
Molecular Alterations and Putative Therapeutic Targeting of Planar Cell Polarity Proteins in Breast Cancer

Abstract
1. Introduction
2. Methods
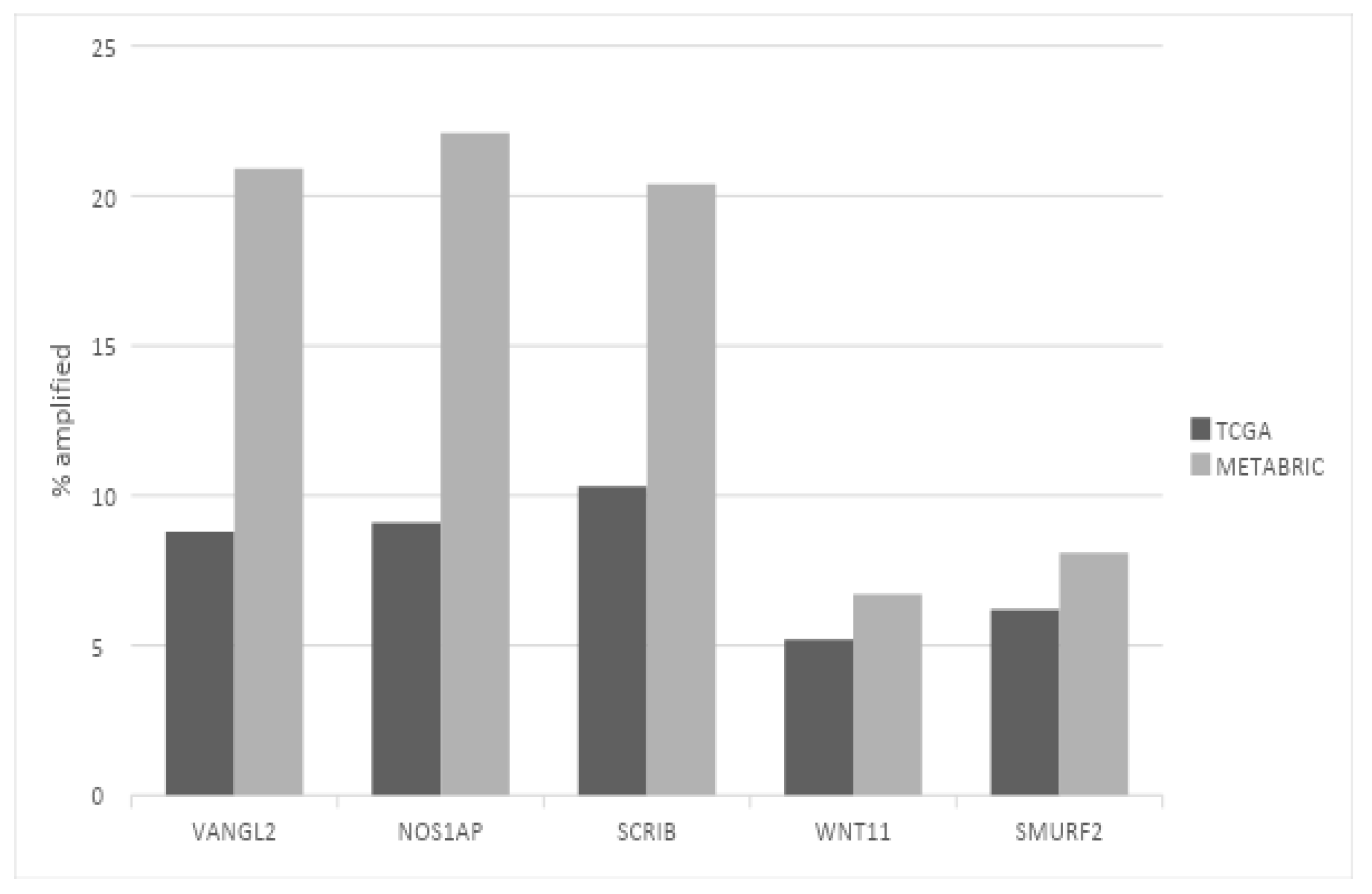
3. Results
4. Discussion
5. Conclusions
Funding
Data Availability Statement
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef] [PubMed]
- DeSantis, C.E.; Ma, J.; Gaudet, M.M.; Newman, L.A.; Miller, K.D.; Goding Sauer, A.; Jemal, A.; Siegel, R.L. Breast cancer statistics, 2019. CA Cancer J. Clin. 2019, 69, 438–451. [Google Scholar] [CrossRef] [PubMed]
- Sørlie, T.; Perou, C.M.; Tibshirani, R.; Aas, T.; Geisler, S.; Johnsen, H.; Hastie, T.; Eisen, M.B.; van de Rijn, M.; Jeffrey, S.S.; et al. Gene expression patterns of breast carcinomas distinguish tumor subclasses with clinical implications. Proc. Natl. Acad. Sci. USA 2001, 98, 10869–10874. [Google Scholar] [CrossRef] [PubMed]
- Voutsadakis, I.A. Chromosome 17 centromere amplification and chromosomal instability (CIN) in breast cancer: Pathogenic and therapeutic implications. Neoplasma 2019, 66, 859–869. [Google Scholar] [CrossRef] [PubMed]
- Voutsadakis, I.A. Clinical Implications of Chromosomal Instability (CIN) and Kinetochore Abnormalities in Breast Cancers. Mol. Diagn. Ther. 2019, 23, 707–721. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.S.; Martin, M.; Rugo, H.S.; Jones, S.; Im, S.A.; Gelmon, K.; Harbeck, N.; Lipatov, O.N.; Walshe, J.M.; Moulder, S.; et al. Palbociclib and Letrozole in Advanced Breast Cancer. N. Engl. J. Med. 2016, 375, 1925–1936. [Google Scholar] [CrossRef] [PubMed]
- Stravodimou, A.; Voutsadakis, I.A. The Future of ER+/HER2- Metastatic Breast Cancer Therapy: Beyond PI3K Inhibitors. Anticancer Res. 2020, 40, 4829–4841. [Google Scholar] [CrossRef]
- Slamon, D.; Eiermann, W.; Robert, N.; Pienkowski, T.; Martin, M.; Press, M.; Mackey, J.; Glaspy, J.; Chan, A.; Pawlicki, M.; et al. Adjuvant trastuzumab in HER2-positive breast cancer. N. Engl. J. Med. 2011, 365, 1273–1283. [Google Scholar] [CrossRef]
- Curigliano, G.; Mueller, V.; Borges, V.; Hamilton, E.; Hurvitz, S.; Loi, S.; Murthy, R.; Okines, A.; Paplomata, E.; Cameron, D.; et al. Tucatinib versus placebo added to trastuzumab and capecitabine for patients with pretreated HER2+ metastatic breast cancer with and without brain metastases (HER2CLIMB): Final overall survival analysis. Ann. Oncol. 2022, 33, 321–329. [Google Scholar] [CrossRef]
