
How a Subclinical Unilateral Vestibular Signal Improves Binocular Vision

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
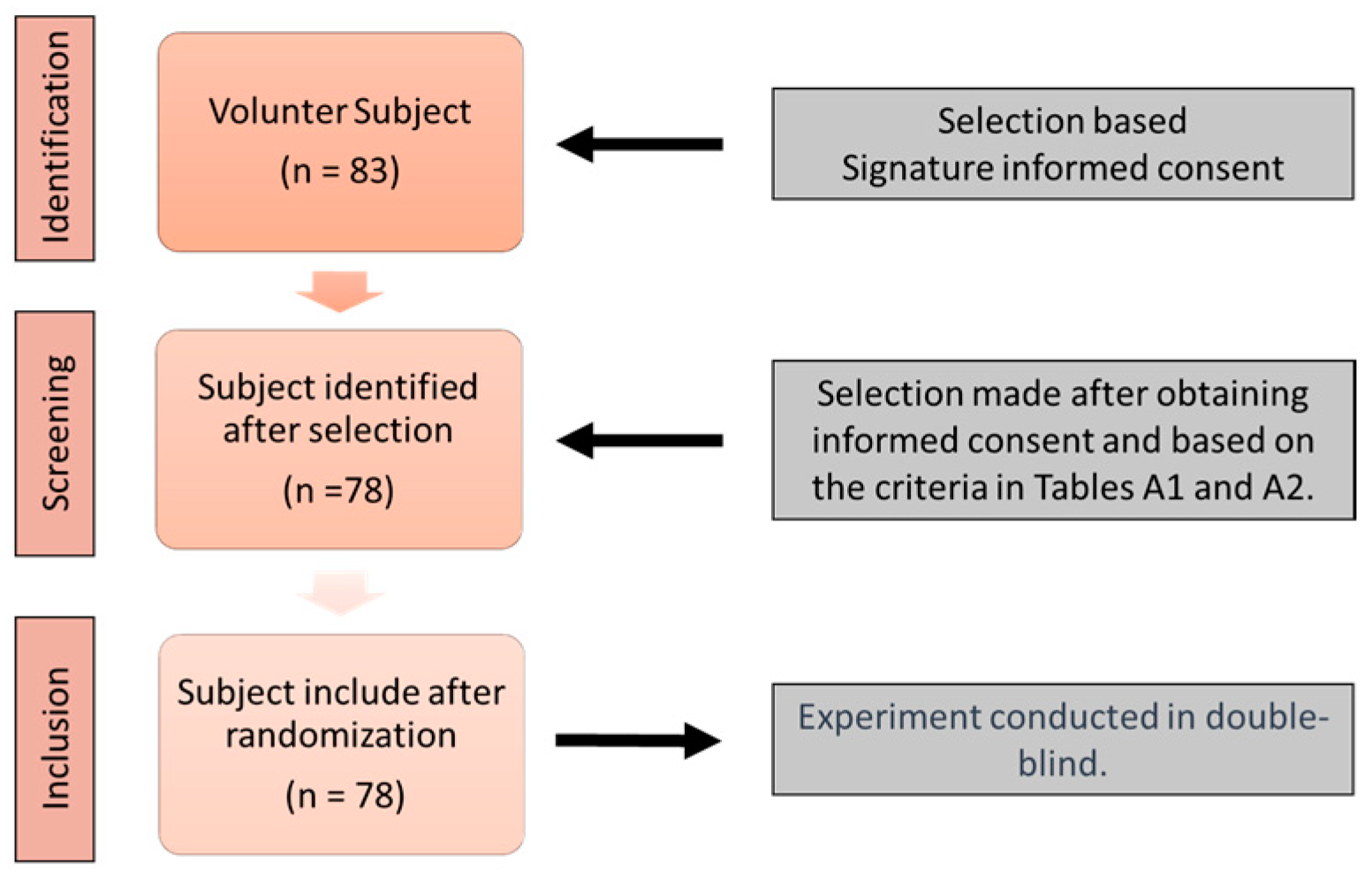
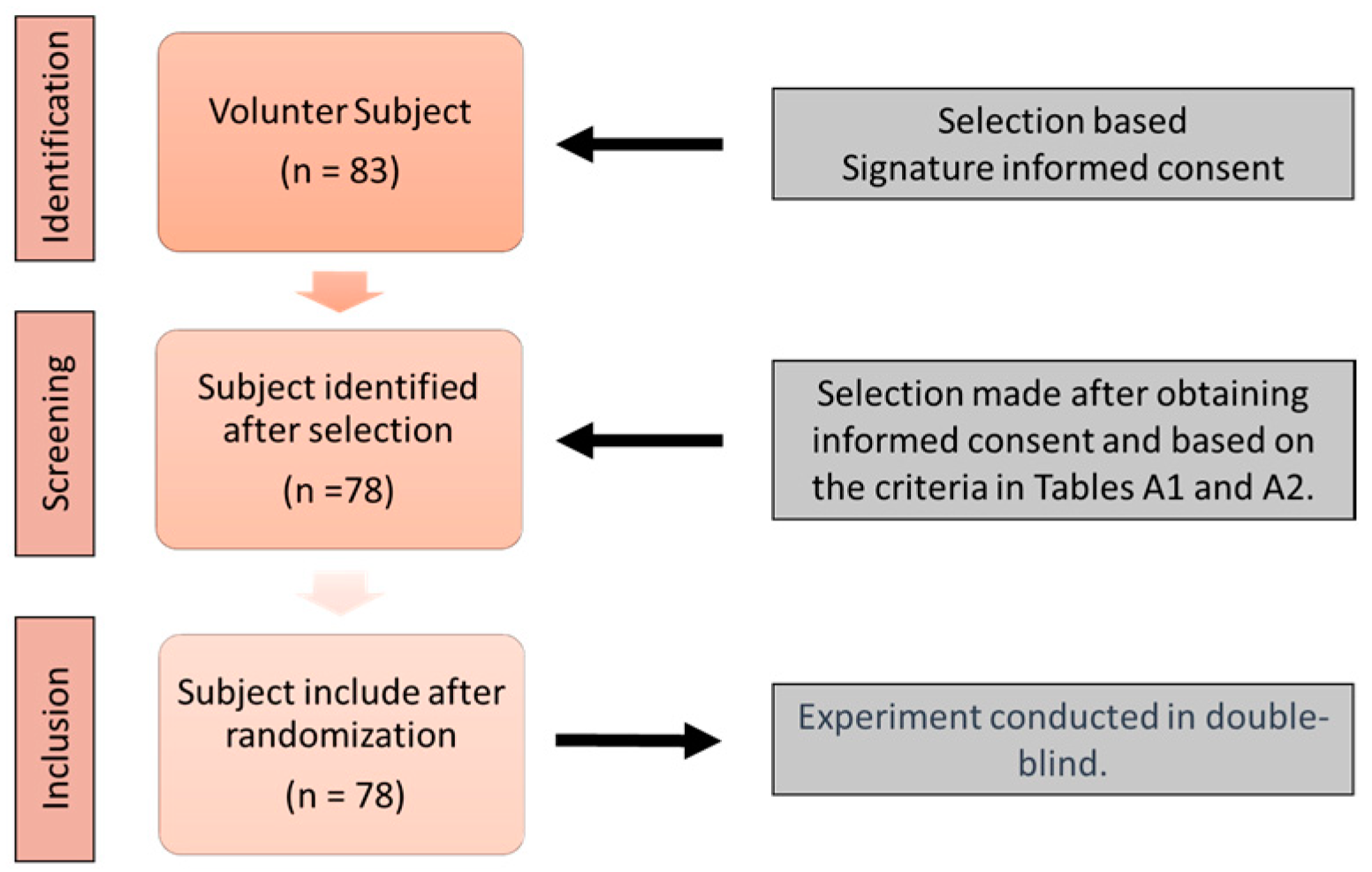
2.1. Study Design
2.2. Experimental Protocol
2.3. Statistical Analysis
3. Results
3.1. Data Description
3.2. Indicators Evolution According to the Stimulation Factor
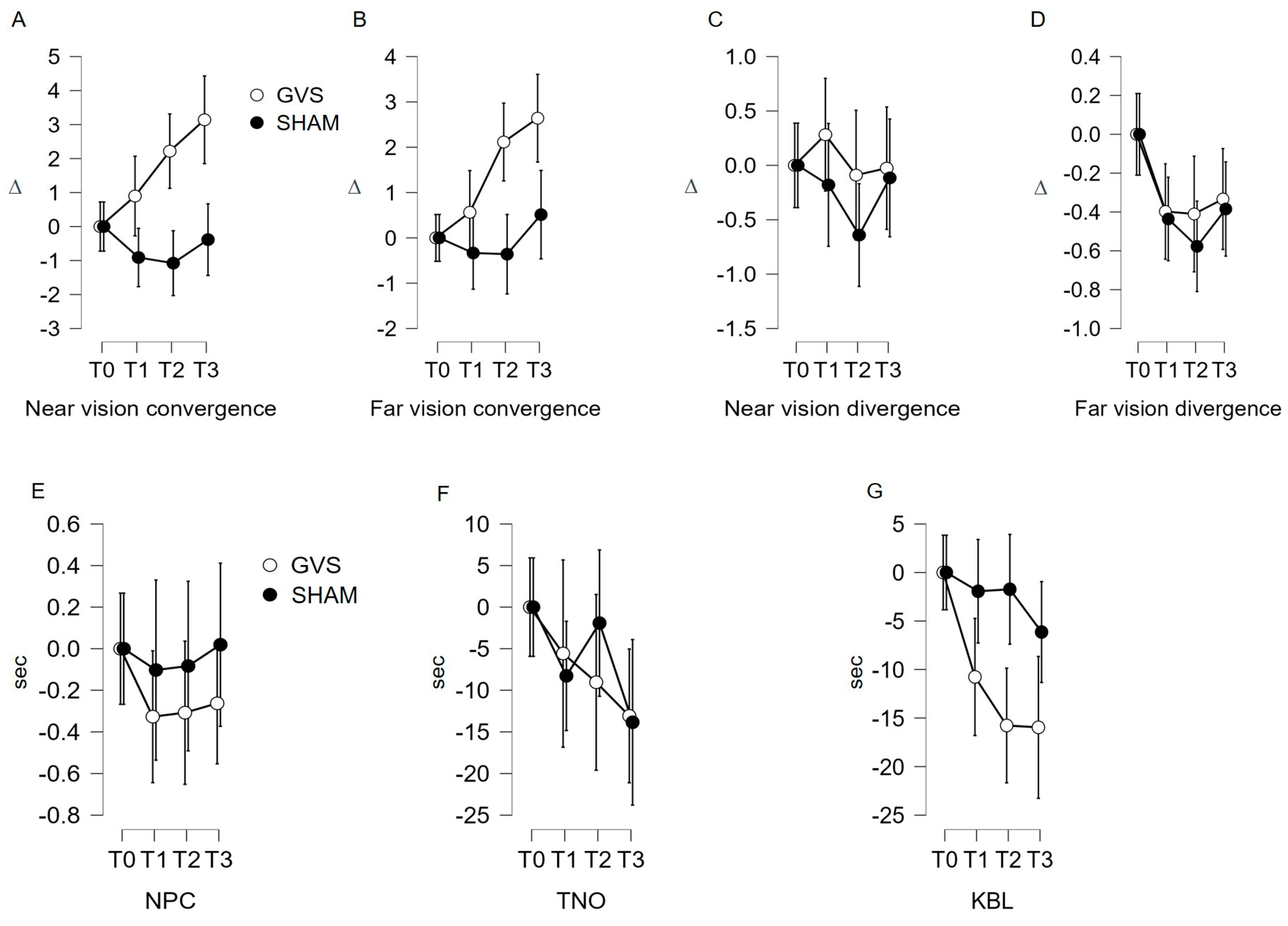
3.2.1. Near Convergence Indicator (C′; Figure 2A; Table 1)
3.2.2. Far Convergence Indicator (C; Figure 2B; Table 1)
3.2.3. Near Divergence Indicator (D′)
3.2.4. Far Divergence Indicator (D, Figure 2D; Table 1)
3.2.5. Near Point of Convergence (NPC) and Stereopsis with Graded Circle (TNO) Indicators
3.2.6. Kratsa–Barron–Laraudogoitia Indicator (KBL; Figure 2G; Table 1)
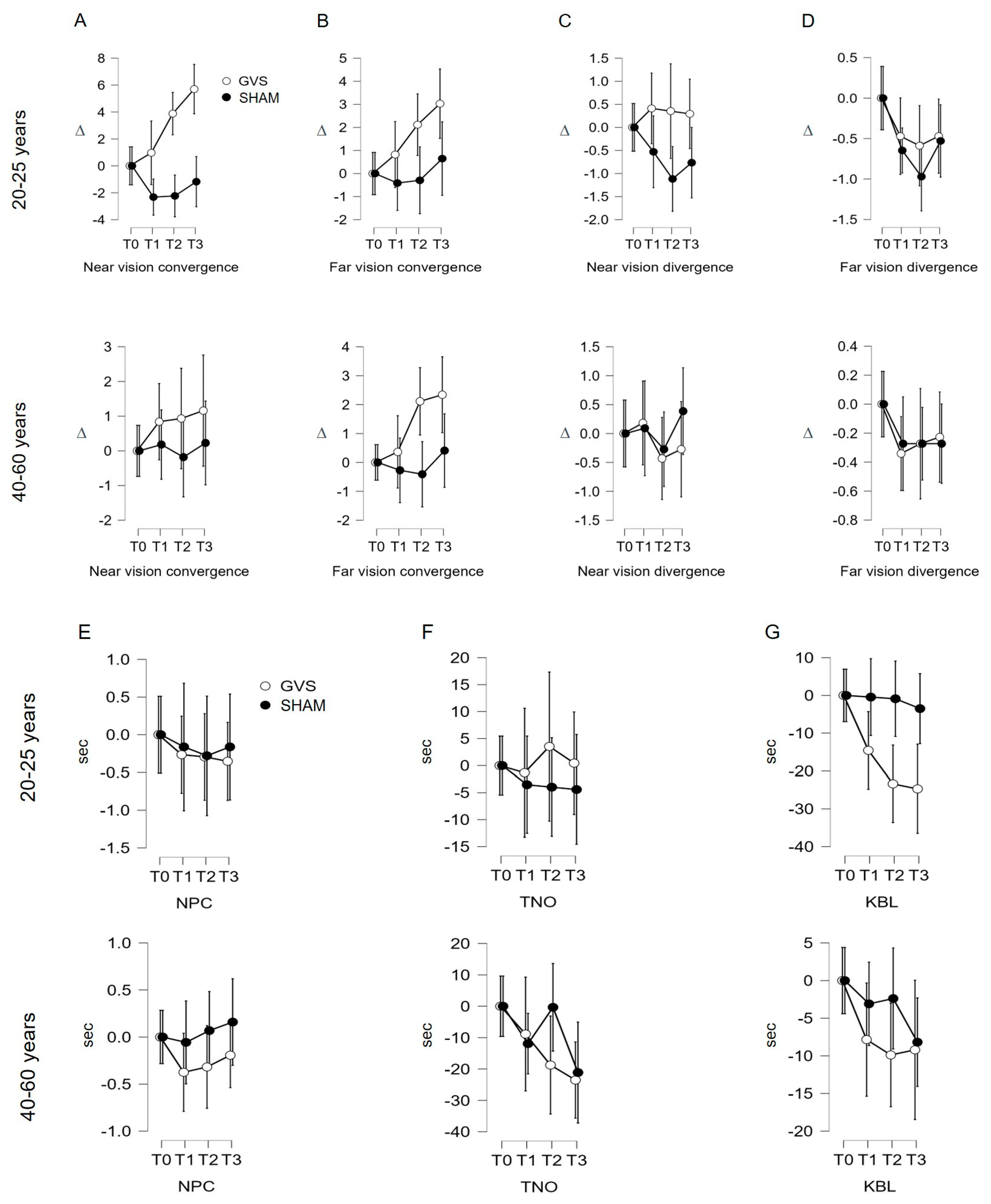
3.3. Evolution of Follow-Up Indicators in Both Age Groups
3.3.1. Near Convergence Indicator (C′)
3.3.2. Analysis of Indicators C, D′, D, NPC, TNO, KBL
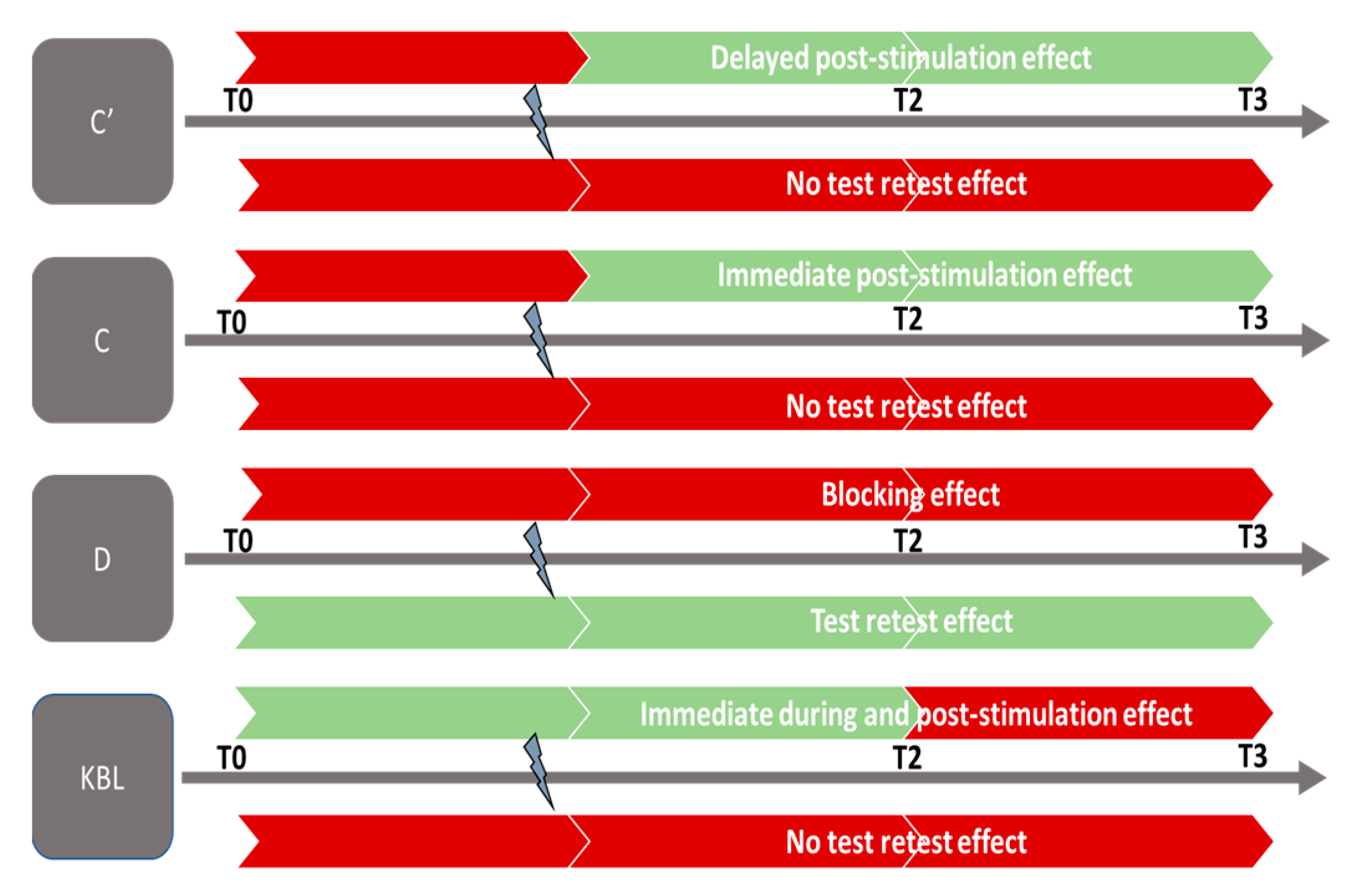
4. Discussion
5. Perspectives and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items |
|---|
| Visual acuity measurement (at 5 m: Monoyer chart and at 40 cm: Parinaud chart). |
| Phoric deviation assessment using the cover test (at 5 m and 40 cm) with horizontal and vertical prism bars. |
| Evaluation of ocular motility and conjugate eye movements using a fixation target. |
| Phoric deviation measurement using a Maddox rod (at 5 m and 40 cm). |
| Measurement of the near point of convergence using a Mawas ruler. Assessment of convergence and divergence fusional amplitudes (at 5 m and 40 cm). Stereo vision examination using the TNO test (at 40 cm) and Laroudoux and Kratz stereograms (at 2.5 m). |
| Items |
|---|
| Heterotropia |
| Abnormal retinal correspondence (ARC) |
| Visual acuity less than 10/10 in both eyes |
| Abnormal fixation (nystagmus) |
| Abnormal eye movements (paresis, paralysis, alphabetic syndrome) Positive diagnosis of an ocular pathology Positive diagnosis of a general pathology that can impact oculomotor function Positive diagnosis of a neurological or neurodegenerative pathology Positive diagnosis of a vestibular pathology Regular presence of vertigo or motion sickness Ongoing orthodontic and/or orthopedic treatment |
| Items | Description |
|---|---|
