
The Importance of the Transcription Factor Foxp3 in the Development of Primary Immunodeficiencies


Abstract
1. Introduction
2. Molecular Characterization of the Foxp3 Protein
3. The Occurrence, Interactions, and Importance of the Foxp3 Protein in the Human Body
3.1. The Occurrence of Foxp3 Protein on Cells of the Immune System and Its Functions in the Human Body
3.2. Interactions of Foxp3 Protein with Proteins and Transcription Factors
3.3. The Importance of Post-Transcriptional and Post-Translational Modifications in the Function of the Foxp3 Protein

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of the Enzyme | Abbreviation | Functions | Reference |
|---|---|---|---|
| Histone acetyltransferase KAT5 | TIP60 | Performs histone acetylation in the nucleosome which changes the binding to DNA. Acetylation neutralizes the positive charge on the histones, reducing the binding affinity of negatively charged DNA. This in turn reduces the steric hindrance of DNA and increases the interaction of transcription factors and other proteins. The three key functions of KAT5 are its ability to regulate transcription, DNA repair, and apoptosis. | [30,120] |
| Histone acetyltransferase | p300 | Acts as a histone acetyltransferase that regulates transcription through chromatin remodeling and is important in cell proliferation and differentiation. It mediates the regulation of the cAMP gene, binding specifically to the phosphorylated CREB protein, and also contains a bromodomain which is involved in IL6 signaling. | [106,121] |
| Histone deacetylase 6 | HDAC6 | This enzyme is located in the cytoplasm where it is responsible for the regulation of acetylation of α-tubulin, HSP90, or glucocorticoid receptors. Upon activation of Treg cells, this enzyme migrates to the cell nucleus where it participates in the regulation of the acetylation level of the Foxp3 protein. Pharmacological inhibition or the use of a genetic knockout of the HDAC6 gene have been shown to increase the level of acetylation of both Foxp3 and Hsp90 proteins which results in an increase in the immunosuppressive activity of Treg cells. | [107,108,122] |
| Histone deacetylase 7 | HDAC7 | HDAC7 has been shown to have low intrinsic deacetylase activity and studies have demonstrated that HDAC7 may have a variety of alternative developmentary, proliferative, and inflammatory functions. | [101,102,123] |
| Histone deacetylase 9 | HDAC9 | Represses the activity of MEF2 by recruiting multi-component complexes containing CtBP and HDAC. May play a role in the process of hematopoiesis. | [108,124] |
| Histone deacetylase 10 | HDAC10 | From studies performed in a mouse model, HDAC10 deletion did not adversely affect the health of mice that retained normal CD4+ and CD8+ T cell function. However, HDAC10−/− Treg showed enhanced suppressive function both in vitro and in vivo. In addition, HDAC10−/− mice that received a heart transplant with a completely mismatched MHC became more tolerant and showed longer allograft survival. | [101] |
| NAD-dependent sirtuin-1 deacetylase | SIRT1 | SIRT1 deacetylates and thus inactivates the p53 protein. SIRT1 also stimulates autophagy by preventing the acetylation of proteins (via deacetylation) required for autophagy, as demonstrated in cultured cells and embryonic and neonatal tissues. This feature provides a link between sirtuin expression and the cellular response to nutrient constraints due to caloric restriction. SIRT1 inhibits NF-κB regulated gene expression by deacetylating the RelA/p65 subunit of NF-κB in lysine 310. SIRT1 plays a role in activating T17 helper cells that contribute to autoimmune disease. | [125,126] |
3.4. Importance of T Cell Metabolic Factors and the Level of Foxp3 Expression
3.5. Regulation of Foxp3 Protein Expression as a Potential Therapeutic Strategy for Autoimmune and Neoplastic Diseases
4. The Role of Foxp3 Protein in the Development of PIDs
4.1. The Role of Mutations within the Foxp3 Protein in PID Development
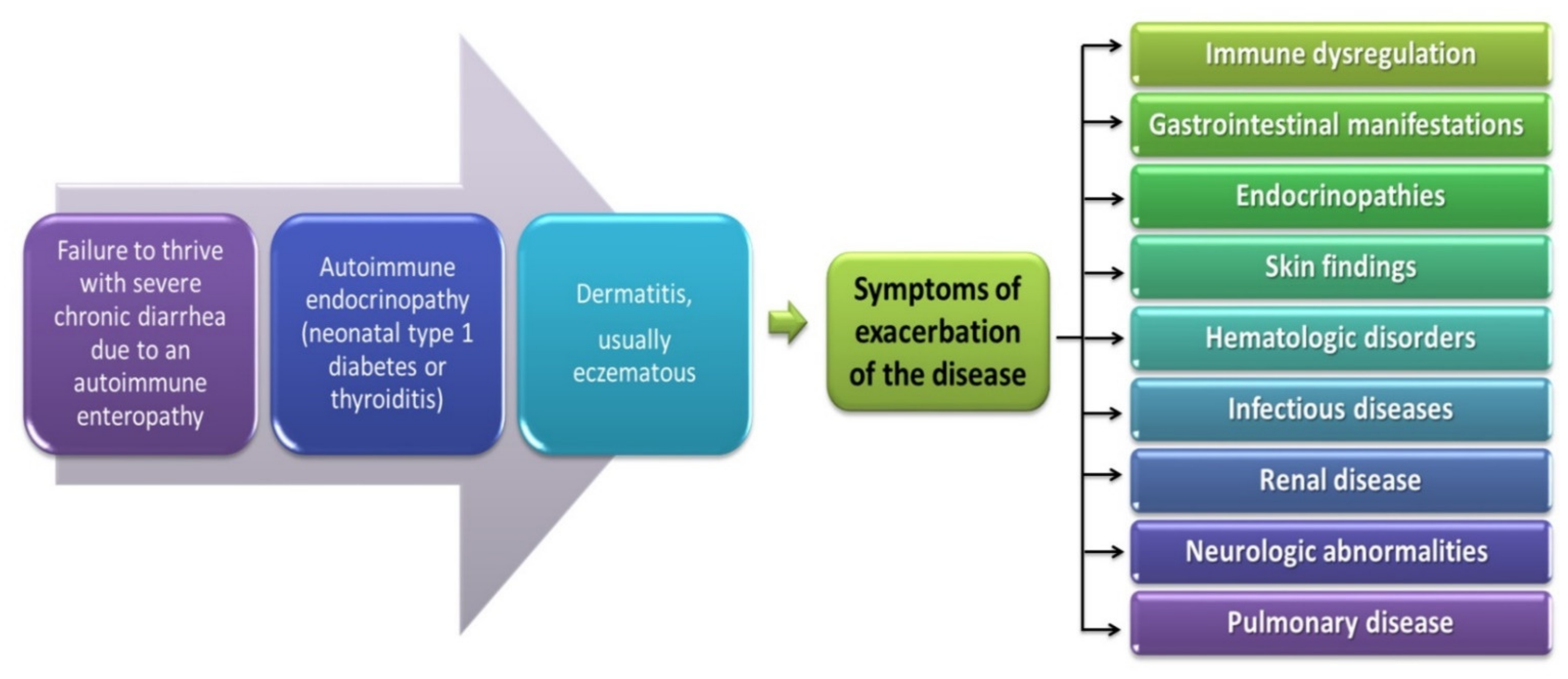
4.1.1. The Role of Foxp3 Protein in IPEX Development
4.1.2. The Role of the Foxp3 Protein in the Development of CVID
5. Materials and Methods
5.1. Search Strategy, Study Selection, and Data Extraction
5.2. Biostatistical Analysis
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mitsis, T.; Efthimiadou, A.; Bacopoulou, F.; Vlachakis, D.; Chrousos, G.; Eliopoulos, E. Transcription factors and evolution: An integral part of gene expression (Review). World Acad. Sci. J. 2020, 2, 3–8. [Google Scholar] [CrossRef]
- Lu, L.; Barbi, J.; Pan, F. The regulation of immune tolerance by FOXP3. Nat. Rev. Immunol. 2017, 17, 703–717. [Google Scholar] [CrossRef] [PubMed]
- Tai, X.; Erman, B.; Alag, A.; Mu, J.; Kimura, M.; Katz, G.; Guinter, T.; McCaughtry, T.; Etzensperger, R.; Feigenbaum, L.; et al. Foxp3 Transcription Factor Is Proapoptotic and Lethal to Developing Regulatory T Cells unless Counterbalanced by Cytokine Survival Signals. Immunity 2013, 38, 1116–1128. [Google Scholar] [CrossRef]
- Colamatteo, A.; Carbone, F.; Bruzzaniti, S.; Galgani, M.; Fusco, C.; Maniscalco, G.T.; Di Rella, F.; De Candia, P.; De Rosa, V. Molecular Mechanisms Controlling Foxp3 Expression in Health and Autoimmunity: From Epigenetic to Post-translational Regulation. Front. Immunol. 2020, 10, 3136. [Google Scholar] [CrossRef] [PubMed]
- Bluestone, J.A. FOXP3, the Transcription Factor at the Heart of the Rebirth of Immune Tolerance. J. Immunol. 2017, 198, 979–980. [Google Scholar] [CrossRef]
- Chang, X.; Zheng, P.; Liu, Y. FoxP3: A genetic link between immunodeficiency and autoimmune diseases. Autoimmun. Rev. 2006, 5, 399–402. [Google Scholar] [CrossRef]
- Justiz Vaillant, A.A.; Qurie, A. Immunodeficiency; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Primary Immunodeficiency-Symptoms and Causes. Available online: https://www.mayoclinic.org/diseases-conditions/primary-immunodeficiency/symptoms-causes/syc-20376905 (accessed on 30 December 2021).