- Cortes, J.; Cescon, D.W.; Rugo, H.S.; Nowecki, Z.; Im, S.A.; Yusof, M.M.; Gallardo, C.; Lipatov, O.; Barrios, C.H.; Holgado, E.; et al. Pembrolizumab plus chemotherapy versus placebo plus chemotherapy for previously untreated locally recurrent inoperable or metastatic triple-negative breast cancer (KEYNOTE-355): A randomised, placebo-controlled, double-blind, phase 3 clinical trial. Lancet 2020, 396, 1817–1828. [Google Scholar] [CrossRef]
- Voutsadakis, I.A. Immune Blockade Inhibition in Breast Cancer. Anticancer Res. 2016, 36, 5607–5622. [Google Scholar] [CrossRef]
- Robson, M.; Im, S.A.; Senkus, E.; Xu, B.; Domchek, S.M.; Masuda, N.; Delaloge, S.; Li, W.; Tung, N.; Armstrong, A.; et al. Olaparib for Metastatic Breast Cancer in Patients with a Germline BRCA Mutation. N. Engl. J. Med. 2017, 377, 523–533. [Google Scholar] [CrossRef]
- Čada, Š.; Bryja, V. Local Wnt signalling in the asymmetric migrating vertebrate cells. Semin. Cell Dev. Biol. 2022, 125, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Montcouquiol, M.; Rachel, R.A.; Lanford, P.J.; Copeland, N.G.; Jenkins, N.A.; Kelley, M.W. Identification of Vangl2 and Scrb1 as planar polarity genes in mammals. Nature 2003, 423, 173–177. [Google Scholar] [CrossRef]
- Baker, L.; BeGora, M.; Au Yeung, F.; Feigin, M.E.; Rosenberg, A.Z.; Lowe, S.W.; Kislinger, T.; Muthuswamy, S.K. Scribble is required for pregnancy-induced alveologenesis in the adult mammary gland. J. Cell Sci. 2016, 129, 2307–2315. [Google Scholar] [CrossRef] [PubMed]
- VanderVorst, K.; Hatakeyama, J.; Berg, A.; Lee, H.; Carraway, K.L., 3rd. Cellular and molecular mechanisms underlying planar cell polarity pathway contributions to cancer malignancy. Semin. Cell Dev. Biol. 2018, 81, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Anastas, J.N.; Biechele, T.L.; Robitaille, M.; Muster, J.; Allison, K.H.; Angers, S.; Moon, R.T. A protein complex of SCRIB, NOS1AP and VANGL1 regulates cell polarity and migration, and is associated with breast cancer progression. Oncogene 2012, 31, 3696–3708. [Google Scholar] [CrossRef]
- Cancer Genome Atlas Network. Comprehensive molecular portraits of human breast tumours. Nature 2012, 490, 61–70. [Google Scholar] [CrossRef]
- Curtis, C.; Shah, S.P.; Chin, S.F.; Turashvili, G.; Rueda, O.M.; Dunning, M.J.; Speed, D.; Lynch, A.G.; Samarajiwa, S.; Yuan, Y.; et al. The genomic and transcriptomic architecture of 2,000 breast tumours reveals novel subgroups. Nature 2012, 486, 346–352. [Google Scholar] [CrossRef]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio cancer genomics portal: An open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci. Signal 2013, 6, pl1. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.M.; Shih, J.; Ha, G.; Gao, G.F.; Zhang, X.; Berger, A.C.; Schumacher, S.E.; Wang, C.; Hu, H.; Liu, J.; et al. Genomic and Functional Approaches to Understanding Cancer Aneuploidy. Cancer Cell 2018, 33, 676–689. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed]
