [CrossRef]
- Xue, L.; Lou, Y.; Feng, X.; Wang, C.; Ran, Z.; Zhang, X. Prevalence of Chronic Kidney Disease and Associated Factors among the Chinese Population in Taian, China. BMC Nephrol. 2014, 15, 205. [Google Scholar] [CrossRef] [PubMed]
- Loutradis, C.; Price, A.; Ferro, C.J.; Sarafidis, P. Renin-Angiotensin System Blockade in Patients with Chronic Kidney Disease: Benefits, Problems in Everyday Clinical Use, and Open Questions for Advanced Renal Dysfunction. J. Hum. Hypertens. 2021, 35, 499–509. [Google Scholar] [CrossRef]
- Walther, C.P.; Winkelmayer, W.C.; Richardson, P.A.; Virani, S.S.; Navaneethan, S.D. Renin-Angiotensin System Blocker Discontinuation and Adverse Outcomes in Chronic Kidney Disease. Nephrol. Dial. Transplant. 2021, 36, 1893–1899. [Google Scholar] [CrossRef] [PubMed]
- Natale, G.; Calabrese, V.; Marino, G.; Campanelli, F.; Urciuolo, F.; de Iure, A.; Ghiglieri, V.; Calabresi, P.; Bossola, M.; Picconi, B. Effects of Uremic Toxins on Hippocampal Synaptic Transmission: Implication for Neurodegeneration in Chronic Kidney Disease. Cell Death Discov. 2021, 7, 295. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Bowe, B.; Mokdad, A.H.; Xian, H.; Yan, Y.; Li, T.; Maddukuri, G.; Tsai, C.Y.; Floyd, T.; Al-Aly, Z. Analysis of the Global Burden of Disease Study Highlights the Global, Regional, and National Trends of Chronic Kidney Disease Epidemiology from 1990 to 2016. Kidney Int. 2018, 94, 567–581. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.K.; Winocour, P.; Farrington, K. Oxidative Stress in Early Diabetic Nephropathy: Fueling the Fire. Nat. Rev. Endocrinol. 2011, 7, 176–184. [Google Scholar] [CrossRef]
- Kim, H.J.; Vaziri, N.D. Contribution of Impaired Nrf2-Keap1 Pathway to Oxidative Stress and Inflammation in Chronic Renal Failure. Am. J. Physiol. Renal. Physiol. 2010, 298, F662–F671. [Google Scholar] [CrossRef]
- Noonan, M.L.; Clinkenbeard, E.L.; Ni, P.; Swallow, E.A.; Tippen, S.P.; Agoro, R.; Allen, M.R.; White, K.E. Erythropoietin and a Hypoxia-Inducible Factor Prolyl Hydroxylase Inhibitor (Hif-Phdi) Lowers Fgf23 in a Model of Chronic Kidney Disease (Ckd). Physiol. Rep. 2020, 8, e14434. [Google Scholar] [CrossRef]
- Noonan, M.L.; Ni, P.; Agoro, R.; Sacks, S.A.; Swallow, E.A.; Wheeler, J.A.; Clinkenbeard, E.L.; Capitano, M.L.; Prideaux, M.; Atkins, G.J.; et al. The Hif-Phi Bay 85-3934 (Molidustat) Improves Anemia and Is Associated with Reduced Levels of Circulating Fgf23 in a Ckd Mouse Model. J. Bone Miner. Res. 2021, 36, 1117–1130. [Google Scholar] [CrossRef]
- Pieniazek, A.; Bernasinska-Slomczewska, J.; Gwozdzinski, L. Uremic Toxins and Their Relation with Oxidative Stress Induced in Patients with Ckd. Int. J. Mol. Sci. 2021, 22, 6196. [Google Scholar] [CrossRef]
- Shaykh, M.; Pegoraro, A.A.; Mo, W.; Arruda, J.A.L.; Dunea, G.; Singh, A.K. Carbamylated Proteins Activate Glomerular Mesangial Cells and Stimulate Collagen Deposition. J. Lab. Clin. Med. 1999, 133, 302–308. [Google Scholar] [CrossRef]
- Pieniazek, A.; Brzeszczynska, J.; Kruszynska, I.; Gwozdzinski, K. Investigation of Albumin Properties in Patients with Chronic Renal Failure. Free Radic. Res. 2009, 43, 1008–1018. [Google Scholar] [CrossRef]
- Jaisson, S.; Pietrement, C.; Gillery, P. Carbamylation-Derived Products: Bioactive Compounds and Potential Biomarkers in Chronic Renal Failure and Atherosclerosis. Clin. Chem. 2011, 57, 1499–1505. [Google Scholar] [CrossRef]
- Koppe, L.; Nyam, E.; Vivot, K.; Fox, J.E.M.; Dai, X.Q.; Nguyen, B.N.; Trudel, D.; Attané, C.; Moullé, V.S.; MacDonald, P.E.; et al. Urea Impairs Beta Cell Glycolysis and Insulin Secretion in Chronic Kidney Disease. J. Clin. Investig. 2016, 126, 3598–3612. [Google Scholar] [CrossRef]
- van Gelder, M.K.; Middel, I.R.; Vernooij, R.W.; Bots, M.L.; Verhaar, M.C.; Masereeuw, R.; Grooteman, M.P.; Nubé, M.J.; van den Dorpel, M.A.; Blankestijn, P.J.; et al. Protein-Bound Uremic Toxins in Hemodialysis Patients Relate to Residual Kidney Function, Are Not Influenced by Convective Transport, and Do Not Relate to Outcome. Toxins 2020, 12, 234. [Google Scholar] [CrossRef]
- Yaker, L.; Kamel, S.; Ausseil, J.; Boullier, A. Effects of Chronic Kidney Disease and Uremic Toxins on Extracellular Vesicle Biology. Toxins 2020, 12, 811. [Google Scholar] [CrossRef]
- Chao, C.T.; Lin, S.H. Uremic Toxins and Frailty in Patients with Chronic Kidney Disease: A Molecular Insight. Int. J. Mol. Sci. 2021, 22, 6270. [Google Scholar] [CrossRef]
- Lim, Y.J.; Sidor, N.A.; Tonial, N.C.; Che, A.; Urquhart, B.L. Uremic Toxins in the Progression of Chronic Kidney Disease and Cardiovascular Disease: Mechanisms and Therapeutic Targets. Toxins 2021, 13, 142. [Google Scholar] [CrossRef]
- Wojtaszek, E.; Oldakowska-Jedynak, U.; Kwiatkowska, M.; Glogowski, T.; Malyszko, J. Uremic Toxins, Oxidative Stress, Atherosclerosis in Chronic Kidney Disease, and Kidney Transplantation. Oxid. Med. Cell. Longev. 2021, 2021, 6651367. [Google Scholar] [CrossRef]
- Tamaki, N. Ischemia and Inflammation on Chronic Kidney Disease. J. Nucl. Cardiol. 2019, 26, 441–442. [Google Scholar] [CrossRef]
- Pereira-Balbino, K.; de-Paula-Jorge, M.; Queiroz-Ribeiro, A.; Stampini-Duarte-Martino, H. Modulation of Intestinal Microbiota, Control of Nitrogen Products and Inflammation by Pre/Probiotics in Chronic Kidney Disease: A Systematic Review. Nutr. Hosp. 2018, 35, 722–730. [Google Scholar]