| Far convergence at 5 meters: C | The subject fixates the light and sees only one, without neutralization. The horizontal prism bar is placed at the base-in position in front of one eye. The operator increases the power of the prism until the subject can no longer fuse. The measurement of convergence is given by the strongest prism that could be compensated, indicated as C + value in diopters (∆). Norms range from 8 to 10 ∆. |
| Near convergence at 40 cm: C′ | Same procedure. The measurement of convergence is given by the strongest prism that could be compensated, indicated as C’ ∆. Norms range from 30 to 40 ∆. |
| Far divergence at 5 m: D | Same procedure, but the horizontal prism bar is placed base-out in front of one eye: the measurement of divergence is given by the strongest prism that could be compensated, indicated as D ∆. Norms range from 2 to 4 ∆. |
| Near divergence at 40 cm: D′ | Same procedure, but the horizontal prism bar is placed base-out in front of one eye: the measurement of divergence is given by the strongest prism that could be compensated, indicated as D′ ∆. Norms range from 6 to 8 ∆. |
| Near Point of Convergence: NPC | An object is brought closer until one eye deviates outward, and the NPC (near point of convergence) is measured using a ruler. Its normal value is around 8 to 10 cm from the orbital rim. It is trainable and can be modified voluntarily. |
| Far Stereoscopic Acuity at 2.5 m: Kratsa-Barron-Laraudogoitia (KBL) | It consists of random red–green dot pattern and is performed using red and green filters. The stereoscopic acuity is measured at 250 s of arc at 5 m and 500 s of arc at 2.50 m. At 5 m, it is a central test, while closer distances involve peripheral fusion. Norms: stereoscopic vision less than 100 s of arc is considered good. |
| Near Stereoscopic Acuity at 40 cm: Stereopsis with Graded Circle (TNO). | The TNO stereotest consists of six plates (ranging from 480 to 15 s of arc) of anaglyph random-dot stereograms. They should be viewed through red–green glasses. This test measures very fine stereoscopic acuity. Norms: The average stereoscopic acuity in the population is 20 s of arc. For individuals over forty years old, the average value is 58 s of arc. |
References
- Tarnutzer, A.A.; Berkowitz, A.L.; Robinson, K.A.; Hsieh, Y.-H.; Newman-Toker, D.E. Does My Dizzy Patient Have a Stroke? A Systematic Review of Bedside Diagnosis in Acute Vestibular Syndrome. CMAJ 2011, 183, E571–E592. [Google Scholar] [CrossRef] [PubMed]