- Chakravarty, D.; Gao, J.; Phillips, S.M.; Kundra, R.; Zhang, H.; Wang, J.; Rudolph, J.E.; Yaeger, R.; Soumerai, T.; Nissan, M.H.; et al. OncoKB: A Precision Oncology Knowledge Base. JCO Precis. Oncol. 2017, 2017, PO.17.00011. [Google Scholar] [CrossRef]
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef] [PubMed]
- Barretina, J.; Caponigro, G.; Stransky, N.; Venkatesan, K.; Margolin, A.A.; Kim, S.; Wilson, C.J.; Lehár, J.; Kryukov, G.V.; Sonkin, D.; et al. The Cancer Cell Line Encyclopedia enables predictive modelling of anticancer drug sensitivity. Nature 2012, 483, 603–607. [Google Scholar] [CrossRef] [PubMed]
- Iorio, F.; Knijnenburg, T.A.; Vis, D.J.; Bignell, G.R.; Menden, M.P.; Schubert, M.; Aben, N.; Gonçalves, E.; Barthorpe, S.; Lightfoot, H.; et al. A Landscape of Pharmacogenomic Interactions in Cancer. Cell 2016, 166, 740–754. [Google Scholar] [CrossRef] [PubMed]
- Behan, F.M.; Iorio, F.; Picco, G.; Gonçalves, E.; Beaver, C.M.; Migliardi, G.; Santos, R.; Rao, Y.; Sassi, F.; Pinnelli, M.; et al. Prioritization of cancer therapeutic targets using CRISPR-Cas9 screens. Nature 2019, 568, 511–516. [Google Scholar] [CrossRef]
- Van der Meer, D.; Barthorpe, S.; Yang, W.; Lightfoot, H.; Hall, C.; Gilbert, J.; Francies, H.E.; Garnett, M.J. Cell Model Passports-a hub for clinical, genetic and functional datasets of preclinical cancer models. Nucleic Acids Res. 2019, 47, D923–D929. [Google Scholar] [CrossRef]
- Tsherniak, A.; Vazquez, F.; Montgomery, P.G.; Weir, B.A.; Kryukov, G.; Cowley, G.S.; Gill, S.; Harrington, W.F.; Pantel, S.; Krill-Burger, J.M.; et al. Defining a Cancer Dependency Map. Cell 2017, 170, 564–576.e16. [Google Scholar] [CrossRef]
- Marcotte, R.; Sayad, A.; Brown, K.R.; Sanchez-Garcia, F.; Reimand, J.; Haider, M.; Virtanen, C.; Bradner, J.E.; Bader, G.D.; Mills, G.B.; et al. Functional Genomic Landscape of Human Breast Cancer Drivers, Vulnerabilities, and Resistance. Cell 2016, 164, 293–309. [Google Scholar] [CrossRef] [PubMed]
- Fomicheva, M.; Tross, E.M.; Macara, I.G. Polarity proteins in oncogenesis. Curr. Opin. Cell Biol. 2020, 62, 26–30. [Google Scholar] [CrossRef] [PubMed]
- Stahley, S.N.; Basta, L.P.; Sharan, R.; Devenport, D. Celsr1 adhesive interactions mediate the asymmetric organization of planar polarity complexes. Elife 2021, 10, e62097. [Google Scholar] [CrossRef] [PubMed]