- Mihai, S.; Codrici, E.; Popescu, I.D.; Enciu, A.M.; Albulescu, L.; Necula, L.G.; Mambet, C.; Anton, G.; Tanase, C. Inflammation-Related Mechanisms in Chronic Kidney Disease Prediction, Progression, and Outcome. J. Immunol. Res. 2018, 2018, 2180373. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Ricart, M.; Torramade-Moix, S.; Pascual, G.; Palomo, M.; Moreno-Castaño, A.B.; Martinez-Sanchez, J.; Vera, M.; Cases, A.; Escolar, G. Endothelial Damage, Inflammation and Immunity in Chronic Kidney Disease. Toxins 2020, 12, 361. [Google Scholar] [CrossRef] [PubMed]
- Ebert, T.; Pawelzik, S.C.; Witasp, A.; Arefin, S.; Hobson, S.; Kublickiene, K.; Shiels, P.G.; Bäck, M.; Stenvinkel, P. Inflammation and Premature Ageing in Chronic Kidney Disease. Toxins 2020, 12, 227. [Google Scholar] [CrossRef]
- Mazzaferro, S.; De Martini, N.; Rotondi, S.; Tartaglione, L.; Ureña-Torres, P.; Bover, J.; Pasquali, M.; Era-Edta Working Group on CKD-MBD. Bone, Inflammation and Chronic Kidney Disease. Clin. Chim. Acta 2020, 506, 236–240. [Google Scholar] [CrossRef]
- Brennan, E.; Kantharidis, P.; Cooper, M.E.; Godson, C. Pro-Resolving Lipid Mediators: Regulators of Inflammation, Metabolism and Kidney Function. Nat. Rev. Nephrol. 2021, 17, 725–739. [Google Scholar] [CrossRef]
- Düsing, P.; Zietzer, A.; Goody, P.R.; Hosen, M.R.; Kurts, C.; Nickenig, G.; Jansen, F. Vascular Pathologies in Chronic Kidney Disease: Pathophysiological Mechanisms and Novel Therapeutic Approaches. J. Mol. Med. 2021, 99, 335–348. [Google Scholar] [CrossRef]
- Panizo, S.; Martínez-Arias, L.; Alonso-Montes, C.; Cannata, P.; Martín-Carro, B.; Fernández-Martín, J.L.; Naves-Díaz, M.; Carrillo-López, N.; Cannata-Andía, J.B. Fibrosis in Chronic Kidney Disease: Pathogenesis and Consequences. Int. J. Mol. Sci. 2021, 22, 408. [Google Scholar] [CrossRef]
- Kakani, E.; Elyamny, M.; Ayach, T.; El-Husseini, A. Pathogenesis and Management of Vascular Calcification in Ckd and Dialysis Patients. Semin. Dial. 2019, 32, 553–561. [Google Scholar] [CrossRef]
- Mace, M.L.; Gravesen, E.; Nordholm, A.; Egstrand, S.; Morevati, M.; Nielsen, C.; Kjaer, A.; Behets, G.; d′Haese, P.; Olgaard, K.; et al. Chronic Kidney Disease-Induced Vascular Calcification Impairs Bone Metabolism. J. Bone Miner. Res. 2021, 36, 510–522. [Google Scholar] [CrossRef]
- Singh, A.; Tandon, S.; Tandon, C. An Update on Vascular Calcification and Potential Therapeutics. Mol. Biol. Rep. 2021, 48, 887–896. [Google Scholar] [CrossRef]
- Zhang, Y.X.; Tang, R.N.; Wang, L.T.; Liu, B.C. Role of Crosstalk between Endothelial Cells and Smooth Muscle Cells in Vascular Calcification in Chronic Kidney Disease. Cell Prolif. 2021, 54, e12980. [Google Scholar] [CrossRef]
- Xie, X.; Liu, Y.; Perkovic, V.; Li, X.; Ninomiya, T.; Hou, W.; Zhao, N.; Liu, L.; Lv, J.; Zhang, H.; et al. Renin-Angiotensin System Inhibitors and Kidney and Cardiovascular Outcomes in Patients with Ckd: A Bayesian Network Meta-Analysis of Randomized Clinical Trials. Am. J. Kidney Dis. 2016, 67, 728–741. [Google Scholar] [CrossRef]
- Zhou, L.; Liu, Y. Wnt/Beta-Catenin Signaling and Renin-Angiotensin System in Chronic Kidney Disease. Curr. Opin. Nephrol. Hypertens. 2016, 25, 100–106. [Google Scholar] [CrossRef]
- Saldanha da Silva, A.A.; Rodrigues Prestes, T.R.; Lauar, A.O.; Finotti, B.B.; Simoes e Silva, A.C. Renin Angiotensin System and Cytokines in Chronic Kidney Disease: Clinical and Experimental Evidence. Protein Pept. Lett. 2017, 24, 799–808. [Google Scholar] [CrossRef]
- Nistor, I.; De Sutter, J.; Drechsler, C.; Goldsmith, D.; Soler, M.J.; Tomson, C.; Wiecek, A.; Donciu, M.D.; Bolignano, D.; Van Biesen, W.; et al. Effect of Renin-Angiotensin-Aldosterone System Blockade in Adults with Diabetes Mellitus and Advanced Chronic Kidney Disease Not on Dialysis: A Systematic Review and Meta-Analysis. Nephrol. Dial. Transplant. 2018, 33, 12–22. [Google Scholar] [CrossRef]
- Leon, S.J.; Tangri, N. The Use of Renin-Angiotensin System Inhibitors in Patients with Chronic Kidney Disease. Can. J. Cardiol. 2019, 35, 1220–1227. [Google Scholar] [CrossRef]
- Laffer, C.L.; Elijovich, F.; Sahinoz, M.; Pitzer, A.; Kirabo, A. New Insights into the Renin-Angiotensin System in Chronic Kidney Disease. Circ. Res. 2020, 127, 607–609. [Google Scholar] [CrossRef]
- Karpman, D.; Ståhl, A.L.; Arvidsson, I. Extracellular Vesicles in Renal Disease. Nat. Rev. Nephrol. 2017, 13, 545–562. [Google Scholar] [CrossRef]
- Ando, M.; Iwata, A.; Ozeki, Y.; Tsuchiya, K.; Akiba, T.; Nihei, H. Circulating Platelet-Derived Microparticles with Procoagulant Activity May Be a Potential Cause of Thrombosis in Uremic Patients. Kidney Int. 2002, 62, 1757–1763. [Google Scholar] [CrossRef]
- Faure, V.; Dou, L.; Sabatier, F.; Cerini, C.; Sampol, J.; Berland, Y.; Brunet, P.; Dignat-George, F. Elevation of Circulating Endothelial Microparticles in Patients with Chronic Renal Failure. J. Thromb. Haemost. 2006, 4, 566–573. [Google Scholar] [CrossRef] [PubMed]
- Burton, J.O.; Hamali, H.A.; Singh, R.; Abbasian, N.; Parsons, R.; Patel, A.K.; Goodall, A.H.; Brunskill, N.J. Elevated Levels of Procoagulant Plasma Microvesicles in Dialysis Patients. PLoS ONE 2013, 8, e72663. [Google Scholar] [CrossRef] [PubMed]
- Dursun, I.; Poyrazoglu, H.M.; Gunduz, Z.; Ulger, H.; Yýkýlmaz, A.; Dusunsel, R.; Patýroglu, T.; Gurgoze, M. The Relationship between Circulating Endothelial Microparticles and Arterial Stiffness and Atherosclerosis in Children with Chronic Kidney Disease. Nephrol. Dial. Transplant. 2009, 24, 2511–2518. [Google Scholar] [CrossRef] [PubMed]
- Charles, C.; Ferris, A.H. Chronic Kidney Disease. Prim. Care 2020, 47, 585–595. [Google Scholar] [CrossRef]