- Edlow, J.A.; Gurley, K.L.; Newman-Toker, D.E. A new diagnostic approach to the adult patient with acute dizziness. J. Emerg. Med. 2018, 54, 469–483. [Google Scholar] [CrossRef] [PubMed]
- Newman-Toker, D.E.; Edlow, J.A. TiTrATE: A Novel Approach to Diagnosing Acute Dizziness and Vertigo. Neurol. Clin. 2015, 33, 577–599. [Google Scholar] [CrossRef] [PubMed]
- Ahsan, S.F.; Syamal, M.N.; Yaremchuk, K.; Peterson, E.; Seidman, M. The Costs and Utility of Imaging in Evaluating Dizzy Patients in the Emergency Room. Laryngoscope 2013, 123, 2250–2253. [Google Scholar] [CrossRef]
- Hülse, R.; Biesdorf, A.; Hörmann, K.; Stuck, B.; Erhart, M.; Hülse, M.; Wenzel, A. Peripheral Vestibular Disorders: An Epidemiologic Survey in 70 Million Individuals. Otol. Neurotol. 2019, 40, 88–95. [Google Scholar] [CrossRef]
- Laurens, J.; Awai, L.; Bockisch, C.J.; Hegemann, S.; van Hedel, H.J.A.; Dietz, V.; Straumann, D. Visual Contribution to Postural Stability: Interaction between Target Fixation or Tracking and Static or Dynamic Large-Field Stimulus. Gait Posture 2010, 31, 37–41. [Google Scholar] [CrossRef]
- Angelaki, D.E.; Hess, B.J. Self-motion-induced eye movements: Effects on visual acuity and navigation. Nat. Rev. Neurosci. 2005, 612, 966–976. [Google Scholar] [CrossRef]
- Krishnan, V.; Aruin, A.S. Postural Control in Response to a Perturbation: Role of Vision and Additional Support. Exp. Brain Res. 2011, 212, 385–397. [Google Scholar] [CrossRef]
- Ruf-Bächtiger, L. [Visual perception and its disorders]. Schweiz Rundsch Med. Prax 1989, 78, 1313–1318. [Google Scholar]
- Merfeld, D.M. Signal Detection Theory and Vestibular Thresholds: I. Basic Theory and Practical Considerations. Exp. Brain Res. 2011, 210, 389–405. [Google Scholar] [CrossRef]
- Colebatch, J.G.; Rosengren, S.M. Investigating Short Latency Subcortical Vestibular Projections in Humans: What Have We Learned? J. Neurophysiol. 2019, 122, 2000–2015. [Google Scholar] [CrossRef]
- Diaz-Artiles, A.; Karmali, F. Vestibular Precision at the Level of Perception, Eye Movements, Posture, and Neurons. Neuroscience 2021, 468, 282–320. [Google Scholar] [CrossRef] [PubMed]
- Bittar, R.S.M.; Mezzalira, R.; Ramos, A.C.M.; Risso, G.H.; Real, D.M.; Grasel, S.S. Vestibular recruitment: New application for an old concept. Braz. J. Otorhinolaryngol. 2022, 88, S91–S96. [Google Scholar] [CrossRef]