- Humphries, A.C.; Mlodzik, M. From instruction to output: Wnt/PCP signaling in development and cancer. Curr. Opin. Cell Biol. 2018, 51, 110–116. [Google Scholar] [CrossRef]
- Martin, E.; Girardello, R.; Dittmar, G.; Ludwig, A. New insights into the organization and regulation of the apical polarity network in mammalian epithelial cells. FEBS J. 2021, 288, 7073–7095. [Google Scholar] [CrossRef]
- How, J.Y.; Stephens, R.K.; Lim, K.Y.B.; Humbert, P.O.; Kvansakul, M. Structural basis of the human Scribble-Vangl2 association in health and disease. Biochem. J. 2021, 478, 1321–1332. [Google Scholar] [CrossRef]
- Feng, D.; Wang, J.; Yang, W.; Li, J.; Lin, X.; Zha, F.; Wang, X.; Ma, L.; Choi, N.T.; Mii, Y.; et al. Regulation of Wnt/PCP signaling through p97/VCP-KBTBD7-mediated Vangl ubiquitination and endoplasmic reticulum-associated degradation. Sci. Adv. 2021, 7, eabg2099. [Google Scholar] [CrossRef]
- Katoh, M. Canonical and non-canonical WNT signaling in cancer stem cells and their niches: Cellular heterogeneity, omics reprogramming, targeted therapy and tumor plasticity (Review). Int. J. Oncol. 2017, 51, 1357–1369. [Google Scholar] [CrossRef]
- Bernatik, O.; Paclikova, P.; Sri Ganji, R.; Bryja, V. Activity of Smurf2 Ubiquitin Ligase Is Regulated by the Wnt Pathway Protein Dishevelled. Cells 2020, 9, 1147. [Google Scholar] [CrossRef]
- Guo, Y.; Arciero, C.A.; Jiang, R.; Behera, M.; Peng, L.; Li, X. Different Breast Cancer Subtypes Show Different Metastatic Patterns: A Study from a Large Public Database. Asian Pac. J. Cancer Prev. 2020, 21, 3587–3593. [Google Scholar] [CrossRef]
- Van Mechelen, M.; Van Herck, A.; Punie, K.; Nevelsteen, I.; Smeets, A.; Neven, P.; Weltens, C.; Han, S.; Vanderstichele, A.; Floris, G.; et al. Behavior of metastatic breast cancer according to subtype. Breast Cancer Res. Treat. 2020, 181, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Jin, J.; Liang, T.; Feng, X.H. To Ub or not to Ub: A regulatory question in TGF-β signaling. Trends Biochem. Sci. 2022, 47, 1059–1072. [Google Scholar] [CrossRef] [PubMed]
- Stephens, R.; Lim, K.; Portela, M.; Kvansakul, M.; Humbert, P.O.; Richardson, H.E. The Scribble Cell Polarity Module in the Regulation of Cell Signaling in Tissue Development and Tumorigenesis. J. Mol. Biol. 2018, 430, 3585–3612. [Google Scholar] [CrossRef]