- Lane, C.A.; Hardy, J.; Schott, J.M. Alzheimer’s Disease. Eur. J. Neurol. 2018, 25, 59–70. [Google Scholar] [CrossRef]
- Scheltens, P.; De Strooper, B.; Kivipelto, M.; Holstege, H.; Chételat, G.; Teunissen, C.E.; Cummings, J.; van der Flier, W.M. Alzheimer’s Disease. Lancet 2021, 397, 1577–1590. [Google Scholar] [CrossRef]
- Ou, Z.; Jiang, T.; Gao, Q.; Tian, Y.Y.; Zhou, J.S.; Wu, L.; Shi, J.Q.; Zhang, Y.D. Mitochondrial-Dependent Mechanisms Are Involved in Angiotensin Ii-Induced Apoptosis in Dopaminergic Neurons. J. Renin-Angiotensin-Aldosterone Syst. 2016, 17, 1470320316672349. [Google Scholar] [CrossRef]
- He, S.; Wang, L.H.; Liu, Y.; Li, Y.Q.; Chen, H.T.; Xu, J.H.; Peng, W.; Lin, G.W.; Wei, P.P.; Li, B.; et al. Single-Cell Transcriptome Profiling of an Adult Human Cell Atlas of 15 Major Organs. Genome Biol. 2020, 21, 294. [Google Scholar] [CrossRef]
- Khan, S.; Barve, K.H.; Kumar, M.S. Recent Advancements in Pathogenesis, Diagnostics and Treatment of Alzheimer’s Disease. Curr. Neuropharmacol. 2020, 18, 1106–1125. [Google Scholar] [CrossRef]
- Vaz, M.; Silvestre, S. Alzheimer’s Disease: Recent Treatment Strategies. Eur. J. Pharmacol. 2020, 887, 173554. [Google Scholar] [CrossRef]
- Xin, S.H.; Tan, L.; Cao, X.; Yu, J.T.; Tan, L. Clearance of Amyloid Beta and Tau in Alzheimer’s Disease: From Mechanisms to Therapy. Neurotox. Res. 2018, 34, 733–748. [Google Scholar] [CrossRef]
- Tiwari, S.; Atluri, V.; Kaushik, A.; Yndart, A.; Nair, M. Alzheimer’s Disease: Pathogenesis, Diagnostics, and Therapeutics. Int. J. Nanomed. 2019, 14, 5541–5554. [Google Scholar] [CrossRef]
- Zhang, H.; Zheng, Y. Beta Amyloid Hypothesis in Alzheimer’s Disease: Pathogenesis, Prevention, and Management. Zhongguo Yi Xue Ke Xue Yuan Xue Bao 2019, 41, 702–708. [Google Scholar]
- Han, C.; Yang, Y.; Guan, Q.; Zhang, X.; Shen, H.; Sheng, Y.; Wang, J.; Zhou, X.; Li, W.; Guo, L.; et al. New Mechanism of Nerve Injury in Alzheimer’s Disease: Beta-Amyloid-Induced Neuronal Pyroptosis. J. Cell. Mol. Med. 2020, 24, 8078–8090. [Google Scholar] [CrossRef]
- Li, L.; Shi, R.; Gu, J.; Tung, Y.C.; Zhou, Y.; Zhou, D.; Wu, R.; Chu, D.; Jin, N.; Deng, K.; et al. Alzheimer’s Disease Brain Contains Tau Fractions with Differential Prion-Like Activities. Acta Neuropathol. Commun. 2021, 9, 28. [Google Scholar] [CrossRef]
- Breijyeh, Z.; Karaman, R. Comprehensive Review on Alzheimer’s Disease: Causes and Treatment. Molecules 2020, 25, 5789. [Google Scholar] [CrossRef]
- Ashrafian, H.; Zadeh, E.H.; Khan, R.H. Review on Alzheimer’s Disease: Inhibition of Amyloid Beta and Tau Tangle Formation. Int. J. Biol. Macromol. 2021, 167, 382–394. [Google Scholar] [CrossRef]
- Hill, A.F. Extracellular Vesicles and Neurodegenerative Diseases. J. Neurosci. 2019, 39, 9269–9273. [Google Scholar] [CrossRef]
- Jiang, L.; Dong, H.; Cao, H.; Ji, X.; Luan, S.; Liu, J. Exosomes in Pathogenesis, Diagnosis, and Treatment of Alzheimer’s Disease. Med. Sci. Monit. 2019, 25, 3329–3335. [Google Scholar] [CrossRef]
- Martins, T.S.; Trindade, D.; Vaz, M.; Campelo, I.; Almeida, M.; Trigo, G.; da Cruz e Silva, O.A.; Henriques, A.G. Diagnostic and Therapeutic Potential of Exosomes in Alzheimer’s Disease. J. Neurochem. 2021, 156, 162–181. [Google Scholar] [CrossRef]
- Zhang, T.; Ma, S.; Lv, J.; Wang, X.; Afewerky, H.K.; Li, H.; Lu, Y. The Emerging Role of Exosomes in Alzheimer’s Disease. Ageing Res. Rev. 2021, 68, 101321. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.Y.; Ma, S.; Huang, Y.; Wong, W.T. A Revisit on the Renin-Angiotensin System in Cardiovascular Biology. Trends Cardiovasc. Med. 2016, 26, 229–231. [Google Scholar] [CrossRef] [PubMed]
- Claflin, K.E.; Sandgren, J.A.; Lambertz, A.M.; Weidemann, B.J.; Littlejohn, N.K.; Burnett, C.M.; Pearson, N.A.; Morgan, D.A.; Gibson-Corley, K.N.; Rahmouni, K.; et al. Angiotensin At1a Receptors on Leptin Receptor-Expressing Cells Control Resting Metabolism. J. Clin. Investig. 2017, 127, 1414–1424. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Zhang, H.; Le, K.D.; Chao, J.; Gao, L. Activation of Central Angiotensin Type 2 Receptors Suppresses Norepinephrine Excretion and Blood Pressure in Conscious Rats. Am. J. Hypertens. 2011, 24, 724–730. [Google Scholar] [CrossRef] [PubMed]
- Chao, J.; Gao, J.; Parbhu, K.J.K.; Gao, L. Angiotensin Type 2 Receptors in the Intermediolateral Cell Column of the Spinal Cord: Negative Regulation of Sympathetic Nerve Activity and Blood Pressure. Int. J. Cardiol. 2013, 168, 4046–4055. [Google Scholar] [CrossRef]
- Gao, J.; Zucker, I.H.; Gao, L. Activation of Central Angiotensin Type 2 Receptors by Compound 21 Improves Arterial Baroreflex Sensitivity in Rats with Heart Failure. Am. J. Hypertens. 2014, 27, 1248–1256. [Google Scholar] [CrossRef]
- Harfouch, E.; Daoud, S. Allelic Variation in Hla-Drb1* Loci in Syrian Pemphigus Vulgaris Patients. Int. J. Dermatol. 2014, 53, 1460–1463. [Google Scholar] [CrossRef]
- Kemp, B.A.; Howell, N.L.; Gildea, J.J.; Keller, S.R.; Padia, S.H.; Carey, R.M. At2 Receptor Activation Induces Natriuresis and Lowers Blood Pressure. Circ. Res. 2014, 115, 388–399. [Google Scholar] [CrossRef]
- De Kloet, A.D.; Wang, L.; Ludin, J.A.; Smith, J.A.; Pioquinto, D.J.; Hiller, H.; Steckelings, U.M.; Scheuer, D.A.; Sumners, C.; Krause, E.G. Reporter Mouse Strain Provides a Novel Look at Angiotensin Type-2 Receptor Distribution in the Central Nervous System. Brain Struct. Funct. 2016, 221, 891–912. [Google Scholar] [CrossRef]