- Cohen, R.A. Neural Mechanisms of Attention. In The Neuropsychology of Attention; Springer: Boston, MA, USA, 2013; pp. 211–264. [Google Scholar] [CrossRef]
- Grantham, D.W. Detection and Discrimination of Simulated Motion of Auditory Targets in the Horizontal Plane. J. Acoust. Soc. Am. 1986, 79, 1939–1949. [Google Scholar] [CrossRef] [PubMed]
- Tilikete, C.; Vighetto, A. Expertise clinique et fonctionnelle du nerf vestibulaire. Neurochirurgie 2009, 55, 158–161. [Google Scholar] [CrossRef] [PubMed]
- Dai, C.; Fridman, G.Y.; Chiang, B.; Davidovics, N.S.; Melvin, T.-A.; Cullen, K.E.; Della Santina, C.C. Cross-Axis Adaptation Improves 3D Vestibulo-Ocular Reflex Alignment during Chronic Stimulation via a Head-Mounted Multichannel Vestibular Prosthesis. Exp. Brain Res. 2011, 210, 595–606. [Google Scholar] [CrossRef]
- Fridman, G.Y.; Della Santina, C.C. Progress toward Development of a Multichannel Vestibular Prosthesis for Treatment of Bilateral Vestibular Deficiency. Anat. Rec. 2012, 295, 2010–2029. [Google Scholar] [CrossRef]
- Sluydts, M.; Curthoys, I.; Vanspauwen, R.; Papsin, B.C.; Cushing, S.L.; Ramos, A.; Ramos de Miguel, A.; Borkoski Barreiro, S.; Barbara, M.; Manrique, M.; et al. Electrical Vestibular Stimulation in Humans: A Narrative Review. Audiol. Neurootol. 2020, 25, 6–24. [Google Scholar] [CrossRef]
- Wiboonsaksakul, K.P.; Roberts, D.C.; Della Santina, C.C.; Cullen, K.E. A Prosthesis Utilizing Natural Vestibular Encoding Strategies Improves Sensorimotor Performance in Monkeys. PLoS Biol. 2022, 20, e3001798. [Google Scholar] [CrossRef]
- Goldberg, J.M.; Fernández, C.; Smith, C.E. Responses of Vestibular-Nerve Afferents in the Squirrel Monkey to Externally Applied Galvanic Currents. Brain Res. 1982, 252, 156–160. [Google Scholar] [CrossRef]
- Kwan, A.; Forbes, P.A.; Mitchell, D.E.; Blouin, J.-S.; Cullen, K.E. Neural Substrates, Dynamics and Thresholds of Galvanic Vestibular Stimulation in the Behaving Primate. Nat. Commun. 2019, 10, 1904. [Google Scholar] [CrossRef]
- Reynolds, R.F.; Osler, C.J. Galvanic Vestibular Stimulation Produces Sensations of Rotation Consistent with Activation of Semicircular Canal Afferents. Front. Neurol. 2012, 3, 104. [Google Scholar] [CrossRef] [PubMed]