- Feigin, M.E.; Akshinthala, S.D.; Araki, K.; Rosenberg, A.Z.; Muthuswamy, L.B.; Martin, B.; Lehmann, B.D.; Berman, H.K.; Pietenpol, J.A.; Cardiff, R.D.; et al. Mislocalization of the cell polarity protein scribble promotes mammary tumorigenesis and is associated with basal breast cancer. Cancer Res. 2014, 74, 3180–3194. [Google Scholar] [CrossRef]
- Lin, W.H.; Asmann, Y.W.; Anastasiadis, P.Z. Expression of polarity genes in human cancer. Cancer Inform. 2015, 14 (Suppl. 3), 15–28. [Google Scholar] [CrossRef] [PubMed]
- Sakakibara, J.; Sakakibara, M.; Shiina, N.; Fujimori, T.; Okubo, Y.; Fujisaki, K.; Nagashima, T.; Sangai, T.; Nakatani, Y.; Miyazaki, M. Expression of cell polarity protein scribble differently affects prognosis in primary tumor and lymph node metastasis of breast cancer patients. Breast Cancer 2017, 24, 393–399. [Google Scholar] [CrossRef]
- Krishnamurthy, N.; Kurzrock, R. Targeting the Wnt/beta-catenin pathway in cancer: Update on effectors and inhibitors. Cancer Treat. Rev. 2018, 62, 50–60. [Google Scholar] [CrossRef]
- Torres, V.I.; Godoy, J.A.; Inestrosa, N.C. Modulating Wnt signaling at the root: Porcupine and Wnt acylation. Pharmacol. Ther. 2019, 198, 34–45. [Google Scholar] [CrossRef]
- Riffell, J.L.; Lord, C.J.; Ashworth, A. Tankyrase-targeted therapeutics: Expanding opportunities in the PARP family. Nat. Rev. Drug Discov. 2012, 11, 923–936. [Google Scholar] [CrossRef]
- Liu, Y.; Qi, X.; Donnelly, L.; Elghobashi-Meinhardt, N.; Long, T.; Zhou, R.W.; Sun, Y.; Wang, B.; Li, X. Mechanisms and inhibition of Porcupine-mediated Wnt acylation. Nature 2022, 607, 816–822. [Google Scholar] [CrossRef]
- Wang, W.; Liu, P.; Lavrijsen, M.; Li, S.; Zhang, R.; Li, S.; van de Geer, W.S.; van de Werken, H.J.G.; Peppelenbosch, M.P.; Smits, R. Evaluation of AXIN1 and AXIN2 as targets of tankyrase inhibition in hepatocellular carcinoma cell lines. Sci. Rep. 2021, 11, 7470. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.; Dynek, J.N.; Smith, S. NuMA is a major acceptor of poly(ADP-ribosyl)ation by tankyrase 1 in mitosis. Biochem. J. 2005, 391 Pt 2, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, S.J.; Smith, S. Sister telomeres rendered dysfunctional by persistent cohesion are fused by NHEJ. J. Cell Biol. 2009, 184, 515–526. [Google Scholar] [CrossRef] [PubMed]
- Diamond, J.R.; Becerra, C.; Richards, D.; Mita, A.; Osborne, C.; O’Shaughnessy, J.; Zhang, C.; Henner, R.; Kapoun, A.M.; Xu, L.; et al. Phase Ib clinical trial of the anti-frizzled antibody vantictumab (OMP-18R5) plus paclitaxel in patients with locally advanced or metastatic HER2-negative breast cancer. Breast Cancer Res. Treat. 2020, 184, 53–62. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Function | Mutations (%) | Missense | Nonsense | FSs | Amplifications (%) | Deletions (%) |