- Sokol, K.; Milner, J.D. The overlap between allergy and immunodeficiency. Curr. Opin. Pediatr. 2018, 30, 848–854. [Google Scholar] [CrossRef]
- Amaya-Uribe, L.; Rojas, M.; Azizi, G.; Anaya, J.-M.; Gershwin, M.E. Primary immunodeficiency and autoimmunity: A comprehensive review. J. Autoimmun. 2019, 99, 52–72. [Google Scholar] [CrossRef] [PubMed]
- Todoric, K.; Koontz, J.B.; Mattox, D.; Tarrant, T.K. Autoimmunity in Immunodeficiency. Curr. Allergy Asthma Rep. 2013, 13, 361–370. [Google Scholar] [CrossRef]
- Cowen, J.E.; Stevenson, J.; Paravasthu, M.; Darroch, J.; Jacob, A.; Tueger, S.; Gosney, J.R.; Simons, A.; Spencer, L.G.; Judge, E.P. Common variable immunodeficiency with granulomatous-lymphocytic interstitial lung disease and preceding neurological involvement: A case-report. BMC Pulm. Med. 2020, 20, 205. [Google Scholar] [CrossRef]
- Mortaz, E.; Tabarsi, P.; Mansouri, D.; Khosravi, A.; Garssen, J.; Velayati, A.; Adcock, I.M. Cancers Related to Immunodeficiencies: Update and Perspectives. Front. Immunol. 2016, 7, 365. [Google Scholar] [CrossRef] [PubMed]
- Haas, O.A. Primary Immunodeficiency and Cancer Predisposition Revisited: Embedding Two Closely Related Concepts into an Integrative Conceptual Framework. Front. Immunol. 2019, 9, 3136. [Google Scholar] [CrossRef] [PubMed]
- Nozaki, T.; Takada, H.; Ishimura, M.; Ihara, K.; Imai, K.; Morio, T.; Kobayashi, M.; Nonoyama, S.; Hara, T. Endocrine complications in primary immunodeficiency diseases in Japan. Clin. Endocrinol. 2012, 77, 628–634. [Google Scholar] [CrossRef] [PubMed]
- Coopmans, E.C.; Chunharojrith, P.; Neggers, S.J.C.M.M.; Van Der Ent, M.W.; Swagemakers, S.M.A.; Hollink, I.H.; Barendregt, B.H.; Van Der Spek, P.J.; Van Der Lely, A.-J.; Van Hagen, P.M.; et al. Endocrine Disorders Are Prominent Clinical Features in Patients with Primary Antibody Deficiencies. Front. Immunol. 2019, 10, 2079. [Google Scholar] [CrossRef] [PubMed]
- Mohtashami, M.; Razavi, A.; Abolhassani, H.; Aghamohammadi, A.; Yazdani, R. Primary Immunodeficiency and Thrombocytopenia. Int. Rev. Immunol. 2021, 40, 1–43. [Google Scholar] [CrossRef] [PubMed]
- Rezaei, N.; Moazzami, K.; Aghamohammadi, A.; Klein, C. Neutropenia and Primary Immunodeficiency Diseases. Int. Rev. Immunol. 2009, 28, 335–366. [Google Scholar] [CrossRef]
- Sullivan, K.E. Neutropenia as a sign of immunodeficiency. J. Allergy Clin. Immunol. 2019, 143, 96–100. [Google Scholar] [CrossRef]
- Soler-Palacín, P.; De Gracia, J.; Gonzalez-Granado, L.I.; Martín, C.; Rodríguez-Gallego, C.; Sánchez-Ramón, S. Lung ID-Signal Group Primary immunodeficiency diseases in lung disease: Warning signs, diagnosis and management. Respir. Res. 2018, 19, 219. [Google Scholar] [CrossRef]
- Agarwal, S.; Mayer, L. Diagnosis and Treatment of Gastrointestinal Disorders in Patients with Primary Immunodeficiency. Clin. Gastroenterol. Hepatol. 2013, 11, 1050–1063. [Google Scholar] [CrossRef]
- Lazaridis, N. Current insights into the innate immune system dysfunction in irritable bowel syndrome. Ann. Gastroenterol. 2018, 31, 171–187. [Google Scholar] [CrossRef]
- Brede, K.K.; Wandel, M.; Wiig, I.; von der Lippe, C. Primary Immunodeficiency Diseases and Gastrointestinal Distress: Coping Strategies and Dietary Experiences to Relieve Symptoms. Qual. Health Res. 2020, 31, 361–372. [Google Scholar] [CrossRef] [PubMed]
- Overview of Immunodeficiency Disorders-Immunology; Allergic Disorders. Available online: https://www.msdmanuals.com/en-kr/professional/immunology-allergic-disorders/immunodeficiency-disorders/overview-of-immunodeficiency-disorders (accessed on 30 December 2021).
- Sánchez-Ramón, S.; Bermúdez, A.; González-Granado, L.I.; Rodríguez-Gallego, C.; Sastre, A.; Soler-Palacín, P.; the ID-Signal Onco-Haematology Group; Allende, L.; Alsina, L.; Bielsa, A.M.; et al. Primary and Secondary Immunodeficiency Diseases in Oncohaematology: Warning Signs, Diagnosis, and Management. Front. Immunol. 2019, 10, 586. [Google Scholar] [CrossRef] [PubMed]
- Brunkow, M.E.; Jeffery, E.W.; Hjerrild, K.A.; Paeper, B.; Clark, L.B.; Yasayko, S.-A.; Wilkinson, J.E.; Galas, D.; Ziegler, S.F.; Ramsdell, F. Disruption of a new forkhead/winged-helix protein, scurfin, results in the fatal lymphoproliferative disorder of the scurfy mouse. Nat. Genet. 2001, 27, 68–73. [Google Scholar] [CrossRef] [PubMed]
- Bennett, C.; Yoshioka, R.; Kiyosawa, H.; Barker, D.F.; Fain, P.R.; Shigeoka, A.O.; Chance, P.F. X-Linked Syndrome of Polyendocrinopathy, Immune Dysfunction, and Diarrhea Maps to Xp11.23-Xq13.3. Am. J. Hum. Genet. 2000, 66, 461–468. [Google Scholar] [CrossRef] [PubMed]
- Georgiev, P.; Charbonnier, L.-M.; Chatila, T.A. Regulatory T Cells: The Many Faces of Foxp3. J. Clin. Immunol. 2019, 39, 623–640. [Google Scholar] [CrossRef]
- FOXP3-Forkhead Box Protein P3-Homo Sapiens (Human)-FOXP3 Gene & Protein. Available online: https://www.uniprot.org/uniprot/Q9BZS1 (accessed on 30 December 2021).