- Ghanim, Z.; Lamy, J.C.; Lackmy, A.; Achache, V.; Roche, N.; Pénicaud, A.; Meunier, S.; Katz, R. Effects of Galvanic Mastoid Stimulation in Seated Human Subjects. J. Appl. Physiol. 2009, 106, 893–903. [Google Scholar] [CrossRef]
- Curthoys, I.S.; MacDougall, H.G. What Galvanic Vestibular Stimulation Actually Activates. Front. Neurol. 2012, 3, 117. [Google Scholar] [CrossRef] [PubMed]
- Pires, A.P.B.d.Á.; Silva, T.R.; Torres, M.S.; Diniz, M.L.; Tavares, M.C.; Gonçalves, D.U. Galvanic Vestibular Stimulation and Its Applications: A Systematic Review. Braz. J. Otorhinolaryngol. 2022, 88, S202–S211. [Google Scholar] [CrossRef] [PubMed]
- Dlugaiczyk, J.; Gensberger, K.D.; Straka, H. Galvanic Vestibular Stimulation: From Basic Concepts to Clinical Applications. J. Neurophysiol. 2019, 121, 2237–2255. [Google Scholar] [CrossRef] [PubMed]
- De Maio, G.; Bottini, G.; Ferré, E.R. Galvanic Vestibular Stimulation Influences Risk-Taking Behaviour. Neuropsychologia 2021, 160, 107965. [Google Scholar] [CrossRef]
- Aedo-Jury, F.; Cottereau, B.R.; Celebrini, S.; Séverac Cauquil, A. Antero-Posterior vs. Lateral Vestibular Input Processing in Human Visual Cortex. Front. Integr. Neurosci. 2020, 14, 43. [Google Scholar] [CrossRef]
- Rosenfield, M.; Ciuffreda, K.J.; Ong, E.; Super, S. Vergence Adaptation and the Order of Clinical Vergence Range Testing. Optom. Vis. Sci. 1995, 72, 219–223. [Google Scholar] [CrossRef]
- Waespe, W.; Henn, V. Neuronal Activity in the Vestibular Nuclei of the Alert Monkey during Vestibular and Optokinetic Stimulation. Exp. Brain Res. 1977, 27, 523–538. [Google Scholar] [CrossRef]
- Angelaki, D.E.; Cullen, K.E. Vestibular System: The Many Facets of a Multimodal Sense. Annu. Rev. Neurosci. 2008, 31, 125–150. [Google Scholar] [CrossRef]
- Kheradmand, A.; Colpak, A.I.; Zee, D.S. Eye Movements in Vestibular Disorders. Handb. Clin. Neurol. 2016, 137, 103–117. [Google Scholar] [CrossRef] [PubMed]
- Carcaud, J.; França de Barros, F.; Idoux, E.; Eugène, D.; Reveret, L.; Moore, L.E.; Vidal, P.-P.; Beraneck, M. Long-Lasting Visuo-Vestibular Mismatch in Freely-Behaving Mice Reduces the Vestibulo-Ocular Reflex and Leads to Neural Changes in the Direct Vestibular Pathway. eNeuro 2017, 4, ENEURO.0290-16.2017. [Google Scholar] [CrossRef]
- Scudder, C.A.; Fuchs, A.F. Physiological and Behavioral Identification of Vestibular Nucleus Neurons Mediating the Horizontal Vestibuloocular Reflex in Trained Rhesus Monkeys. J. Neurophysiol. 1992, 68, 244–264. [Google Scholar] [CrossRef]