|---|---|---|---|---|---|---|---|
| WNT5A | Ligands of the β-catenin independent WNT pathway | 0 | 0 | 4 (0.4) | |||
| WNT11 | 1 (0.1) | 1 | 0 | 0 | 56 (5.2) | 2 (0.2) | |
| FZD7 | Receptor of the β-catenin independent WNT pathway | 2 (0.2) | 1 | 1 | 0 | 2 (0.2) | 5 (0.5) |
| VANGL1 | Membrane tetraspanin | 7 (0.7) | 6 | 0 | 1 | 6 (0.6) | 3 (0.3) |
| VANGL2 | Membrane tetraspanin | 8 (0.8) | 8 | 0 | 0 | 94 (8.8) | 0 |
| PRICKLE1 | Homologous scaffold proteins interacting with VANGL proteins and negative regulating DVL | 7 (0.7) | 3 | 2 | 2 | 10 (0.9) | 0 |
| PRICKLE2 | 6 (0.6) | 4 | 2 | 0 | 0 | 6 (0.6) | |
| PRICKLE3 | 5 (0.5) | 0 | 0 | 0 | 10 (0.9) | 1 (0.1) | |
| PRICKLE4 | 4 (0.4) | 3 | 0 | 0 | 18 (1.7) | 3 (0.3) | |
| DVL1 | Scaffold proteins interacting with FZD receptors of the WNT/β-catenin-dependent and -independent pathways | 3 (0.3) | 3 | 0 | 0 | 3 (0.3) | 10 (0.9) |
| DVL2 | 3 (0.3) | 2 | 0 | 1 | 0 | 7 (0.7) | |
| DVL3 | 3 (0.3) | 3 | 0 | 0 | 22 (2.1) | 0 | |
| DAAM1 | Formin homology domain scaffolding proteins interacting with DVL and RhoA | 10 (0.9) | 8 | 0 | 1 | 8 (0.7) | 0 |
| DAAM2 | 12 (1.1) | 9 | 0 | 1 | 11 (1) | 2 (0.2) | |
| SCRIB | Scaffold protein with PDZ domain | 8 (0.8) | 5 | 0 | 3 | 110 (10.3) | 3 (0.3) |
| RNF41 | Ubiquitin ligase, negative regulator of DVL | 5 (0.5) | 3 | 1 | 1 | 2 (0.2) | 0 |
| NOS1AP | Adaptor protein interacting with SCRIB involved in migration | 5 (0.5) | 5 | 0 | 0 | 97 (9.1) | 0 |
| RHOA | Small GTPase involved in actin cytoskeleton reorganization | 8 (0.8) | 6 | 0 | 2 | 1 (0.1) | 5 (0.5) |
| ROCK1 | Serine threonine kinase bound to GTP-bound RhoA | 9 (0.8) | 7 | 1 | 1 | 3 (0.3) | 2 (0.2) |
| SQSTM1 | Adaptor protein involved in endocytosis of VANGL proteins | 5 (0.5) | 3 | 0 | 1 | 10 (0.9) | 2 (0.2) |
| MINK1 | Kinase phosphorylating PRICKLE proteins promoting asymmetric localization | 5 (0.5) | 4 | 0 | 0 | 2 (0.2) | 4 (0.4) |
| SMURF2 | Ubiquitin ligase, negative regulator of PRICKLE | 6 (0.6) | 4 | 1 | 1 | 66 (6.2) | 0 |
| Gene | Mutations | Amplifications | Deletions |
|---|---|---|---|
| SCRIB | HCC1395, HCC1569, KPL1, MCF7, MDA-MB-361 | BT483, HCC1187, HCC1806 BT549, HCC1395, HCC1937, CAMA1, HCC1500, HCC2157, EFM19, HCC1569, HCC2218, HCC1143, HCC1599, HCC70, HDQ-P1, HS-578-T, MCF7, MDA-MB-436, UACC893, ZR7530 | HCC1428 |
| VANGL2 | HCC1569, HS-343-T | BT483, CAL148, CAMA1, DU4475, HCC2157, HCC38, MDA-MB-436, ZR7530 | MCF7 |
| NOS1AP | HCC1143, MDA-MB-415 | BT483, CAL148, CAMA1, DU4475, HCC38, MDA-MB-436, ZR7530 | MCF7 |