- Kwon, H.; Chen, H.-M.; Mathis, D.; Benoist, C. Different molecular complexes that mediate transcriptional induction and repression by FoxP3. Nat. Immunol. 2017, 18, 1238–1248. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Martin, S.; Pfleger, C.; Du, J.; Buckner, J.H.; Bluestone, J.A.; Riley, J.L.; Ziegler, S.F. Cutting Edge: A Novel, Human-Specific Interacting Protein Couples FOXP3 to a Chromatin-Remodeling Complex That Contains KAP1/TRIM28. J. Immunol. 2013, 190, 4470–4473. [Google Scholar] [CrossRef]
- Magg, T.; Mannert, J.; Ellwart, J.W.; Schmid, I.; Albert, M.H. Subcellular localization of FOXP3 in human regulatory and nonregulatory T cells. Eur. J. Immunol. 2012, 42, 1627–1638. [Google Scholar] [CrossRef]
- Du, J.; Huang, C.; Zhou, B.; Ziegler, S.F. Isoform-Specific Inhibition of RORα-Mediated Transcriptional Activation by Human FOXP3. J. Immunol. 2008, 180, 4785–4792. [Google Scholar] [CrossRef]
- Mailer, R.K.W. Alternative Splicing of FOXP3—Virtue and Vice. Front. Immunol. 2018, 9, 530. [Google Scholar] [CrossRef]
- Mailer, R.K. IPEX as a Consequence of Alternatively Spliced FOXP3. Front. Pediatr. 2020, 8, 594375. [Google Scholar] [CrossRef] [PubMed]
- Kawajiri, K.; Ikuta, T.; Suzuki, T.; Kusaka, M.; Muramatsu, M.; Fujieda, K.; Tachibana, M.; Morohashi, K.-I. Role of the LXXLL-Motif and Activation Function 2 Domain in Subcellular Localization of Dax-1 (Dosage-Sensitive Sex Reversal-Adrenal Hypoplasia Congenita Critical Region on the X Chromosome, Gene 1). Mol. Endocrinol. 2003, 17, 994–1004. [Google Scholar] [CrossRef] [PubMed]
- Nakae, J.; Cao, Y.; Daitoku, H.; Fukamizu, A.; Ogawa, W.; Yano, Y.; Hayashi, Y. The LXXLL motif of murine forkhead transcription factor FoxO1 mediates Sirt1-dependent transcriptional activity. J. Clin. Investig. 2006, 116, 2473–2483. [Google Scholar] [CrossRef] [PubMed]
- Clerc, I.; Polakowski, N.; André-Arpin, C.; Cook, P.; Barbeau, B.; Mesnard, J.-M.; Lemasson, I. An Interaction between the Human T Cell Leukemia Virus Type 1 Basic Leucine Zipper Factor (HBZ) and the KIX Domain of p300/CBP Contributes to the Down-regulation of Tax-dependent Viral Transcription by HBZ. J. Biol. Chem. 2008, 283, 23903–23913. [Google Scholar] [CrossRef] [PubMed]
- Garber, P.M.; Vidanes, G.M.; Toczyski, D.P. Damage in transition. Trends Biochem. Sci. 2005, 30, 63–66. [Google Scholar] [CrossRef]
- Ikuta, T.; Watanabe, J.; Kawajiri, K. Characterization of the LxxLL Motif in the Aryl Hydrocarbon Receptor: Effects on Subcellular Localization and Transcriptional Activity. J. Biochem. 2002, 131, 79–85. [Google Scholar] [CrossRef]
- Cassandri, M.; Smirnov, A.; Novelli, F.; Pitolli, C.; Agostini, M.; Malewicz, M.; Melino, G.; Raschellà, G. Zinc-finger proteins in health and disease. Cell Death Discov. 2017, 3, 17071. [Google Scholar] [CrossRef]
- Song, X.; Li, B.; Xiao, Y.; Chen, C.; Wang, Q.; Liu, Y.; Berezov, A.; Xu, C.; Gao, Y.; Li, Z.; et al. Structural and Biological Features of FOXP3 Dimerization Relevant to Regulatory T Cell Function. Cell Rep. 2012, 1, 665–675. [Google Scholar] [CrossRef]
- Li, J.; Jiang, L.; Liang, X.; Qu, L.; Wu, D.; Chen, X.; Guo, M.; Chen, Z.; Chen, L.; Chen, Y. DNA-binding properties of FOXP3 transcription factor. Acta Biochim. Biophys. Sin. 2017, 49, 792–799. [Google Scholar] [CrossRef]
- Chae, W.-J.; Henegariu, O.; Lee, S.-K.; Bothwell, A.L.M. The mutant leucine-zipper domain impairs both dimerization and suppressive function of Foxp3 in T cells. Proc. Natl. Acad. Sci. USA 2006, 103, 9631–9636. [Google Scholar] [CrossRef]
- Lopes, J.E.; Torgerson, T.R.; Schubert, L.A.; Anover, S.D.; Ocheltree, E.L.; Ochs, H.D.; Ziegler, S.F. Analysis of FOXP3 Reveals Multiple Domains Required for Its Function as a Transcriptional Repressor. J. Immunol. 2006, 177, 3133–3142. [Google Scholar] [CrossRef] [PubMed]
- Mackey-Cushman, S.L.; Gao, J.; Holmes, A.D.; Nunoya, J.-I.; Wang, R.; Unutmaz, D.; Su, L. FoxP3 interacts with linker histone H1.5 to modulate gene expression and program Treg cell activity. Genes Immun. 2011, 12, 559–567. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lozano, T.; Casares, N.; Lasarte, J.J. Searching for the Achilles Heel of FOXP3. Front. Oncol. 2013, 3, 294. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, L.; Han, R.; Beier, U.H.; Hancock, W.W. Two Lysines in the Forkhead Domain of Foxp3 Are Key to T Regulatory Cell Function. PLoS ONE 2012, 7, e29035. [Google Scholar] [CrossRef]
- Wu, Y.; Borde, M.; Heissmeyer, V.; Feuerer, M.; Lapan, A.D.; Stroud, J.; Bates, D.L.; Guo, L.; Han, A.; Ziegler, S.F.; et al. FOXP3 Controls Regulatory T Cell Function through Cooperation with NFAT. Cell 2006, 126, 375–387. [Google Scholar] [CrossRef]
- Torgerson, T.R.; Genin, A.; Chen, C.; Zhang, M.; Zhou, B.; Añover-Sombke, S.; Frank, M.B.; Dozmorov, I.; Ocheltree, E.; Kulmala, P.; et al. FOXP3 Inhibits Activation-Induced NFAT2 Expression in T Cells Thereby Limiting Effector Cytokine Expression. J. Immunol. 2009, 183, 907–915. [Google Scholar] [CrossRef]
- Romano, M.; Fanelli, G.; Albany, C.J.; Giganti, G.; Lombardi, G. Past, Present, and Future of Regulatory T Cell Therapy in Transplantation and Autoimmunity. Front. Immunol. 2019, 10, 43. [Google Scholar] [CrossRef]
- Yu, G.; Berquist, S.; Huang, J.; Martinez, J.; Hoyte, E.; Vanichsarn, C.; Nadeau, K. Natural Regulatory T Cell Dysfunction in Subjects with Common Variable Immunodeficiency Complicated by Autoimmune Disease is associated with activation of the AkT-mTOR pathway. J. Allergy Clin. Immunol. 2009, 123, S264. [Google Scholar] [CrossRef]
- Rudensky, A.Y. Regulatory T cells and Foxp3. Immunol. Rev. 2011, 241, 260–268. [Google Scholar] [CrossRef]
- Bacchetta, R.; Gambineri, E.; Roncarolo, M.-G. Role of regulatory T cells and FOXP3 in human diseases. J. Allergy Clin. Immunol. 2007, 120, 227–235. [Google Scholar] [CrossRef]
- Li, Z.; Li, D.; Tsun, A.; Li, B. FOXP3+ regulatory T cells and their functional regulation. Cell. Mol. Immunol. 2015, 12, 558–565. [Google Scholar] [CrossRef] [PubMed]
- Barnes, M.J.; Griseri, T.; Johnson, A.M.F.; Young, W.; Powrie, F.; Izcue, A. CTLA-4 promotes Foxp3 induction and regulatory T cell accumulation in the intestinal lamina propria. Mucosal Immunol. 2012, 6, 324–334. [Google Scholar] [CrossRef]
- Xiao, X.; Shi, X.; Fan, Y.; Zhang, X.; Wu, M.; Lan, P.; Minze, L.; Fu, Y.-X.; Ghobrial, R.M.; Liu, W.; et al. GITR subverts Foxp3+ Tregs to boost Th9 immunity through regulation of histone acetylation. Nat. Commun. 2015, 6, 8266. [Google Scholar] [CrossRef]
- Bertolini, T.B.; Piñeros, A.R.; Prado, R.Q.; Gembre, A.F.; Ramalho, L.N.Z.; Alves-Filho, J.C.; Bonato, V.L.D. CCR4-dependent reduction in the number and suppressor function of CD4+Foxp3+ cells augments IFN-γ-mediated pulmonary inflammation and aggravates tuberculosis pathogenesis. Cell Death Dis. 2018, 10, 11. [Google Scholar] [CrossRef] [PubMed]
- Klapa, S.; Mueller, A.; Csernok, E.; Fagin, U.; Klenerman, P.; Holl-Ulrich, K.; Gross, W.L.; Lamprecht, P. Lower numbers of FoxP3 and CCR4 co-expressing cells in an elevated subpopulation of CD4+CD25high regulatory T cells from Wegener’s granulomatosis. Clin. Exp. Rheumatol. 2010, 28, 72–80. [Google Scholar]