- Ji, Y.; Wang, Z.; Li, Z.; Zhang, A.; Jin, Y.; Chen, H.; Le, X. Angiotensin Ii Enhances Proliferation and Inflammation through At1/Pkc/Nf-Kappab Signaling Pathway in Hepatocellular Carcinoma Cells. Cell. Physiol. Biochem. 2016, 39, 13–32. [Google Scholar] [CrossRef]
- Kemp, B.A.; Howell, N.L.; Keller, S.R.; Gildea, J.J.; Padia, S.H.; Carey, R.M. At2 Receptor Activation Prevents Sodium Retention and Reduces Blood Pressure in Angiotensin Ii-Dependent Hypertension. Circ. Res. 2016, 119, 532–543. [Google Scholar] [CrossRef]
- Goel, R.; Bhat, S.A.; Hanif, K.; Nath, C.; Shukla, R. Angiotensin Ii Receptor Blockers Attenuate Lipopolysaccharide-Induced Memory Impairment by Modulation of Nf-Kappab-Mediated Bdnf/Creb Expression and Apoptosis in Spontaneously Hypertensive Rats. Mol. Neurobiol. 2018, 55, 1725–1739. [Google Scholar] [CrossRef]
- Gao, J.; Marc, Y.; Iturrioz, X.; Leroux, V.; Balavoine, F.; Llorens-Cortes, C. A New Strategy for Treating Hypertension by Blocking the Activity of the Brain Renin-Angiotensin System with Aminopeptidase a Inhibitors. Clin. Sci. 2014, 127, 135–148. [Google Scholar] [CrossRef]
- Sparks, M.A.; Crowley, S.D.; Gurley, S.B.; Mirotsou, M.; Coffman, T.M. Classical Renin-Angiotensin System in Kidney Physiology. Compr. Physiol. 2014, 4, 1201–1228. [Google Scholar]
- Park, B.M.; Gao, S.; Cha, S.A.; Park, B.H.; Kim, S.H. Cardioprotective Effects of Angiotensin Iii against Ischemic Injury Via the At2 Receptor and Katp Channels. Physiol. Rep. 2013, 1, e00151. [Google Scholar] [CrossRef]
- Yeatman, H.R.; Albiston, A.L.; Burns, P.; Chai, S.Y. Forebrain Neurone-Specific Deletion of Insulin-Regulated Aminopeptidase Causes Age Related Deficits in Memory. Neurobiol. Learn. Mem. 2016, 136, 174–182. [Google Scholar] [CrossRef]
- Paris, J.J.; Eans, S.O.; Mizrachi, E.; Reilley, K.J.; Ganno, M.L.; McLaughlin, J.P. Central Administration of Angiotensin Iv Rapidly Enhances Novel Object Recognition among Mice. Neuropharmacology 2013, 70, 247–253. [Google Scholar] [CrossRef]
- Hennrikus, M.; Gonzalez, A.A.; Prieto, M.C. The Prorenin Receptor in the Cardiovascular System and Beyond. Am. J. Physiol. Heart Circ. Physiol. 2018, 314, H139–H145. [Google Scholar] [CrossRef]
- Nunes-Silva, A.; Rocha, G.C.; Magalhaes, D.M.; Vaz, L.N.; de Faria, M.H.S.; Simoes e Silva, A.C. Physical Exercise and Ace2-Angiotensin-(1-7)-Mas Receptor Axis of the Renin Angiotensin System. Protein Pept. Lett. 2017, 24, 809–816. [Google Scholar] [CrossRef] [PubMed]
- Top 10 Health Technology Hazards for 2013 Are Named. OR Manag. 2013, 29, 16–19.
- Ola, M.S.; Alhomida, A.S.; Ferrario, C.M.; Ahmad, S. Role of Tissue Renin-Angiotensin System and the Chymase/Angiotensin-(1-12) Axis in the Pathogenesis of Diabetic Retinopathy. Curr. Med. Chem. 2017, 24, 3104–3114. [Google Scholar] [CrossRef] [PubMed]
- Stornetta, R.L.; Hawelu-Johnson, C.L.; Guyenet, P.G.; Lynch, K.R. Astrocytes Synthesize Angiotensinogen in Brain. Science 1988, 242, 1444–1446. [Google Scholar] [CrossRef]
- Labandeira-Garcia, J.L.; Rodríguez-Perez, A.I.; Garrido-Gil, P.; Rodriguez-Pallares, J.; Lanciego, J.L.; Guerra, M.J. Brain Renin-Angiotensin System and Microglial Polarization: Implications for Aging and Neurodegeneration. Front. Aging Neurosci. 2017, 9, 129. [Google Scholar] [CrossRef]
- Zeisel, A.; Muñoz-Manchado, A.B.; Codeluppi, S.; Lönnerberg, P.; La Manno, G.; Juréus, A.; Marques, S.; Munguba, H.; He, L.; Betsholtz, C.; et al. Brain Structure. Cell Types in the Mouse Cortex and Hippocampus Revealed by Single-Cell Rna-Seq. Science 2015, 347, 1138–1142. [Google Scholar] [CrossRef]
- Agarwal, D.; Sandor, C.; Volpato, V.; Caffrey, T.M.; Monzón-Sandoval, J.; Bowden, R.; Alegre-Abarrategui, J.; Wade-Martins, R.; Webber, C. A Single-Cell Atlas of the Human Substantia Nigra Reveals Cell-Specific Pathways Associated with Neurological Disorders. Nat. Commun. 2020, 11, 4183. [Google Scholar] [CrossRef]
- Kanton, S.; Boyle, M.J.; He, Z.; Santel, M.; Weigert, A.; Sanchís-Calleja, F.; Guijarro, P.; Sidow, L.; Fleck, J.S.; Han, D.; et al. Organoid Single-Cell Genomic Atlas Uncovers Human-Specific Features of Brain Development. Nature 2019, 574, 418–422. [Google Scholar] [CrossRef]
- Rivas-Santisteban, R.; Lillo, J.; Muñoz, A.; Rodríguez-Pérez, A.I.; Labandeira-García, J.L.; Navarro, G.; Franco, R. Novel Interactions Involving the Mas Receptor Show Potential of the Renin-Angiotensin System in the Regulation of Microglia Activation: Altered Expression in Parkinsonism and Dyskinesia. Neurotherapeutics 2021, 18, 998–1016. [Google Scholar] [CrossRef]
- Rivas-Santisteban, R.; Rodriguez-Perez, A.I.; Muñoz, A.; Reyes-Resina, I.; Labandeira-García, J.L.; Navarro, G.; Franco, R. Angiotensin At1 and At2 Receptor Heteromer Expression in the Hemilesioned Rat Model of Parkinson’s Disease That Increases with Levodopa-Induced Dyskinesia. J. Neuroinflammation 2020, 17, 243. [Google Scholar] [CrossRef]
- Loera-Valencia, R.; Eroli, F.; Garcia-Ptacek, S.; Maioli, S. Brain Renin-Angiotensin System as Novel and Potential Therapeutic Target for Alzheimer’s Disease. Int. J. Mol. Sci. 2021, 22, 10139. [Google Scholar] [CrossRef]
- Cassader, M.; Ruiu, G.; Leoncavallo, A.R.; Pagano, G. Changes in Carbohydrate and Lipid Metabolism in Chronic Renal Insufficiency. Clin. Ter. 1987, 122, 271–279. [Google Scholar]
- Royea, J.; Hamel, E. Brain Angiotensin Ii and Angiotensin Iv Receptors as Potential Alzheimer’s Disease Therapeutic Targets. Geroscience 2020, 42, 1237–1256. [Google Scholar] [CrossRef] [PubMed]