- Heermann, S. [Neuroanatomy of the Visual Pathway]. Klin Monbl Augenheilkd 2017, 234, 1327–1333. [Google Scholar] [CrossRef]
- Johnson, B.P.; Lum, J.A.G.; Rinehart, N.J.; Fielding, J. Ocular Motor Disturbances in Autism Spectrum Disorders: Systematic Review and Comprehensive Meta-Analysis. Neurosci. Biobehav. Rev. 2016, 69, 260–279. [Google Scholar] [CrossRef] [PubMed]
- Beltramo, R.; Scanziani, M. A Collicular Visual Cortex: Neocortical Space for an Ancient Midbrain Visual Structure. Science 2019, 363, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Basso, M.A.; Bickford, M.E.; Cang, J. Unraveling Circuits of Visual Perception and Cognition through the Superior Colliculus. Neuron 2021, 109, 918–937. [Google Scholar] [CrossRef]
- McDougal, D.H.; Gamlin, P.D. Autonomic Control of the Eye. Compr. Physiol. 2015, 5, 439–473. [Google Scholar] [CrossRef]
- Binda, P.; Gamlin, P.D. Renewed Attention on the Pupil Light Reflex. Trends Neurosci. 2017, 40, 455–457. [Google Scholar] [CrossRef]
- Séverac Cauquil, A.; Faldon, M.; Popov, K.; Day, B.L.; Bronstein, A.M. Short-Latency Eye Movements Evoked by Near-Threshold Galvanic Vestibular Stimulation. Exp. Brain Res. 2003, 148, 414–418. [Google Scholar] [CrossRef]
- Bense, S.; Stephan, T.; Yousry, T.A.; Brandt, T.; Dieterich, M. Multisensory Cortical Signal Increases and Decreases During Vestibular Galvanic Stimulation (FMRI). J. Neurophysiol. 2001, 85, 886–899. [Google Scholar] [CrossRef]
- Helmchen, C.; Machner, B.; Rother, M.; Spliethoff, P.; Göttlich, M.; Sprenger, A. Effects of Galvanic Vestibular Stimulation on Resting State Brain Activity in Patients with Bilateral Vestibulopathy. Hum. Brain Mapp. 2020, 41, 2527–2547. [Google Scholar] [CrossRef] [PubMed]
- Miladinović, A.; Quaia, C.; Ajčević, M.; Diplotti, L.; Cumming, B.G.; Pensiero, S.; Accardo, A. Ocular-following responses in school-age children. PLoS ONE 2022, 17, e0277443. [Google Scholar] [CrossRef] [PubMed]
- Quaia, C.; FitzGibbon, E.J.; Optican, L.M.; Cumming, B.G. Binocular Summation for Reflexive Eye Movements: A Potential Diagnostic Tool for Stereodeficiencies. Investig. Ophthalmol. Vis. Sci. 2018, 59, 5816–5822. [Google Scholar] [CrossRef] [PubMed]

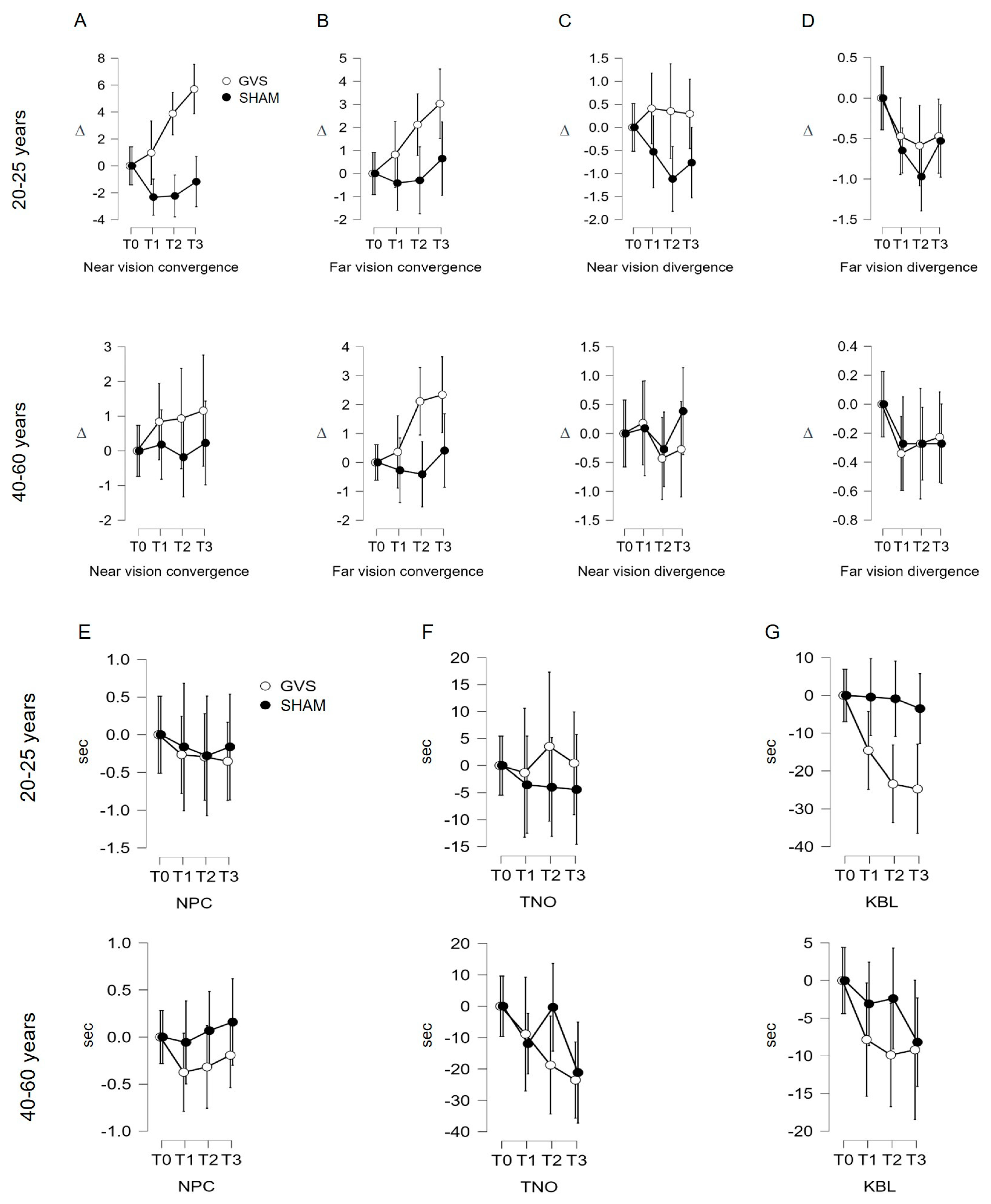
 no significant effect;
no significant effect;  significant effect.
no significant effect; significant effect.
significant effect.