- Pelletier, B.; Mondoulet, L.; Ligouis, M.; Dhelft, V.; Plaquet, C.; Dupont, C.M.; Benhamou, P.-H.; Sampson, H.A. Foxp3+CD62L+ Tregs induced by EPIT have the potency to suppress effector T cells proliferation in specific and bystander conditions. J. Allergy Clin. Immunol. 2017, 139, AB255. [Google Scholar] [CrossRef]
- Xu, L.; Kitani, A.; Strober, W. Molecular mechanisms regulating TGF-β-induced Foxp3 expression. Mucosal Immunol. 2010, 3, 230–238. [Google Scholar] [CrossRef]
- Lin, X.; Chen, M.; Liu, Y.; Guo, Z.; He, X.; Brand, D.; Zheng, S.G. Advances in distinguishing natural from induced Foxp3(+) regulatory T cells. Int. J. Clin. Exp. Pathol. 2013, 6, 116–123. [Google Scholar]
- Cosovanu, C.; Neumann, C. The Many Functions of Foxp3+ Regulatory T Cells in the Intestine. Front. Immunol. 2020, 11, 600973. [Google Scholar] [CrossRef]
- Lio, C.-W.J.; Dodson, L.F.; Deppong, C.M.; Hsieh, C.-S.; Green, J.M. CD28 Facilitates the Generation of Foxp3− Cytokine Responsive Regulatory T Cell Precursors. J. Immunol. 2010, 184, 6007–6013. [Google Scholar] [CrossRef]
- He, X.; Smeets, R.L.; Van Rijssen, E.; Boots, A.M.H.; Joosten, I.; Koenen, H.J.P.M. Single CD28 stimulation induces stable and polyclonal expansion of human regulatory T cells. Sci. Rep. 2017, 7, 43003. [Google Scholar] [CrossRef]
- Freudenberg, K.; Lindner, N.; Dohnke, S.; Garbe, A.I.; Schallenberg, S.; Kretschmer, K. Critical Role of TGF-β and IL-2 Receptor Signaling in Foxp3 Induction by an Inhibitor of DNA Methylation. Front. Immunol. 2018, 9, 125. [Google Scholar] [CrossRef]
- Schuster, M.; Plaza-Sirvent, C.; Visekruna, A.; Huehn, J.; Schmitz, I. Generation of Foxp3+CD25− Regulatory T-Cell Precursors Requires c-Rel and IκBNS. Front. Immunol. 2019, 10, 1583. [Google Scholar] [CrossRef]
- Won, H.Y.; Shin, J.H.; Oh, S.; Jeong, H.; Hwang, E.S. Enhanced CD25+Foxp3+ regulatory T cell development by amodiaquine through activation of nuclear receptor 4A. Sci. Rep. 2017, 7, 16946. [Google Scholar] [CrossRef]
- Shevach, E.M. Foxp3+ T Regulatory Cells: Still Many Unanswered Questions—A Perspective after 20 Years of Study. Front. Immunol. 2018, 9, 1048. [Google Scholar] [CrossRef]
- Chien, C.-H.; Chiang, B.-L. Regulatory T cells induced by B cells: A novel subpopulation of regulatory T cells. J. Biomed. Sci. 2017, 24, 86. [Google Scholar] [CrossRef]
- Engelmann, P.; Farkas, K.; Kis, J.; Richman, G.; Zhang, Z.; Liew, C.W.; Borowiec, M.; Niewczas, M.A.; Jalahej, H.; Orbán, T. Characterization of human invariant natural killer T cells expressing FoxP3. Int. Immunol. 2011, 23, 473–484. [Google Scholar] [CrossRef] [PubMed]
- Niemeyer, M.; Darmoise, A.; Mollenkopf, H.-J.; Hahnke, K.; Hurwitz, R.; Besra, G.S.; Schaible, U.E.; Kaufmann, S.H.E. Natural killer T-cell characterization through gene expression profiling: An account of versatility bridging T helper type 1 (Th1), Th2 and Th17 immune responses. Immunology 2007, 123, 45–56. [Google Scholar] [CrossRef]
- Koguchi, Y.; Buenafe, A.C.; Thauland, T.J.; Gardell, J.L.; Bivins-Smith, E.R.; Jacoby, D.B.; Slifka, M.K.; Parker, D.C. Preformed CD40L Is Stored in Th1, Th2, Th17, and T Follicular Helper Cells as Well as CD4+8− Thymocytes and Invariant NKT Cells but Not in Treg Cells. PLoS ONE 2012, 7, e31296. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, M.; Almeida, C.F.; Caridade, M.; Ribot, J.; Duarte, J.; Agua-Doce, A.; Wollenberg, I.; Silva-Santos, B.; Graca, L. Identification of Regulatory Foxp3+ Invariant NKT Cells Induced by TGF-β. J. Immunol. 2010, 185, 2157–2163. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Guo, Y.; Xu, L.; Li, Y.; Cao, L. Foxp3 regulates ratio of Treg and NKT cells in a mouse model of asthma. Mol. Cell. Biochem. 2015, 403, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Noh, J.; Choi, W.S.; Noh, G.; Lee, J.H. Presence ofFoxp3-expressing CD19(+)CD5(+) B Cells in Human Peripheral Blood Mononuclear Cells: Human CD19(+)CD5(+)Foxp3(+) Regulatory B Cell (Breg). Immune Netw. 2010, 10, 247–249. [Google Scholar] [CrossRef]
- Mohajeri, M.; Farazmand, A.; Bonab, M.M.; Nikbin, B.; Minagar, A. FOXP3 gene expression in multiple sclerosis patients pre- and post mesenchymal stem cell therapy. Iran. J. Allergy Asthma Immunol. 2011, 10, 155–161. [Google Scholar]
- Abbass, A.A.; Mohamed, A.N.; Abdel-Rehim, A.S.M. Association of FOXP3 regulatory gene expression with systemic lupus erythematosus disease activity among Egyptian patients. Egypt. J. Immunol. 2013, 20, 21–28. [Google Scholar] [PubMed]
- Bonelli, M.; Göschl, L.; Blüml, S.; Rath, E.; Smolen, J.S.; Scheinecker, C. CD4+CD25-Foxp3+ T cells are increased in systemic lupus erythematosus patients with active glomerulonephritis. Ann. Rheum. Dis. 2011, 70, A47–A48. [Google Scholar] [CrossRef]
- Slobodin, G.; Sheikh-Ahmad, M.; Rosner, I.; Peri, R.; Rozenbaum, M.; Kessel, A.; Toubi, E.; Odeh, M. Regulatory T cells (CD4+CD25brightFoxP3+) expansion in systemic sclerosis correlates with disease activity and severity. Cell. Immunol. 2010, 261, 77–80. [Google Scholar] [CrossRef]
- Vadasz, Z.; Toubi, E. FoxP3 Expression in Macrophages, Cancer, and B Cells—Is It Real? Clin. Rev. Allergy Immunol. 2016, 52, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Lozano, T.; Villanueva, L.; Durantez, M.; Gorraiz, M.; Ruiz, M.; Belsue, V.; Riezu-Boj, J.-I.; Hervás-Stubbs, S.; Oyarzabal, J.; Bandukwala, H.; et al. Inhibition of FOXP3/NFAT Interaction Enhances T Cell Function after TCR Stimulation. J. Immunol. 2015, 195, 3180–3189. [Google Scholar] [CrossRef]
- Ono, M.; Yaguchi, H.; Ohkura, N.; Kitabayashi, I.; Nagamura, Y.; Nomura, T.; Miyachi, Y.; Tsukada, T.; Sakaguchi, S. Foxp3 controls regulatory T-cell function by interacting with AML1/Runx1. Nature 2007, 446, 685–689. [Google Scholar] [CrossRef] [PubMed]
- Maeda, M.; Matsuzaki, H.; Yamamoto, S.; Lee, S.; Kumagai-Takei, N.; Yoshitome, K.; Min, Y.; Sada, N.; Nishimura, Y.; Otsuki, T. Aberrant expression of FoxP3 in a human T cell line possessing regulatory T cell-like function and exposed continuously to asbestos fibers. Oncol. Rep. 2018, 40, 748–758. [Google Scholar] [CrossRef] [PubMed]
- Kitoh, A.; Ono, M.; Naoe, Y.; Ohkura, N.; Yamaguchi, T.; Yaguchi, H.; Kitabayashi, I.; Tsukada, T.; Nomura, T.; Miyachi, Y.; et al. Indispensable Role of the Runx1-Cbfβ Transcription Complex for In Vivo-Suppressive Function of FoxP3+ Regulatory T Cells. Immunity 2009, 31, 609–620. [Google Scholar] [CrossRef]
- Walker, L.S. Treg and CTLA-4: Two intertwining pathways to immune tolerance. J. Autoimmun. 2013, 45, 49–57. [Google Scholar] [CrossRef]
- Fan, M.Y.; Low, J.S.; Tanimine, N.; Finn, K.K.; Priyadharshini, B.; Germana, S.K.; Kaech, S.M.; Turka, L.A. Differential Roles of IL-2 Signaling in Developing versus Mature Tregs. Cell Rep. 2018, 25, 1204–1213.e4. [Google Scholar] [CrossRef]
- Ferreira, R.C.; Simons, H.Z.; Thompson, W.S.; Rainbow, D.B.; Yang, X.; Cutler, A.J.; Oliveira, J.; Dopico, X.C.; Smyth, D.; Savinykh, N.; et al. Cells with Treg-specific FOXP3 demethylation but low CD25 are prevalent in autoimmunity. J. Autoimmun. 2017, 84, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Driesen, J.; Popov, A.; Schultze, J.L. CD25 as an immune regulatory molecule expressed on myeloid dendritic cells. Immunobiology 2008, 213, 849–858. [Google Scholar] [CrossRef] [PubMed]