- Mowry, F.E.; Peaden, S.C.; Stern, J.E.; Biancardi, V.C. Tlr4 and At1r Mediate Blood-Brain Barrier Disruption, Neuroinflammation, and Autonomic Dysfunction in Spontaneously Hypertensive Rats. Pharmacol. Res. 2021, 174, 105877. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, M.; Berdasco, C.; Lazartigues, E. Brain Angiotensin Converting Enzyme-2 in Central Cardiovascular Regulation. Clin. Sci. 2020, 134, 2535–2547. [Google Scholar] [CrossRef]
- Maning, J.; Negussie, S.; Clark, M.A.; Lymperopoulos, A. Biased Agonism/Antagonism at the Angii-At1 Receptor: Implications for Adrenal Aldosterone Production and Cardiovascular Therapy. Pharmacol. Res. 2017, 125, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Dang, Z.; Su, S.; Jin, G.; Nan, X.; Ma, L.; Li, Z.; Lu, D.; Ge, R. Tsantan Sumtang Attenuated Chronic Hypoxia-Induced Right Ventricular Structure Remodeling and Fibrosis by Equilibrating Local Ace-Angii-At1r/Ace2-Ang1-7-Mas Axis in Rat. J. Ethnopharmacol. 2020, 250, 112470. [Google Scholar] [CrossRef]
- Barthold, D.; Joyce, G.; Brinton, R.D.; Wharton, W.; Kehoe, P.G.; Zissimopoulos, J. Association of Combination Statin and Antihypertensive Therapy with Reduced Alzheimer’s Disease and Related Dementia Risk. PLoS ONE 2020, 15, e0229541. [Google Scholar] [CrossRef]
- Ouk, M.; Wu, C.Y.; Rabin, J.S.; Jackson, A.; Edwards, J.D.; Ramirez, J.; Masellis, M.; Swartz, R.H.; Herrmann, N.; Lanctot, K.L.; et al. The Use of Angiotensin-Converting Enzyme Inhibitors vs. Angiotensin Receptor Blockers and Cognitive Decline in Alzheimer’s Disease: The Importance of Blood-Brain Barrier Penetration and Apoe Epsilon4 Carrier Status. Alzheimers Res. Ther. 2021, 13, 43. [Google Scholar] [CrossRef]
- Miners, J.S.; Ashby, E.; Van Helmond, Z.; Chalmers, K.A.; Palmer, L.E.; Love, S.; Kehoe, P.G. Angiotensin-Converting Enzyme (Ace) Levels and Activity in Alzheimer’s Disease, and Relationship of Perivascular Ace-1 to Cerebral Amyloid Angiopathy. Neuropathol. Appl. Neurobiol. 2008, 34, 181–193. [Google Scholar] [CrossRef]
- Kehoe, P.G.; Wong, S.; Al Mulhim, N.; Palmer, L.E.; Miners, J.S. Angiotensin-Converting Enzyme 2 Is Reduced in Alzheimer’s Disease in Association with Increasing Amyloid-Beta and Tau Pathology. Alzheimers Res. Ther. 2016, 8, 50. [Google Scholar] [CrossRef]
- Barnes, N.M.; Cheng, C.H.; Costall, B.; Naylor, R.J.; Williams, T.J.; Wischik, C.M. Angiotensin Converting Enzyme Density Is Increased in Temporal Cortex from Patients with Alzheimer’s Disease. Eur. J. Pharmacol. 1991, 200, 289–292. [Google Scholar] [CrossRef]
- Soto, M.E.; van Kan, G.A.; Nourhashemi, F.; Gillette-Guyonnet, S.; Cesari, M.; Cantet, C.; Rolland, Y.; Vellas, B. Angiotensin-Converting Enzyme Inhibitors and Alzheimer’s Disease Progression in Older Adults: Results from the Reseau Sur La Maladie D’alzheimer Francais Cohort. J. Am. Geriatr. Soc. 2013, 61, 1482–1488. [Google Scholar] [CrossRef]
- Li, N.C.; Lee, A.; Whitmer, R.A.; Kivipelto, M.; Lawler, E.; Kazis, L.E.; Wolozin, B. Use of Angiotensin Receptor Blockers and Risk of Dementia in a Predominantly Male Population: Prospective Cohort Analysis. BMJ 2010, 340, b5465. [Google Scholar] [CrossRef] [PubMed]
- Ongali, B.; Nicolakakis, N.; Tong, X.K.; Aboulkassim, T.; Papadopoulos, P.; Rosa-Neto, P.; Lecrux, C.; Imboden, H.; Hamel, E. Angiotensin Ii Type 1 Receptor Blocker Losartan Prevents and Rescues Cerebrovascular, Neuropathological and Cognitive Deficits in an Alzheimer’s Disease Model. Neurobiol. Dis. 2014, 68, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Fogari, R.; Mugellini, A.; Zoppi, A.; Derosa, G.; Pasotti, C.; Fogari, E.; Preti, P. Influence of Losartan and Atenolol on Memory Function in Very Elderly Hypertensive Patients. J. Hum. Hypertens. 2003, 17, 781–785. [Google Scholar] [CrossRef] [PubMed]
- Fournier, A.; Oprisiu-Fournier, R.; Serot, J.M.; Godefroy, O.; Achard, J.M.; Faure, S.; Mazouz, H.; Temmar, M.; Albu, A.; Bordet, R.; et al. Prevention of Dementia by Antihypertensive Drugs: How At1-Receptor-Blockers and Dihydropyridines Better Prevent Dementia in Hypertensive Patients Than Thiazides and Ace-Inhibitors. Expert Rev. Neurother. 2009, 9, 1413–1431. [Google Scholar] [CrossRef]
- Lemma, F.; Bombardieri, T.; Salibra, M.; Trovato, P.; De Pasquale, M.C.; Faraone, V.; Punturiero, R.; Di Mauro, S. Tumor Markers in the Diagnosis of Pancreatic Carcinoma. Minerva Med. 1989, 80, 645–649. [Google Scholar]
- Karbowska, M.; Hermanowicz, J.M.; Tankiewicz-Kwedlo, A.; Kalaska, B.; Kaminski, T.W.; Nosek, K.; Wisniewska, R.J.; Pawlak, D. Neurobehavioral Effects of Uremic Toxin-Indoxyl Sulfate in the Rat Model. Sci. Rep. 2020, 10, 9483. [Google Scholar] [CrossRef]
- Larsson, S.C.; Traylor, M.; Markus, H.S.; Michaëlsson, K. Serum Parathyroid Hormone, 25-Hydroxyvitamin D, and Risk of Alzheimer’s Disease: A Mendelian Randomization Study. Nutrients 2018, 10, 1243. [Google Scholar] [CrossRef]
- Assaraf, M.I.; Diaz, Z.; Liberman, A.; Miller, W.H., Jr.; Arvanitakis, Z.; Li, Y.; Bennett, D.A.; Schipper, H.M. Brain Erythropoietin Receptor Expression in Alzheimer Disease and Mild Cognitive Impairment. J. Neuropathol. Exp. Neurol. 2007, 66, 389–398. [Google Scholar] [CrossRef]
- Viviani, B.; Bartesaghi, S.; Corsini, E.; Villa, P.; Ghezzi, P.; Garau, A.; Galli, C.L.; Marinovich, M. Erythropoietin Protects Primary Hippocampal Neurons Increasing the Expression of Brain-Derived Neurotrophic Factor. J. Neurochem. 2005, 93, 412–421. [Google Scholar] [CrossRef]