| Cell Line | DepMap ID | Type | Ploidy | Mutations/Mb |
|---|---|---|---|---|
| CAMA1 | ACH-000783 | ER+/HER2−, Luminal | 1.93 | 47.03 |
| BT-483 | ACH-000818 | ER+/HER2−, Luminal | 3.84 | 46.45 |
| MDA-MB-436 | ACH-000573 | ER−/HER2− Basal B | 2.97 | 32.76 |
| ZR-75-30 | ACH-000828 | ER +/HER2+ amplified | 3.74 | 45.32 |
| Cell Line | Target | Dataset | N with/n without | Mean IC50 with (µM) | Mean IC50 without (µM) | p |
|---|---|---|---|---|---|---|
| IWP-2 | PORCN | GDSC2 | 7/34 | 17.03 | 17.9 | 0.79 |
| LGK974 | PORCN | GDSC1 | 7/41 | 7.08 | 6.82 | 1 |
| GDSC2 | 9/39 | 44.97 | 52.21 | 0.69 | ||
| Wnt-C59 | PORCN | GDSC2 | 7/34 | 45.2 | 70.67 | 0.26 |
| GDSC1 | 7/41 | 6.78 | 6.63 | 0.88 | ||
| AZ6102 | Tankyrases 1 and 2 | GDSC2 | 9/35 | 20.04 | 15.6 | 0.3 |
| MN-64 | Tankyrases 1 and 2 | GDSC2 | 7/34 | 62.19 | 90.47 | 0.34 |
| WIKI4 | Tankyrases 1 and 2 | GDSC2 | 9/35 | 68.49 | 38.54 | 0.36 |
| XAV939 | Tankyrases 1 and 2 | GDSC2 | 7/34 | 106.47 | 85.35 | 0.24 |
| GDSC1 | 9/38 | 56.76 | 33.99 | 0.17 |
| Cell Line | CRISPR Preferential Essential Genes | RNAi |
|---|---|---|
| CAMA1 | EIF1AX, YPEL5, TRPS1, UBE2H, PCYT1A, PSMB5, ARIH1, ACTB, NAMPT and GPX4 | UBC, MAP3K7, CDK4, FOXA1, TFDP1, CIT, LRRC46, STRAP, MAGOHB and TRPS1 |
| BT-483 | NA | PREB, PTP4A1, NACAD, PITPNM1, HIGD1A, NEK11, POLM, NDC80, BTN3A1 and SNW1 |
| MDA-MB-436 | TYMS, INTS6, VPS4A, GPATCH1, DNAJC9, IER3IP1, WDR48, LEMD2, XRCC1 and ACO2 | HSPA8, PPIL2, NSF, DHX8, CDC27, VPS4A, COPG1, TOPBP1, PCNA and WDR70 |
| ZR-75-30 | NA | MYL12B, UBA6, MED1, PIK3CA, CDC37, FOXA1, SRPRA, FSTL4, ERBB2 and ASCC3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Voutsadakis, I.A. Molecular Alterations and Putative Therapeutic Targeting of Planar Cell Polarity Proteins in Breast Cancer. J. Clin. Med. 2023, 12, 411. https://doi.org/10.3390/jcm12020411
Voutsadakis IA. Molecular Alterations and Putative Therapeutic Targeting of Planar Cell Polarity Proteins in Breast Cancer. Journal of Clinical Medicine. 2023; 12(2):411. https://doi.org/10.3390/jcm12020411
Chicago/Turabian StyleVoutsadakis, Ioannis A. 2023. "Molecular Alterations and Putative Therapeutic Targeting of Planar Cell Polarity Proteins in Breast Cancer" Journal of Clinical Medicine 12, no. 2: 411. https://doi.org/10.3390/jcm12020411
APA StyleVoutsadakis, I. A. (2023). Molecular Alterations and Putative Therapeutic Targeting of Planar Cell Polarity Proteins in Breast Cancer. Journal of Clinical Medicine, 12(2), 411. https://doi.org/10.3390/jcm12020411