- Brunner-Weinzierl, M.; Rudd, C.E. CTLA-4 and PD-1 Control of T-Cell Motility and Migration: Implications for Tumor Immunotherapy. Front. Immunol. 2018, 9, 2737. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-H.; Hwang, J.; Jung, J.H.; Lee, H.-J.; Lee, D.Y.; Kim, S.-H. Molecular networks of FOXP family: Dual biologic functions, interplay with other molecules and clinical implications in cancer progression. Mol. Cancer 2019, 18, 180. [Google Scholar] [CrossRef]
- Zorn, E.; Nelson, E.A.; Mohseni, M.; Porcheray, F.; Kim, H.; Litsa, D.; Bellucci, R.; Raderschall, E.; Canning, C.; Soiffer, R.J.; et al. IL-2 regulates FOXP3 expression in human CD4+CD25+ regulatory T cells through a STAT-dependent mechanism and induces the expansion of these cells in vivo. Blood 2006, 108, 1571–1579. [Google Scholar] [CrossRef]
- Hench, V.K.; Su, L. Regulation of IL-2 gene expression by Siva and FOXP3 in human T cells. BMC Immunol. 2011, 12, 54. [Google Scholar] [CrossRef][Green Version]
- Bendfeldt, H.; Benary, M.; Scheel, T.; Steinbrink, K.; Radbruch, A.; Herzel, H.; Baumgrass, R. IL-2 Expression in Activated Human Memory FOXP3+ Cells Critically Depends on the Cellular Levels of FOXP3 as Well as of Four Transcription Factors of T Cell Activation. Front. Immunol. 2012, 3, 264. [Google Scholar] [CrossRef]
- Dikiy, S.; Li, J.; Bai, L.; Jiang, M.; Janke, L.; Zong, X.; Hao, X.; Hoyos, B.; Wang, Z.-M.; Xu, B.; et al. A distal Foxp3 enhancer enables interleukin-2 dependent thymic Treg cell lineage commitment for robust immune tolerance. Immunity 2021, 54, 931–946.e11. [Google Scholar] [CrossRef] [PubMed]
- Konopacki, C.; Pritykin, Y.; Rubtsov, Y.; Leslie, C.S.; Rudensky, A.Y. Transcription factor Foxp1 regulates Foxp3 chromatin binding and coordinates regulatory T cell function. Nat. Immunol. 2019, 20, 232–242. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Rowell, E.A.; Thomas, R.M.; Hancock, W.W.; Wells, A.D. Transcriptional Regulation by Foxp3 Is Associated with Direct Promoter Occupancy and Modulation of Histone Acetylation. J. Biol. Chem. 2006, 281, 36828–36834. [Google Scholar] [CrossRef]
- Ohkura, N.; Sakaguchi, S. Transcriptional and epigenetic basis of Treg cell development and function: Its genetic anomalies or variations in autoimmune diseases. Cell Res. 2020, 30, 465–474. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Ni, B.; Tian, Y.; Tian, Z.; Chen, Y.; Liu, Z.; Yang, X.; Lv, Y.; Zhang, Y. Histone methylation mediates plasticity of human FOXP3+regulatory T cells by modulating signature gene expressions. Immunology 2013, 141, 362–376. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Samanta, A.; Song, X.; Iacono, K.T.; Bembas, K.; Tao, R.; Basu, S.; Riley, J.; Hancock, W.W.; Shen, Y.; et al. FOXP3 interactions with histone acetyltransferase and class II histone deacetylases are required for repression. Proc. Natl. Acad. Sci. USA 2007, 104, 4571–4576. [Google Scholar] [CrossRef]
- Dahiya, S.; Beier, U.H.; Wang, L.; Han, R.; Jiao, J.; Akimova, T.; Angelin, A.; Wallace, D.C.; Hancock, W.W. HDAC10 deletion promotes Foxp3+ T-regulatory cell function. Sci. Rep. 2020, 10, 424. [Google Scholar] [CrossRef] [PubMed]
- Beier, U.; Akimova, T.; Liu, Y.; Wang, L.; Hancock, W.W. Histone/protein deacetylases control Foxp3 expression and the heat shock response of T-regulatory cells. Curr. Opin. Immunol. 2011, 23, 670–678. [Google Scholar] [CrossRef]
- Zhang, H.; Xiao, Y.; Zhu, Z.; Li, B.; Greene, I.M. Immune regulation by histone deacetylases: A focus on the alteration of FOXP3 activity. Immunol. Cell Biol. 2011, 90, 95–100. [Google Scholar] [CrossRef]
- van Loosdregt, J.; Vercoulen, Y.; Guichelaar, T.; Gent, Y.Y.J.; Beekman, J.M.; Van Beekum, O.; Brenkman, A.B.; Hijnen, D.; Mutis, T.; Kalkhoven, E.; et al. Regulation of Treg functionality by acetylation-mediated Foxp3 protein stabilization. Blood 2010, 115, 965–974. [Google Scholar] [CrossRef]
- Deng, G.; Song, X.; Fujimoto, S.; Piccirillo, C.A.; Nagai, Y.; Greene, M.I. Foxp3 Post-translational Modifications and Treg Suppressive Activity. Front. Immunol. 2019, 10, 2486. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Nagai, Y.; Deng, G.; Ohtani, T.; Zhu, Z.; Zhou, Z.; Zhang, H.; Ji, M.Q.; Lough, J.W.; Samanta, A.; et al. Dynamic Interactions between TIP60 and p300 Regulate FOXP3 Function through a Structural Switch Defined by a Single Lysine on TIP60. Cell Rep. 2014, 7, 1471–1480. [Google Scholar] [CrossRef] [PubMed]
- De Zoeten, E.; Wang, L.; Butler, K.V.; Beier, U.; Akimova, T.; Sai, H.; Bradner, J.E.; Mazitschek, R.; Kozikowski, A.P.; Matthias, P.; et al. Histone Deacetylase 6 and Heat Shock Protein 90 Control the Functions of Foxp3+ T-Regulatory Cells. Mol. Cell. Biol. 2011, 31, 2066–2078. [Google Scholar] [CrossRef] [PubMed]
- Mollapour, M.; Neckers, L. Post-translational modifications of Hsp90 and their contributions to chaperone regulation. Biochim. Biophys. Acta 2012, 1823, 648–655. [Google Scholar] [CrossRef]
- Nagai, Y.; Lam, L.; Greene, M.I.; Zhang, H. FOXP3 and Its Cofactors as Targets of Immunotherapies. Engineering 2019, 5, 115–121. [Google Scholar] [CrossRef]
- Van Loosdregt, J.; Brunen, D.; Fleskens, V.; Pals, C.E.G.M.; Lam, E.W.F.; Coffer, P.J. Rapid Temporal Control of Foxp3 Protein Degradation by Sirtuin-1. PLoS ONE 2011, 6, e19047. [Google Scholar] [CrossRef] [PubMed]
- Polansky, J.K.; Kretschmer, K.; Freyer, J.; Floess, S.; Garbe, A.; Baron, U.; Olek, S.; Hamann, A.; von Boehmer, H.; Huehn, J. DNA methylation controls Foxp3 gene expression. Eur. J. Immunol. 2008, 38, 1654–1663. [Google Scholar] [CrossRef] [PubMed]
- Morikawa, H.; Ohkura, N.; Vandenbon, A.; Itoh, M.; Nagao-Sato, S.; Kawaji, H.; Lassmann, T.; Carninci, P.; Hayashizaki, Y.; Forrest, A.; et al. Differential roles of epigenetic changes and Foxp3 expression in regulatory T cell-specific transcriptional regulation. Proc. Natl. Acad. Sci. USA 2014, 111, 5289–5294. [Google Scholar] [CrossRef]
- Maruyama, T.; Konkel, J.E.; Zamarron, B.F.; Chen, W. The molecular mechanisms of Foxp3 gene regulation. Semin. Immunol. 2011, 23, 418–423. [Google Scholar] [CrossRef]
- Zheng, Y.; Josefowicz, S.; Chaudhry, A.; Peng, X.P.; Forbush, K.; Rudensky, A.Y. Role of conserved non-coding DNA elements in the Foxp3 gene in regulatory T-cell fate. Nature 2010, 463, 808–812. [Google Scholar] [CrossRef]
- Montauti, E.; Zhang, Y.; Cortez, J.; Marson, A.; Fang, D. USP22 Stabilizes FoxP3 to Maintain Regulatory T Cell Function through a Dual Mechanism Resulting in Tumor Immune Evasion. J. Immunol. 2020, 204, 244.1. [Google Scholar]
- Montauti, E.; Zhang, Y.; Fang, D. USP22 Maintains Regulatory T Cells by Stabilizing Foxp3 Protein Level and Impairs Anti-Tumor Immune Response. J. Immunol. 2018, 200, 57.10. [Google Scholar]
- Zhang, Y.; Liu, W.; Chen, Y.; Liu, J.; Wu, K.; Su, L.; Zhang, W.; Jiang, Y.; Zhang, X.; Zhang, Y.; et al. A Cellular MicroRNA Facilitates Regulatory T Lymphocyte Development by Targeting the FOXP3 Promoter TATA-Box Motif. J. Immunol. 2017, 200, 1053–1063. [Google Scholar] [CrossRef]