- Ma, R.; Xiong, N.; Huang, C.; Tang, Q.; Hu, B.; Xiang, J.; Li, G. Erythropoietin Protects Pc12 Cells from Beta-Amyloid(25-35)-Induced Apoptosis Via Pi3k/Akt Signaling Pathway. Neuropharmacology 2009, 56, 1027–1034. [Google Scholar] [CrossRef]
- Tazangi, P.E.; Moosavi, S.M.S.; Shabani, M.; Haghani, M. Erythropoietin Improves Synaptic Plasticity and Memory Deficits by Decrease of the Neurotransmitter Release Probability in the Rat Model of Alzheimer’s Disease. Pharmacol. Biochem. Behav. 2015, 130, 15–21. [Google Scholar] [CrossRef]
- Yuyama, K.; Sun, H.; Sakai, S.; Mitsutake, S.; Okada, M.; Tahara, H.; Furukawa, J.I.; Fujitani, N.; Shinohara, Y.; Igarashi, Y. Decreased Amyloid-Beta Pathologies by Intracerebral Loading of Glycosphingolipid-Enriched Exosomes in Alzheimer Model Mice. J. Biol. Chem. 2014, 289, 24488–24498. [Google Scholar] [CrossRef]
- Yuyama, K.; Sun, H.; Mitsutake, S.; Igarashi, Y. Sphingolipid-Modulated Exosome Secretion Promotes Clearance of Amyloid-Beta by Microglia. J. Biol. Chem. 2012, 287, 10977–10989. [Google Scholar] [CrossRef]
- Kehoe, P.G.; Miners, S.; Love, S. Angiotensins in Alzheimer’s Disease—Friend or Foe? Trends Neurosci. 2009, 32, 619–628. [Google Scholar] [CrossRef]
- Zhu, D.; Shi, J.; Zhang, Y.; Wang, B.; Liu, W.; Chen, Z.; Tong, Q. Central Angiotensin Ii Stimulation Promotes Beta Amyloid Production in Sprague Dawley Rats. PLoS ONE 2011, 6, e16037. [Google Scholar]
- Zou, K.; Yamaguchi, H.; Akatsu, H.; Sakamoto, T.; Ko, M.; Mizoguchi, K.; Gong, J.S.; Yu, W.; Yamamoto, T.; Kosaka, K.; et al. Angiotensin-Converting Enzyme Converts Amyloid Beta-Protein 1-42 (Abeta(1-42)) to Abeta(1-40), and Its Inhibition Enhances Brain Abeta Deposition. J. Neurosci. 2007, 27, 8628–8635. [Google Scholar] [CrossRef]
- Hu, X.; Liou, A.K.; Leak, R.K.; Xu, M.; An, C.; Suenaga, J.; Shi, Y.; Gao, Y.; Zheng, P.; Chen, J. Neurobiology of Microglial Action in Cns Injuries: Receptor-Mediated Signaling Mechanisms and Functional Roles. Prog. Neurobiol. 2014, 119–120, 60–84. [Google Scholar] [CrossRef]
- Prinz, M.; Priller, J. Microglia and Brain Macrophages in the Molecular Age: From Origin to Neuropsychiatric Disease. Nat. Rev. Neurosci. 2014, 15, 300–312. [Google Scholar] [CrossRef]
- Cui, C.; Xu, P.; Li, G.; Qiao, Y.; Han, W.; Geng, C.; Liao, D.; Yang, M.; Chen, D.; Jiang, P. Vitamin D Receptor Activation Regulates Microglia Polarization and Oxidative Stress in Spontaneously Hypertensive Rats and Angiotensin Ii-Exposed Microglial Cells: Role of Renin-Angiotensin System. Redox Biol. 2019, 26, 101295. [Google Scholar] [CrossRef]
- Jackson, L.; Eldahshan, W.; Fagan, S.C.; Ergul, A. Within the Brain: The Renin Angiotensin System. Int. J. Mol. Sci. 2018, 19, 876. [Google Scholar] [CrossRef] [PubMed]
- Biancardi, V.C.; Stranahan, A.M.; Krause, E.G.; de Kloet, A.D.; Stern, J.E. Cross Talk between At1 Receptors and Toll-Like Receptor 4 in Microglia Contributes to Angiotensin Ii-Derived Ros Production in the Hypothalamic Paraventricular Nucleus. Am. J. Physiol. Heart Circ. Physiol. 2016, 310, H404–H415. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, K.E.; Koronyo, Y.; Salumbides, B.C.; Sheyn, J.; Pelissier, L.; Lopes, D.H.; Shah, K.H.; Bernstein, E.A.; Fuchs, D.T.; Yu, J.J.Y.; et al. Angiotensin-Converting Enzyme Overexpression in Myelomonocytes Prevents Alzheimer’s-Like Cognitive Decline. J. Clin. Investig. 2014, 124, 1000–1012. [Google Scholar] [CrossRef] [PubMed]
- Arroja, M.M.C.; Reid, E.; McCabe, C. Therapeutic Potential of the Renin Angiotensin System in Ischaemic Stroke. Exp. Transl. Stroke Med. 2016, 8, 8. [Google Scholar] [CrossRef]
- Regenhardt, R.W.; Desland, F.; Mecca, A.P.; Pioquinto, D.J.; Afzal, A.; Mocco, J.; Sumners, C. Anti-Inflammatory Effects of Angiotensin-(1-7) in Ischemic Stroke. Neuropharmacology 2013, 71, 154–163. [Google Scholar] [CrossRef]
- Fouda, A.Y.; Pillai, B.; Dhandapani, K.M.; Ergul, A.; Fagan, S.C. Role of Interleukin-10 in the Neuroprotective Effect of the Angiotensin Type 2 Receptor Agonist, Compound 21, after Ischemia/Reperfusion Injury. Eur. J. Pharmacol. 2017, 799, 128–134. [Google Scholar] [CrossRef]
- Liu, M.; Shi, P.; Sumners, C. Direct Anti-Inflammatory Effects of Angiotensin-(1-7) on Microglia. J. Neurochem. 2016, 136, 163–171. [Google Scholar] [CrossRef]
- Zenaro, E.; Piacentino, G.; Constantin, G. The Blood-Brain Barrier in Alzheimer’s Disease. Neurobiol. Dis. 2017, 107, 41–56. [Google Scholar] [CrossRef]
- Sweeney, M.D.; Zhao, Z.; Montagne, A.; Nelson, A.R.; Zlokovic, B.V. Blood-Brain Barrier: From Physiology to Disease and Back. Physiol. Rev. 2019, 99, 21–78. [Google Scholar] [CrossRef]
- Obermeier, B.; Daneman, R.; Ransohoff, R.M. Development, Maintenance and Disruption of the Blood-Brain Barrier. Nat. Med. 2013, 19, 1584–1596. [Google Scholar] [CrossRef]
- Maarouf, C.L.; Kokjohn, T.A.; Whiteside, C.M.; Macias, M.P.; Kalback, W.M.; Sabbagh, M.N.; Beach, T.G.; Vassar, R.; Roher, A.E. Molecular Differences and Similarities between Alzheimer’s Disease and the 5xfad Transgenic Mouse Model of Amyloidosis. Biochem. Insights 2013, 6, BCI-S13025. [Google Scholar] [CrossRef]
- Lin, B.; Hasegawa, Y.; Takane, K.; Koibuchi, N.; Cao, C.; Kim-Mitsuyama, S. High-Fat-Diet Intake Enhances Cerebral Amyloid Angiopathy and Cognitive Impairment in a Mouse Model of Alzheimer’s Disease, Independently of Metabolic Disorders. J. Am. Heart Assoc. 2016, 5, e003154. [Google Scholar] [CrossRef]