no significant effect; significant effect.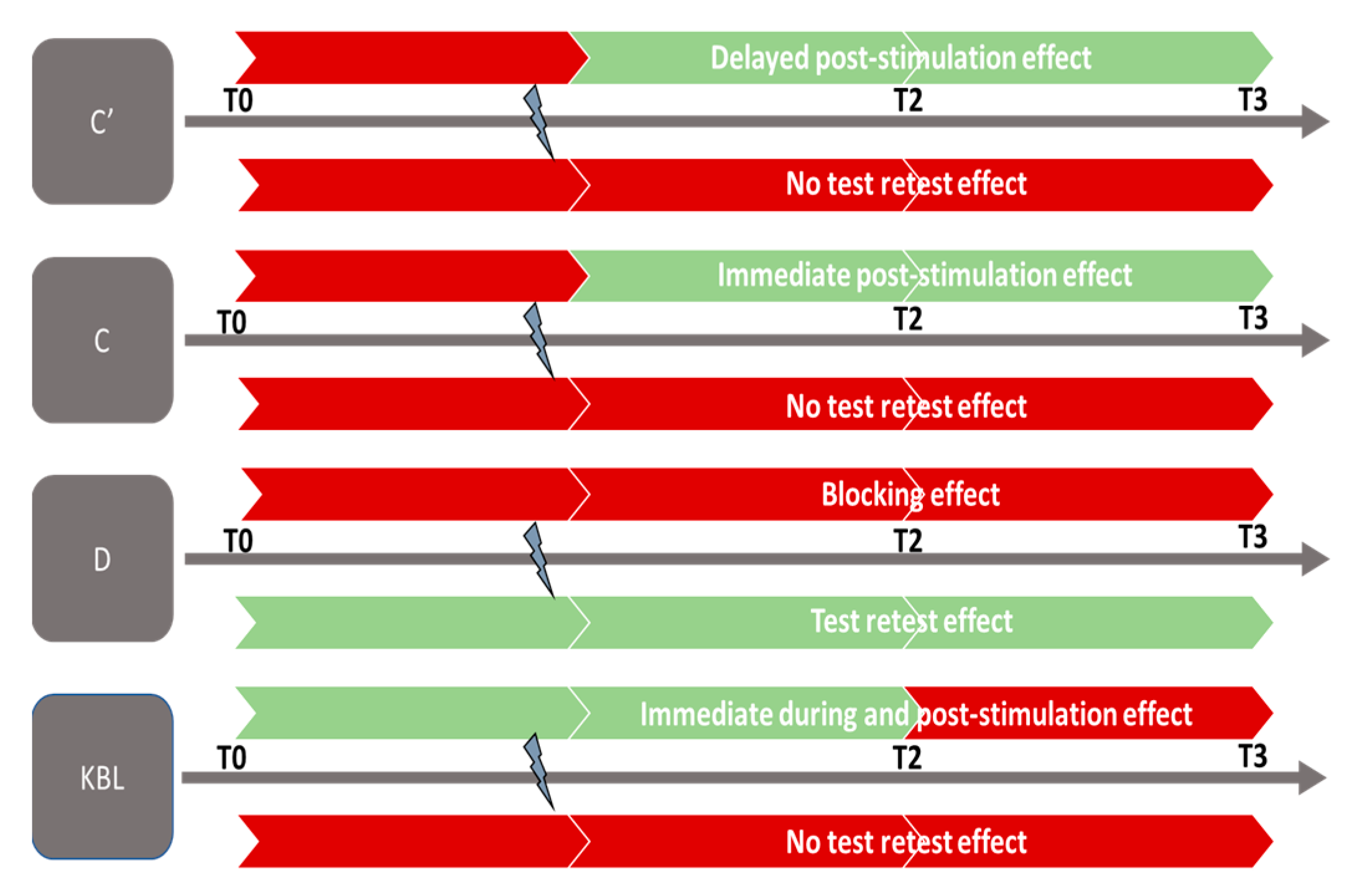
| Measurements | Stimulation | ANOVA Results | p | Significant Post Hoc Analysis |
|---|---|---|---|---|
| GVS | C′ | F (2.613, 198.569) = 10.073 | p < 0.001 | μ(T0)–μ(T2) = −2.407; p < 0.002 μ(T0)–μ(T3) = −3.432; p < 0.001 μ(T1)–μ(T3) = −2.527; p < 0.001 |
| Sham | C′ | F (2.755, 209.389) = 2.358 | p = 0.078 | |
| GVS | C | F (2.772, 210.642) = 13.027 | p < 0.001 | μ(T0)–μ(T2) = −2.116; p < 0.001 μ(T0)–μ(T3) = −2.685; p < 0.001 μ(T1)–μ(T2) = −1.522; p = 0.007 μ(T1)–μ(T3) = −2.092; p < 0.001 |
| Sham | C | F (2.492, 189.425) = 1.556 | p = 0.208 | |
| GVS | D′ | F (2.596, 197.322) = 0.460 | p = 0.683 | |
| Sham | D′ | F (2.587, 205.090) = 2.006 | p = 0.124 | |
| GVS | D | F (2.134, 162.208) = 2.942 | p = 0.052 | |
| Sham | D | F (2.699, 205.090) = 7.641 | p = 0.001 | μ(T0)–μ(T1) = 0.460; p = 0.004 μ(T0)–μ(T2) = 0.622; p < 0.001 μ(T0)–μ(T3) = 0.401; p = 0.013 |
| GVS | NPC | F (2.236, 169.964) = 2.523 | p = 0.077 | |
| Sham | NPC | F (1.270, 96.528) = 0.155 | p = 0.755 | |
| GVS | TNO | F (2.450, 186.182) = 1.281 | p = 0.282 | |
| Sham | TNO | F (1.797, 136.554) = 2.736 | p = 0.074 | |
| GVS | KBL | F (2.959, 224.850) = 9.003 | p < 0.001 | μ(T0)–μ(T1) = 11.200; p = 0.012 μ(T0)–μ(T2) = 16.634; p < 0.001 μ(T0)–μ(T3) = 16.955; p < 0.001 |
| C7 | KBL | F (2.526, 192.010) = 1.435 | p = 0.238 |
| Measurements | Stimulation | ANOVA Results | p | Significant Post Hoc Analysis |
|---|---|---|---|---|
| C′ | GVS | F (2.613, 198.569) = 6.327 | p = 0.002 | 20–25 years: T0–T2 (p = 0.005) T0–T3 (p < 0.001) T1–T3 (p < 0.001) |
| C′ | Sham | F (2.755, 209.389) = 2.251 | p = 0.089 | |
| C | GVS | F (2.772, 210.642) = 0.242 | p = 0.852 | |
| C | Sham | F (2.492, 189.425) = 0.059 | p = 0.967 | |
| D′ | GVS | F (2.596, 197.322) = 0.584 | p = 0.602 | |
| D′ | Sham | F (2.587, 196.629) = 1.360 | p = 0.258 | |
| D | GVS | F (2.134, 162.208) = 0.338 | p = 0.720 | |
| D | Sham | F (2.699, 205.090) = 2.296 | p = 0.086 | |
| NPC | GVS | F (2.236, 1169.964) = 0.351 | p = 0.728 | |
| NPC | Sham | F (1.270, 96.528) = 0.290 | p = 0.647 | |
| TNO | GVS | F (2.450, 186.182) = 1.847 | p = 0.151 | |
| TNO | Sham | F (1.797, 136.554) = 1.709 | p = 0.188 | |
| KBL | GVS | F (2.959, 224.850) = 1.779 | p = 0.153 | |
| KBL | Sham | F (2.526, 192.010) = 0.226 | p = 0.846 |