- Hippen, K.L.; Loschi, M.; Nicholls, J.; MacDonald, K.P.A.; Blazar, B.R. Effects of MicroRNA on Regulatory T Cells and Implications for Adoptive Cellular Therapy to Ameliorate Graft-versus-Host Disease. Front. Immunol. 2018, 9, 57. [Google Scholar] [CrossRef]
- Dong, Y.; Yang, C.; Pan, F. Post-Translational Regulations of Foxp3 in Treg Cells and Their Therapeutic Applications. Front. Immunol. 2021, 12, 6172. [Google Scholar] [CrossRef] [PubMed]
- Bin Dhuban, K.; D’Hennezel, E.; Nagai, Y.; Xiao, Y.; Shao, S.; Istomine, R.; Alvarez, F.; Ben-Shoshan, M.; Ochs, H.; Mazer, B.; et al. Suppression by human FOXP3+ regulatory T cells requires FOXP3-TIP60 interactions. Sci. Immunol. 2017, 2, eaai9297. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, L.; Han, R.; Beier, U.; Akimova, T.; Bhatti, T.; Xiao, H.; Cole, P.A.; Brindle, P.; Hancock, W.W. Two Histone/Protein Acetyltransferases, CBP and p300, Are Indispensable for Foxp3+ T-Regulatory Cell Development and Function. Mol. Cell. Biol. 2014, 34, 3993–4007. [Google Scholar] [CrossRef]
- Wang, L.; Beier, U.; Akimova, T.; Dahiya, S.; Han, R.; Samanta, A.; Levine, M.H.; Hancock, W.W. Histone/protein deacetylase inhibitor therapy for enhancement of Foxp3+ T-regulatory cell function posttransplantation. Am. J. Transplant. 2018, 18, 1596–1603. [Google Scholar] [CrossRef]
- Wang, L.; Liu, Y.; Han, R.; Beier, U.H.; Bhatti, T.R.; Akimova, T.; Greene, M.I.; Hiebert, S.W.; Hancock, W.W. FOXP3+ regulatory T cell development and function require histone/protein deacetylase 3. J. Clin. Investig. 2015, 125, 1111–1123. [Google Scholar] [CrossRef]
- De Zoeten, E.; Wang, L.; Sai, H.; Dillmann, W.H.; Hancock, W.W. Inhibition of HDAC9 Increases T Regulatory Cell Function and Prevents Colitis in Mice. Gastroenterology 2010, 138, 583–594. [Google Scholar] [CrossRef]
- Yang, X.; Lun, Y.; Jiang, H.; Liu, X.; Duan, Z.; Xin, S.; Zhang, J. SIRT1-Regulated Abnormal Acetylation of FOXP3 Induces Regulatory T-Cell Function Defect in Hashimoto’s Thyroiditis. Thyroid 2018, 28, 246–256. [Google Scholar] [CrossRef]
- Beier, U.H.; Wang, L.; Bhatti, T.R.; Liu, Y.; Han, R.; Ge, G.; Hancock, W.W. Sirtuin-1 Targeting Promotes Foxp3+ T-Regulatory Cell Function and Prolongs Allograft Survival. Mol. Cell. Biol. 2011, 31, 1022–1029. [Google Scholar] [CrossRef]
- Kempkes, R.; Joosten, I.; Koenen, H.J.P.M.; He, X. Metabolic Pathways Involved in Regulatory T Cell Functionality. Front. Immunol. 2019, 10, 2839. [Google Scholar] [CrossRef]
- De Rosa, V.; Galgani, M.; Porcellini, A.; Colamatteo, A.; Santopaolo, M.; Zuchegna, C.; Romano, A.; De Simone, S.; Procaccini, C.; La Rocca, C.; et al. Glycolysis controls the induction of human regulatory T cells by modulating the expression of FOXP3 exon 2 splicing variants. Nat. Immunol. 2015, 16, 1174–1184. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Chi, H. Metabolic Control of Treg Cell Stability, Plasticity, and Tissue-Specific Heterogeneity. Front. Immunol. 2019, 10, 2716. [Google Scholar] [CrossRef] [PubMed]
- Chapman, N.M.; Chi, H. mTOR signaling, Tregs and immune modulation. Immunotherapy 2014, 6, 1295–1311. [Google Scholar] [CrossRef] [PubMed]
- An, J.; Ding, Y.; Yu, C.; Li, J.; You, S.; Liu, Z.; Song, P.; Zou, M.-H. AMP-activated protein kinase alpha1 promotes tumor development via FOXP3 elevation in tumor-infiltrating Treg cells. iScience 2022, 25, 103570. [Google Scholar] [CrossRef]
- He, N.; Fan, W.; Henriquez, B.; Yu, R.T.; Atkins, A.R.; Liddle, C.; Zheng, Y.; Downes, M.; Evans, R.M. Metabolic control of regulatory T cell (Treg) survival and function by Lkb1. Proc. Natl. Acad. Sci. USA 2017, 114, 12542–12547. [Google Scholar] [CrossRef]
- Raud, B.; Roy, D.G.; Divakaruni, A.S.; Tarasenko, T.N.; Franke, R.; Ma, E.H.; Samborska, B.; Hsieh, W.Y.; Wong, A.H.; Stüve, P.; et al. Etomoxir Actions on Regulatory and Memory T Cells Are Independent of Cpt1a-Mediated Fatty Acid Oxidation. Cell Metab. 2018, 28, 504–515.e7. [Google Scholar] [CrossRef]
- Zhu, X.; Ji, W.; Guo, S.; Zhu, D.; Yang, Y.; Liu, X. Glycolytic and lipid oxidative metabolic programs are essential for freshly-isolated regulatory T cells in mice with sepsis. RSC Adv. 2020, 10, 21000–21008. [Google Scholar] [CrossRef]
- Brown, Z.J.; Fu, Q.; Ma, C.; Kruhlak, M.; Zhang, H.; Luo, J.; Heinrich, B.; Yu, S.J.; Zhang, Q.; Wilson, A.; et al. Carnitine palmitoyltransferase gene upregulation by linoleic acid induces CD4+ T cell apoptosis promoting HCC development. Cell Death Dis. 2018, 9, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Atif, M.; Mohr, A.; Conti, F.; Scatton, O.; Gorochov, G.; Miyara, M. Metabolic Optimisation of Regulatory T Cells in Transplantation. Front. Immunol. 2020, 11, 2005. [Google Scholar] [CrossRef] [PubMed]
- Witt, D.R.; Kellogg, R.A.; Snyder, M.P.; Dunn, J. Windows into human health through wearables data analytics. Curr. Opin. Biomed. Eng. 2019, 9, 28–46. [Google Scholar] [CrossRef]
- Yuan, Z.; Peng, L.; Radhakrishnan, R.; Seto, E. Histone Deacetylase 9 (HDAC9) Regulates the Functions of the ATDC (TRIM29) Protein. J. Biol. Chem. 2010, 285, 39329–39338. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Pili, R. Class I histone deacetylase inhibition is a novel mechanism to target regulatory T cells in immunotherapy. OncoImmunology 2012, 1, 948–950. [Google Scholar] [CrossRef][Green Version]
- Terranova-Barberio, M.; Thomas, S.; Ali, N.; Pawlowska, N.; Park, J.; Krings, G.; Rosenblum, M.D.; Budillon, A.; Munster, P.N. HDAC inhibition potentiates immunotherapy in triple negative breast cancer. Oncotarget 2017, 8, 114156–114172. [Google Scholar] [CrossRef]
- Chen, I.-C.; Sethy, B.; Liou, J.-P. Recent Update of HDAC Inhibitors in Lymphoma. Front. Cell Dev. Biol. 2020, 8, 906. [Google Scholar] [CrossRef]
- Akimova, T.; Ge, G.; Golovina, T.; Mikheeva, T.; Wang, L.; Riley, J.L.; Hancock, W.W. Histone/protein deacetylase inhibitors increase suppressive functions of human FOXP3+ Tregs. Clin. Immunol. 2010, 136, 348–363. [Google Scholar] [CrossRef]
- Xiao, Y.; Li, B.; Zhou, Z.; Hancock, W.W.; Zhang, H.; I Greene, M. Histone acetyltransferase mediated regulation of FOXP3 acetylation and Treg function. Curr. Opin. Immunol. 2010, 22, 583–591. [Google Scholar] [CrossRef]
- Bettini, M.L.; Pan, F.; Bettini, M.; Finkelstein, D.; Rehg, J.E.; Floess, S.; Bell, B.D.; Ziegler, S.F.; Huehn, J.; Pardoll, D.M.; et al. Loss of Epigenetic Modification Driven by the Foxp3 Transcription Factor Leads to Regulatory T Cell Insufficiency. Immunity 2012, 36, 717–730. [Google Scholar] [CrossRef]
- Wang, L.; de Zoeten, E.; Greene, M.I.; Hancock, W.W. Immunomodulatory effects of deacetylase inhibitors: Therapeutic targeting of FOXP3+ regulatory T cells. Nat. Rev. Drug Discov. 2009, 8, 969–981. [Google Scholar] [CrossRef] [PubMed]
- Saouaf, S.J.; Li, B.; Zhang, G.; Shen, Y.; Furuuchi, N.; Hancock, W.W.; Greene, M.I. Deacetylase inhibition increases regulatory T cell function and decreases incidence and severity of collagen-induced arthritis. Exp. Mol. Pathol. 2009, 87, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; He, X.; Zhang, L.; Zhang, R.; Li, W. Acetylation in Tumor Immune Evasion Regulation. Front. Pharmacol. 2021, 12, 1558. [Google Scholar] [CrossRef] [PubMed]