- Orihuela, R.; McPherson, C.A.; Harry, G.J. Microglial M1/M2 Polarization and Metabolic States. Br. J. Pharmacol. 2016, 173, 649–665. [Google Scholar] [CrossRef]
- Hamed, S.A. Neurologic Conditions and Disorders of Uremic Syndrome of Chronic Kidney Disease: Presentations, Causes, and Treatment Strategies. Expert Rev. Clin. Pharmacol. 2019, 12, 61–90. [Google Scholar] [CrossRef]
- Hailpern, S.M.; Melamed, M.L.; Cohen, H.W.; Hostetter, T.H. Moderate Chronic Kidney Disease and Cognitive Function in Adults 20 to 59 Years of Age: Third National Health and Nutrition Examination Survey (Nhanes Iii). J. Am. Soc. Nephrol. 2007, 18, 2205–2213. [Google Scholar] [CrossRef]
- McQuillan, R.; Jassal, S.V. Neuropsychiatric Complications of Chronic Kidney Disease. Nat. Rev. Nephrol. 2010, 6, 471–479. [Google Scholar] [CrossRef]
- Vannorsdall, T.D.; Jinnah, H.A.; Gordon, B.; Kraut, M.; Schretlen, D.J. Cerebral Ischemia Mediates the Effect of Serum Uric Acid on Cognitive Function. Stroke 2008, 39, 3418–3420. [Google Scholar] [CrossRef]
- Tian, M.; Zhu, D.; Xie, W.; Shi, J. Central Angiotensin Ii-Induced Alzheimer-Like Tau Phosphorylation in Normal Rat Brains. FEBS Lett. 2012, 586, 3737–3745. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.; Gao, L.; Shi, J.; Lu, J.; Wang, Y.; Zhang, Y. Angiotensin-(1-7) Modulates Renin-Angiotensin System Associated with Reducing Oxidative Stress and Attenuating Neuronal Apoptosis in the Brain of Hypertensive Rats. Pharmacol. Res. 2013, 67, 84–93. [Google Scholar] [CrossRef]
- Watanabe, K.; Sato, E.; Mishima, E.; Watanabe, M.; Abe, T.; Takahashi, N.; Nakayama, M. Effect of Uremic Toxins on Hippocampal Cell Damage: Analysis in Vitro and in Rat Model of Chronic Kidney Disease. Heliyon 2021, 7, e06221. [Google Scholar] [CrossRef]
- Haruyama, N.; Fujisaki, K.; Yamato, M.; Eriguchi, M.; Noguchi, H.; Torisu, K.; Tsuruya, K.; Kitazono, T. Improvement in Spatial Memory Dysfunction by Telmisartan through Reduction of Brain Angiotensin Ii and Oxidative Stress in Experimental Uremic Mice. Life Sci. 2014, 113, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Burek, M.; Burmester, S.; Salvador, E.; Möller-Ehrlich, K.; Schneider, R.; Roewer, N.; Nagai, M.; Förster, C.Y. Kidney Ischemia/Reperfusion Injury Induces Changes in the Drug Transporter Expression at the Blood-Brain Barrier in Vivo and in Vitro. Front. Physiol. 2020, 11, 569881. [Google Scholar] [CrossRef]
- Ohtsuki, S.; Asaba, H.; Takanaga, H.; Deguchi, T.; Hosoya, K.I.; Otagiri, M.; Terasaki, T. Role of Blood-Brain Barrier Organic Anion Transporter 3 (Oat3) in the Efflux of Indoxyl Sulfate, a Uremic Toxin: Its Involvement in Neurotransmitter Metabolite Clearance from the Brain. J. Neurochem. 2002, 83, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Wakhloo, D.; Scharkowski, F.; Curto, Y.; Butt, U.J.; Bansal, V.; Steixner-Kumar, A.A.; Wüstefeld, L.; Rajput, A.; Arinrad, S.; Zillmann, M.R.; et al. Functional Hypoxia Drives Neuroplasticity and Neurogenesis Via Brain Erythropoietin. Nat. Commun. 2020, 11, 1313. [Google Scholar] [CrossRef]
- Li, G.; Ma, R.; Huang, C.; Tang, Q.; Fu, Q.; Liu, H.; Hu, B.; Xiang, J. Protective Effect of Erythropoietin on Beta-Amyloid-Induced Pc12 Cell Death through Antioxidant Mechanisms. Neurosci. Lett. 2008, 442, 143–147. [Google Scholar] [CrossRef]
- Hooshmandi, E.; Motamedi, F.; Moosavi, M.; Katinger, H.; Zakeri, Z.; Zaringhalam, J.; Maghsoudi, A.; Ghasemi, R.; Maghsoudi, N. Cepo-Fc (an Epo Derivative) Protects Hippocampus against Abeta-Induced Memory Deterioration: A Behavioral and Molecular Study in a Rat Model of Abeta Toxicity. Neuroscience 2018, 388, 405–417. [Google Scholar] [CrossRef]
- Cevik, B.; Solmaz, V.; Yigitturk, G.; Cavusoğlu, T.; Peker, G.; Erbas, O. Neuroprotective Effects of Erythropoietin on Alzheimer’s Dementia Model in Rats. Adv. Clin. Exp. Med. 2017, 26, 23–29. [Google Scholar] [CrossRef]
- Maurice, T.; Mustafa, M.H.; Desrumaux, C.; Keller, E.; Naert, G.; Garcia-Barcelo, M.D.L.C.; Cruz, Y.R.; Rodriguez, J.C.G. Intranasal Formulation of Erythropoietin (Epo) Showed Potent Protective Activity against Amyloid Toxicity in the Abeta25-35 Non-Transgenic Mouse Model of Alzheimer’s Disease. J. Psychopharmacol. 2013, 27, 1044–1057. [Google Scholar] [CrossRef]
- Carmona, A.; Guerrero, F.; Buendia, P.; Obrero, T.; Aljama, P.; Carracedo, J. Microvesicles Derived from Indoxyl Sulfate Treated Endothelial Cells Induce Endothelial Progenitor Cells Dysfunction. Front. Physiol. 2017, 8, 666. [Google Scholar] [CrossRef]
- Ryu, J.H.; Park, H.; Kim, S.J. The Effects of Indoxyl Sulfate-Induced Endothelial Microparticles on Neointimal Hyperplasia Formation in an Ex Vivo Model. Ann. Surg. Treat. Res. 2017, 93, 11–17. [Google Scholar] [CrossRef]
- Alique, M.; Bodega, G.; Corchete, E.; García-Menéndez, E.; de Sequera, P.; Luque, R.; Rodríguez-Padrón, D.; Marqués, M.; Portolés, J.; Carracedo, J.; et al. Microvesicles from Indoxyl Sulfate-Treated Endothelial Cells Induce Vascular Calcification in Vitro. Comput. Struct. Biotechnol. J. 2020, 18, 953–966. [Google Scholar] [CrossRef] [PubMed]
- Sinha, M.S.; Ansell-Schultz, A.; Civitelli, L.; Hildesjö, C.; Larsson, M.; Lannfelt, L.; Ingelsson, M.; Hallbeck, M. Alzheimer’s Disease Pathology Propagation by Exosomes Containing Toxic Amyloid-Beta Oligomers. Acta Neuropathol. 2018, 136, 41–56. [Google Scholar] [CrossRef]
- McAvoy, K.M.; Sait, H.R.; Marsh, G.; Peterson, M.; Reynolds, T.L.; Gagnon, J.; Geisler, S.; Leach, P.; Roberts, C.; Cahir-McFarland, E.; et al. Cell-Autonomous and Non-Cell Autonomous Effects of Neuronal Bin1 Loss in Vivo. PLoS ONE 2019, 14, e0220125. [Google Scholar] [CrossRef]