| Indicator | Between-Group Variation | Within-Group Variation in the Mean Measurements Taken at Each Time Point (T) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| T0–T3 | p | T0–T1 | p | T0–T2 | p | T0–T3 | p | T1–T2 | p | T1–T3 | p | T2–T3 | p | |
| C′ | Continuous + | ns | + | s | + | s | + | ns | + | s | + | ns | + | ns |
| shamC′ | no variation | ns | − | ns | − | ns | − | ns | − | ns | + | ns | + | ns |
| C | Continuous + | ns | + | s | + | s | + | s | + | s | + | ns | + | ns |
| shamC | no variation | ns | − | ns | − | ns | + | ns | − | ns | + | ns | + | ns |
| D′ | no variation | ns | + | ns | − | ns | − | ns | − | ns | − | ns | + | ns |
| shamD′ | no variation | ns | − | ns | − | ns | − | ns | − | ns | + | ns | + | ns |
| D | no variation | ns | − | ns | − | ns | − | ns | − | ns | + | ns | + | ns |
| shamD | Discontinuous | s | − | s | − | s | − | ns | − | ns | + | ns | + | ns |
| NPC | no variation | ns | − | ns | − | ns | − | ns | + | ns | + | ns | + | ns |
| shamNPC | no variation | ns | − | ns | − | ns | + | ns | + | ns | + | ns | + | ns |
| TNO | Continuous − | ns | − | ns | − | ns | − | ns | − | ns | − | ns | − | ns |
| shamTNO | no variation | ns | − | ns | − | ns | − | ns | + | ns | − | ns | − | ns |
| KBL | Continuous − | s | − | s | − | s | − | ns | − | ns | − | ns | − | ns |
| shamKBL | no variation | ns | − | ns | − | ns | − | ns | + | ns | − | ns | − | ns |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xavier, F.; Chouin, E.; Serin-Brackman, V.; Séverac Cauquil, A. How a Subclinical Unilateral Vestibular Signal Improves Binocular Vision. J. Clin. Med. 2023, 12, 5847. https://doi.org/10.3390/jcm12185847
Xavier F, Chouin E, Serin-Brackman V, Séverac Cauquil A. How a Subclinical Unilateral Vestibular Signal Improves Binocular Vision. Journal of Clinical Medicine. 2023; 12(18):5847. https://doi.org/10.3390/jcm12185847
Chicago/Turabian StyleXavier, Frédéric, Emmanuelle Chouin, Véronique Serin-Brackman, and Alexandra Séverac Cauquil. 2023. "How a Subclinical Unilateral Vestibular Signal Improves Binocular Vision" Journal of Clinical Medicine 12, no. 18: 5847. https://doi.org/10.3390/jcm12185847
APA StyleXavier, F., Chouin, E., Serin-Brackman, V., & Séverac Cauquil, A. (2023). How a Subclinical Unilateral Vestibular Signal Improves Binocular Vision. Journal of Clinical Medicine, 12(18), 5847. https://doi.org/10.3390/jcm12185847