- Grover, P.; Goel, P.N.; Piccirillo, C.A.; Greene, M.I. FOXP3 and Tip60 Structural Interactions Relevant to IPEX Development Lead to Potential Therapeutics to Increase FOXP3 Dependent Suppressor T Cell Functions. Front. Pediatr. 2021, 9, 7292. [Google Scholar] [CrossRef] [PubMed]
- Santer, F.R.; Höschele, P.P.; Oh, S.J.; Erb, H.H.; Bouchal, J.; Cavarretta, I.T.; Parson, W.; Meyers, D.J.; Cole, P.A.; Culig, Z. Inhibition of the Acetyltransferases p300 and CBP Reveals a Targetable Function for p300 in the Survival and Invasion Pathways of Prostate Cancer Cell Lines. Mol. Cancer Ther. 2011, 10, 1644–1655. [Google Scholar] [CrossRef] [PubMed]
- Hamid, I.J.A.; Azman, N.A.; Gennery, A.R.; Mangantig, E.; Hashim, I.F.; Zainudeen, Z.T. Systematic Review of Primary Immunodeficiency Diseases in Malaysia: 1979–2020. Front. Immunol. 2020, 11, 1923. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Sehgal, S. Editorial: Advances in Primary Immunodeficiencies in India. Front. Immunol. 2021, 12, 1335. [Google Scholar] [CrossRef] [PubMed]
- Al-Herz, W.; Bousfiha, A.; Casanova, J.-L.; Chatila, T.; Conley, M.E.; Cunningham-Rundles, C.; Etzioni, A.; Franco, J.L.; Gaspar, H.B.; Holland, S.M.; et al. Primary Immunodeficiency Diseases: An Update on the Classification from the International Union of Immunological Societies Expert Committee for Primary Immunodeficiency. Front. Immunol. 2014, 5, 162. [Google Scholar] [CrossRef]
- Picard, C.; Bobby Gaspar, H.; Al-Herz, W.; Bousfiha, A.; Casanova, J.L.; Chatila, T.; Crow, Y.J.; Cunningham-Rundles, C.; Etzioni, A.; Franco, J.L.; et al. International Union of Immunological Societies: 2017 Primary Immunodeficiency Diseases Committee Report on Inborn Errors of Immunity. J. Clin. Immunol. 2018, 38, 96–128. [Google Scholar] [CrossRef]
- Tangye, S.G.; Al-Herz, W.; Bousfiha, A.; Chatila, T.; Cunningham-Rundles, C.; Etzioni, A.; Franco, J.L.; Holland, S.M.; Klein, C.; Morio, T.; et al. Human Inborn Errors of Immunity: 2019 Update on the Classification from the International Union of Immunological Societies Expert Committee. J. Clin. Immunol. 2020, 40, 24–64. [Google Scholar] [CrossRef]
- Abolhassani, H.; Azizi, G.; Sharifi, L.; Yazdani, R.; Mohsenzadegan, M.; Delavari, S.; Sohani, M.; Shirmast, P.; Chavoshzadeh, Z.; Mahdaviani, S.A.; et al. Global systematic review of primary immunodeficiency registries. Expert Rev. Clin. Immunol. 2020, 16, 717–732. [Google Scholar] [CrossRef]
- Bousfiha, A.; Jeddane, L.; Picard, C.; Al-Herz, W.; Ailal, F.; Chatila, T.; Cunningham-Rundles, C.; Etzioni, A.; Franco, J.; Holland, S.M.; et al. Human Inborn Errors of Immunity: 2019 Update of the IUIS Phenotypical Classification. J. Clin. Immunol. 2020, 40, 66–81. [Google Scholar] [CrossRef] [PubMed]
- Costagliola, G.; Cappelli, S.; Consolini, R. Autoimmunity in Primary Immunodeficiency Disorders: An Updated Review on Pathogenic and Clinical Implications. J. Clin. Med. 2021, 10, 4729. [Google Scholar] [CrossRef] [PubMed]
- McCusker, C.; Upton, J.; Warrington, R. Primary immunodeficiency. Allergy Asthma Clin. Immunol. 2018, 14, 61. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.-J.; Wang, Y.-C.; Liu, X.-M. Clinical and Immunological Features of Common Variable Immunodeficiency in China. Chin. Med. J. 2015, 128, 310–315. [Google Scholar] [CrossRef]
- Wu, J.; Zhong, W.; Yin, Y.; Zhang, H. Primary immunodeficiency disease: A retrospective study of 112 Chinese children in a single tertiary care center. BMC Pediatr. 2019, 19, 410–417. [Google Scholar] [CrossRef]
- Oliveira, J.B.; Fleisher, T.A. Laboratory evaluation of primary immunodeficiencies. J. Allergy Clin. Immunol. 2010, 125, S297–S305. [Google Scholar] [CrossRef]
- Heimall, J.R.; Hagin, D.; Hajjar, J.; Henrickson, S.; Hernandez-Trujillo, H.S.; Tan, Y.; Kobrynski, L.; Paris, K.; Torgerson, T.R.; Verbsky, J.W.; et al. Use of Genetic Testing for Primary Immunodeficiency Patients. J. Clin. Immunol. 2018, 38, 320–329. [Google Scholar] [CrossRef]
- Bacchetta, R.; Barzaghi, F.; Roncarolo, M.-G. From IPEX syndrome to FOXP3 mutation: A lesson on immune dysregulation. Ann. N. Y. Acad. Sci. 2016, 1417, 5–22. [Google Scholar] [CrossRef]
- Sakaguchi, S.; Yamaguchi, T.; Nomura, T.; Ono, M. Regulatory T Cells and Immune Tolerance. Cell 2008, 133, 775–787. [Google Scholar] [CrossRef]
- Gavin, M.A.; Rasmussen, J.; Fontenot, J.D.; Vasta, V.; Manganiello, V.C.; Beavo, J.A.; Rudensky, A.Y. Foxp3-dependent programme of regulatory T-cell differentiation. Nature 2007, 445, 771–775. [Google Scholar] [CrossRef]
- Lin, Y.-L.; Shieh, C.-C.; Wang, J.-Y. The functional insufficiency of human CD4+CD25high T-regulatory cells in allergic asthma is subjected to TNF-α modulation. Allergy 2007, 63, 67–74. [Google Scholar] [CrossRef]
- Fontenot, J.D.; Gavin, M.A.; Rudensky, A.Y. Foxp3 programs the development and function of CD4+CD25+ regulatory T cells. Nat. Immunol. 2003, 4, 330–336. [Google Scholar] [CrossRef]
- Hori, S.; Nomura, T.; Sakaguchi, S. Control of Regulatory T Cell Development by the Transcription Factor Foxp3. Science 2003, 299, 1057–1061. [Google Scholar] [CrossRef] [PubMed]
- Khattri, R.; Cox, T.; Yasayko, S.-A.; Ramsdell, F. An essential role for Scurfin in CD4+CD25+ T regulatory cells. Nat. Immunol. 2003, 4, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Hu, B.; Xu, D.; Liew, F.Y. CD4+CD25+ Regulatory T Cells Cure Murine Colitis: The Role of IL-10, TGF-β, and CTLA4. J. Immunol. 2003, 171, 5012–5017. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, M.-T.J.; Fryml, E.; Sahakian, S.K.; Liu, S.; Michel, R.P.; Lipman, M.L.; Mucsi, I.; Cantarovich, M.; Tchervenkov, J.I.; Paraskevas, S. Pretransplantation Recipient Regulatory T cell Suppressive Function Predicts Delayed and Slow Graft Function after Kidney Transplantation. Transplantation 2014, 98, 745–753. [Google Scholar] [CrossRef]
- Palomares, O.; Yaman, G.; Azkur, A.K.; Akkoc, T.; Akdis, M.; Akdis, C.A. Role of Treg in immune regulation of allergic diseases. Eur. J. Immunol. 2010, 40, 1232–1240. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Wing, J.B.; Sakaguchi, S. Two modes of immune suppression by Foxp3+ regulatory T cells under inflammatory or non-inflammatory conditions. Semin. Immunol. 2011, 23, 424–430. [Google Scholar] [CrossRef]
- Kwon, H.; Chen, H.-M.; Mathis, D.; Benoist, C. FoxP3 scanning mutagenesis reveals functional variegation and mild mutations with atypical autoimmune phenotypes. Proc. Natl. Acad. Sci. USA 2017, 115, E253–E262. [Google Scholar] [CrossRef]
- Van Gool, F.; Nguyen, M.L.; Mumbach, M.R.; Satpathy, A.T.; Rosenthal, W.L.; Giacometti, S.; Le, D.T.; Liu, W.; Brusko, T.M.; Anderson, M.S.; et al. A Mutation in the Transcription Factor Foxp3 Drives T Helper 2 Effector Function in Regulatory T Cells. Immunity 2019, 50, 362–377.e6. [Google Scholar] [CrossRef]
- Van Der Vliet, H.J.J.; Nieuwenhuis, E.E. IPEX as a Result of Mutations in FOXP3. Clin. Dev. Immunol. 2007, 2007, e89017. [Google Scholar] [CrossRef]
- IPEX: Immune Dysregulation, Polyendocrinopathy, Enteropathy, X-Linked-UpToDate. Available online: https://www.uptodate.com/contents/ipex-immune-dysregulation-polyendocrinopathy-enteropathy-x-linked (accessed on 30 December 2021).