- Ruan, Z.; Pathak, D.; Venkatesan Kalavai, S.; Yoshii-Kitahara, A.; Muraoka, S.; Bhatt, N.; Takamatsu-Yukawa, K.; Hu, J.; Wang, Y.; Hersh, S.; et al. Alzheimer’s Disease Brain-Derived Extracellular Vesicles Spread Tau Pathology in Interneurons. Brain 2021, 144, 288–309. [Google Scholar] [CrossRef]
- Kohan, D.E.; Barton, M. Endothelin and Endothelin Antagonists in Chronic Kidney Disease. Kidney Int. 2014, 86, 896–904. [Google Scholar] [CrossRef]
- Eroglu, E.; Kocyigit, I.; Lindholm, B. The Endothelin System as Target for Therapeutic Interventions in Cardiovascular and Renal Disease. Clin. Chim. Acta 2020, 506, 92–106. [Google Scholar] [CrossRef]
- Heerspink, H.J.; Xie, D.; Bakris, G.; Correa-Rotter, R.; Hou, F.F.; Kitzman, D.W.; Kohan, D.; Makino, H.; McMurray, J.J.; Perkovic, V.; et al. Early Response in Albuminuria and Long-Term Kidney Protection During Treatment with an Endothelin Receptor Antagonist: A Prespecified Analysis from the Sonar Trial. J. Am. Soc. Nephrol. 2021, 32, 2900–2911. [Google Scholar] [CrossRef]
- Hsu, Y.H.; Zheng, C.M.; Chou, C.L.; Chen, Y.J.; Lee, Y.H.; Lin, Y.F.; Chiu, H.W. Therapeutic Effect of Endothelin-Converting Enzyme Inhibitor on Chronic Kidney Disease through the Inhibition of Endoplasmic Reticulum Stress and the Nlrp3 Inflammasome. Biomedicines 2021, 9, 398. [Google Scholar] [CrossRef]
- Palmer, J.C.; Barker, R.; Kehoe, P.G.; Love, S. Endothelin-1 Is Elevated in Alzheimer’s Disease and Upregulated by Amyloid-Beta. J. Alzheimers Dis. 2012, 29, 853–861. [Google Scholar] [CrossRef]
- Karakaya, E.; Abdul, Y.; Chowdhury, N.; Wellslager, B.; Jamil, S.; Albayram, O.; Yilmaz, Ö.; Ergul, A. Porphyromonas Gingivalis Infection Upregulates the Endothelin (Et) System in Brain Microvascular Endothelial Cells. Can. J. Physiol. Pharmacol. 2022, 100, 679–688. [Google Scholar] [CrossRef]
- Gulati, A.; Hornick, M.G.; Briyal, S.; Lavhale, M.S. A Novel Neuroregenerative Approach Using Et(B) Receptor Agonist, Irl-1620, to Treat Cns Disorders. Physiol. Res. 2018, 67 (Suppl. 1), S95–S113. [Google Scholar] [CrossRef]
- Pacheco-Quinto, J.; Eckman, E.A. Endothelin-Converting Enzymes Degrade Intracellular Beta-Amyloid Produced within the Endosomal/Lysosomal Pathway and Autophagosomes. J. Biol. Chem. 2013, 288, 5606–5615. [Google Scholar] [CrossRef] [PubMed]
- Pacheco-Quinto, J.; Eckman, C.B.; Eckman, E.A. Major Amyloid-Beta-Degrading Enzymes, Endothelin-Converting Enzyme-2 and Neprilysin, Are Expressed by Distinct Populations of Gabaergic Interneurons in Hippocampus and Neocortex. Neurobiol. Aging 2016, 48, 83–92. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| Factor | Attributes | Reference | |
|---|---|---|---|
| Renin-Angiotensin System | ACE1 | ACE1 accumulates in severe AD patients. | Miners et al. [118] |
| ACE2 | ACE2 activated, causes lower hippocampal Aβ and restored cognition | Kehoe et al. [119] | |
| ACEI | ACEI was higher in the temporal cortex of Alzheimer’s patients | Barnes et al. [120] | |
| ACEI | ACEI causes a slower rate of cognitive decline | Soto et al. [121] | |
| ARBs | ARBs significantly reduce the incidence and progression of Alzheimer’s disease and dementia | Li et al. [122] | |
| ARBs | ARBs rescue cerebrovascular and cognitive function in adults. | Ongali et al. [123] | |
| ARBs | ARBs could improve cognitive function, in particular immediate and delayed memory. | Fogari et al. [124] | |
| ACEI and ARBs | ARBs may have greater cognition protective effects than ACEI | Fournier et al. [125] | |
| Uremic toxins | Uric Acid | Systemic hyperuricemia induces cognitive dysfunction | Lemma et al. [126] |
| Indoxyl Sulfate | Chronic exposure to Indoxyl Sulfate leads to reduced locomotor activity and spatial memory, as well as increased stress sensitivity, and apathetic behavior | Karbowska et al. [127] | |
| Parathyroid hormone | Parathyroid hormone decline brain impairment by vitamin D | Larsson et al. [128] | |
| Erythropoietin | EPO | Epo-EpoR significant cytoprotection by antioxidant, antiapoptotic, anti-inflammatory, neurotrophic, angiogenic, and synaptogenic activities. | Assaraf et al. [129] Viviani et al. [130] Ma et al. [131] Esmaeili Tazangi et al. [132] |
| Extracellular Vehicles | EVs | EVs contribute to peptide clearance from the extracellular space and reducing Aβ pathology | Yuyama et al. [133,134] Soares Martins et al. [80] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, K.; Zheng, Z.-R.; Meng, Y. Pathogenesis of Chronic Kidney Disease Is Closely Bound up with Alzheimer’s Disease, Especially via the Renin-Angiotensin System. J. Clin. Med. 2023, 12, 1459. https://doi.org/10.3390/jcm12041459
Ma K, Zheng Z-R, Meng Y. Pathogenesis of Chronic Kidney Disease Is Closely Bound up with Alzheimer’s Disease, Especially via the Renin-Angiotensin System. Journal of Clinical Medicine. 2023; 12(4):1459. https://doi.org/10.3390/jcm12041459
Chicago/Turabian StyleMa, Ke, Zi-Run Zheng, and Yu Meng. 2023. "Pathogenesis of Chronic Kidney Disease Is Closely Bound up with Alzheimer’s Disease, Especially via the Renin-Angiotensin System" Journal of Clinical Medicine 12, no. 4: 1459. https://doi.org/10.3390/jcm12041459
APA StyleMa, K., Zheng, Z.-R., & Meng, Y. (2023). Pathogenesis of Chronic Kidney Disease Is Closely Bound up with Alzheimer’s Disease, Especially via the Renin-Angiotensin System. Journal of Clinical Medicine, 12(4), 1459. https://doi.org/10.3390/jcm12041459