- Hines, B.; Wright, B.L.; Wadera, S.; Cac, N.; Miller, H.K.; Abraham, R.S.; Bauer, C.S. IPEX Syndrome in Siblings with a Novel Variant in FOXP3. J. Allergy Clin. Immunol. 2019, 143, AB118. [Google Scholar] [CrossRef]
- Ozcan, E.; Notarangelo, L.D.; Geha, R.S. Primary immune deficiencies with aberrant IgE production. J. Allergy Clin. Immunol. 2008, 122, 1054–1062. [Google Scholar] [CrossRef] [PubMed]
- Torgerson, T.R.; Ochs, H.D. Immune dysregulation, polyendocrinopathy, enteropathy, X-linked: Forkhead box protein 3 mutations and lack of regulatory T cells. J. Allergy Clin. Immunol. 2007, 120, 744–750. [Google Scholar] [CrossRef]
- Torgerson, T.R.; Linane, A.; Moes, N.; Anover, S.; Mateo, V.; Rieux-Laucat, F.; Hermine, O.; Vijay, S.; Gambineri, E.; Cerf-Bensussan, N.; et al. Severe Food Allergy as a Variant of IPEX Syndrome Caused by a Deletion in a Noncoding Region of the FOXP3 Gene. Gastroenterology 2007, 132, 1705–1717. [Google Scholar] [CrossRef] [PubMed]
- Lucas, K.G.; Ungar, D.; Comito, M.; Bayerl, M.; Groh, B. Submyeloablative cord blood transplantation corrects clinical defects seen in IPEX syndrome. Bone Marrow Transpl. 2006, 39, 55–56. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Chatila, T.A.; Blaeser, F.; Ho, N.; Lederman, H.M.; Voulgaropoulos, C.; Helms, C.; Bowcock, A. JM2, encoding a fork head–related protein, is mutated in X-linked autoimmunity–allergic disregulation syndrome. J. Clin. Investig. 2000, 106, R75–R81. [Google Scholar] [CrossRef]
- Heltzer, M.L.; Choi, J.K.; Ochs, H.D.; Sullivan, K.E.; Torgerson, T.R.; Ernst, L. A Potential Screening Tool for IPEX Syndrome. Pediatr. Dev. Pathol. 2007, 10, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Owen, C.J.; Jennings, C.E.; Imrie, H.; Lachaux, A.; Bridges, N.A.; Cheetham, T.D.; Pearce, S. Mutational Analysis of the FOXP3 Gene and Evidence for Genetic Heterogeneity in the Immunodysregulation, Polyendocrinopathy, Enteropathy Syndrome. J. Clin. Endocrinol. Metab. 2003, 88, 6034–6039. [Google Scholar] [CrossRef] [PubMed]
- Ochs, H.D.; Torgerson, T.R. Immune dysregulation, polyendocrinopathy, enteropathy, X-linked inheritance: Model for autoaggression. Adv. Exp. Med. Biol. 2007, 601, 27–36. [Google Scholar] [CrossRef]
- Gambineri, E.; Perroni, L.; Passerini, L.; Bianchi, L.; Doglioni, C.; Meschi, F.; Bonfanti, R.; Sznajer, Y.; Tommasini, A.; Lawitschka, A.; et al. Clinical and molecular profile of a new series of patients with immune dysregulation, polyendocrinopathy, enteropathy, X-linked syndrome: Inconsistent correlation between forkhead box protein 3 expression and disease severity. J. Allergy Clin. Immunol. 2008, 122, 1105–1112.e1. [Google Scholar] [CrossRef] [PubMed]
- Barzaghi, F.; Passerini, L.; Bacchetta, R. Immune Dysregulation, Polyendocrinopathy, Enteropathy, X-Linked Syndrome: A Paradigm of Immunodeficiency with Autoimmunity. Front. Immunol. 2012, 3, 211. [Google Scholar] [CrossRef] [PubMed]
- Bacchetta, R.; Passerini, L.; Gambineri, E.; Dai, M.; Allan, S.E.; Perroni, L.; Dagna-Bricarelli, F.; Sartirana, C.; Matthes-Martin, S.; Lawitschka, A.; et al. Defective regulatory and effector T cell functions in patients with FOXP3 mutations. J. Clin. Investig. 2006, 116, 1713–1722. [Google Scholar] [CrossRef] [PubMed]
- Fuchizawa, T.; Adachi, Y.; Ito, Y.; Higashiyama, H.; Kanegane, H.; Futatani, T.; Kobayashi, I.; Kamachi, Y.; Sakamoto, T.; Tsuge, I.; et al. Developmental changes of FOXP3-expressing CD4+CD25+ regulatory T cells and their impairment in patients with FOXP3 gene mutations. Clin. Immunol. 2007, 125, 237–246. [Google Scholar] [CrossRef]
- Walker, M.R.; Kasprowicz, D.J.; Gersuk, V.H.; Bénard, A.; Van Landeghen, M.; Buckner, J.H.; Ziegler, S.F. Induction of FoxP3 and acquisition of T regulatory activity by stimulated human CD4+CD25− T cells. J. Clin. Investig. 2003, 112, 1437–1443. [Google Scholar] [CrossRef]
- Liu, W.; Putnam, A.L.; Xu-Yu, Z.; Szot, G.L.; Lee, M.R.; Zhu, S.; Gottlieb, P.A.; Kapranov, P.; Gingeras, T.R.; de St Groth, B.F.; et al. CD127 expression inversely correlates with FoxP3 and suppressive function of human CD4+ T reg cells. J. Exp. Med. 2006, 203, 1701–1711. [Google Scholar] [CrossRef]
- Seddiki, N.; Santner-Nanan, B.; Martinson, J.; Zaunders, J.; Sasson, S.; Landay, A.; Solomon, M.; Selby, W.; Alexander, S.I.; Nanan, R.; et al. Expression of interleukin (IL)-2 and IL-7 receptors discriminates between human regulatory and activated T cells. J. Exp. Med. 2006, 203, 1693–1700. [Google Scholar] [CrossRef]
- Alves, N.L.; Van Leeuwen, E.M.M.; Derks, I.A.M.; van Lier, R. Differential Regulation of Human IL-7 Receptor α Expression by IL-7 and TCR Signaling. J. Immunol. 2008, 180, 5201–5210. [Google Scholar] [CrossRef]
- Otsubo, K.; Kanegane, H.; Kamachi, Y.; Kobayashi, I.; Tsuge, I.; Imaizumi, M.; Sasahara, Y.; Hayakawa, A.; Nozu, K.; Iijima, K.; et al. Identification of FOXP3-negative regulatory T-like (CD4+CD25+CD127low) cells in patients with immune dysregulation, polyendocrinopathy, enteropathy, X-linked syndrome. Clin. Immunol. 2011, 141, 111–120. [Google Scholar] [CrossRef]
- Imamichi, H.; Sereti, I.; Lane, H.C. IL-15 acts as a potent inducer of CD4+CD25hi cells expressing FOXP3. Eur. J. Immunol. 2008, 38, 1621–1630. [Google Scholar] [CrossRef]
- Tabares, P.; Berr, S.; Langenhorst, D.; Sawitzki, B.; Berge, I.T.; Tony, H.-P.; Hünig, T. Short-term cytokine stimulation reveals regulatory T cells with down-regulated Foxp3 expression in human peripheral blood. Eur. J. Immunol. 2017, 48, 366–379. [Google Scholar] [CrossRef] [PubMed]
- Allan, S.E.; Passerini, L.; Bacchetta, R.; Crellin, N.; Dai, M.; Orban, P.C.; Ziegler, S.F.; Roncarolo, M.G.; Levings, M.K. The role of 2 FOXP3 isoforms in the generation of human CD4+ Tregs. J. Clin. Investig. 2005, 115, 3276–3284. [Google Scholar] [CrossRef] [PubMed]
- Aarts-Riemens, T.; Emmelot, M.E.; Verdonck, L.F.; Mutis, T. Forced overexpression of either of the two common human Foxp3 isoforms can induce regulatory T cells from CD4+CD25− cells. Eur. J. Immunol. 2008, 38, 1381–1390. [Google Scholar] [CrossRef] [PubMed]
- Allan, S.E.; Crome, S.; Crellin, N.K.; Passerini, L.; Steiner, T.; Bacchetta, R.; Roncarolo, M.G.; Levings, M. Activation-induced FOXP3 in human T effector cells does not suppress proliferation or cytokine production. Int. Immunol. 2007, 19, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ioan-Facsinay, A.; van der Voort, E.I.H.; Huizinga, T.W.J.; Toes, R.E.M. Transient expression of FOXP3 in human activated nonregulatory CD4+ T cells. Eur. J. Immunol. 2006, 37, 129–138. [Google Scholar] [CrossRef]
- Passerini, L.; Mel, E.R.; Sartirana, C.; Fousteri, G.; Bondanza, A.; Naldini, L.; Roncarolo, M.G.; Bacchetta, R. CD4+ T Cells from IPEX Patients Convert into Functional and Stable Regulatory T Cells by FOXP3 Gene Transfer. Sci. Transl. Med. 2013, 5, 215ra174. [Google Scholar] [CrossRef]
- Jamee, M.; Zaki-Dizaji, M.; Lo, B.; Abolhassani, H.; Aghamahdi, F.; Mosavian, M.; Nademi, Z.; Mohammadi, H.; Jadidi-Niaragh, F.; Rojas, M.; et al. Clinical, Immunological, and Genetic Features in Patients with Immune Dysregulation, Polyendocrinopathy, Enteropathy, X-linked (IPEX) and IPEX-like Syndrome. J. Allergy Clin. Immunol. Pract. 2020, 8, 2747–2760.e7. [Google Scholar] [CrossRef]
- Varricchi, G.; Poto, R.; Ianiro, G.; Punziano, A.; Marone, G.; Gasbarrini, A.; Spadaro, G. Gut Microbiome and Common Variable Immunodeficiency: Few Certainties and Many Outstanding Questions. Front. Immunol. 2021, 12, 2915. [Google Scholar] [CrossRef]
- NORD (National Organization for Rare Disorders). Common Variable Immune Deficiency. Available online: https://rarediseases.org/rare-diseases/common-variable-immune-deficiency (accessed on 30 December 2021).
- Yakaboski, E.; Fuleihan, R.L.; Sullivan, K.E.; Cunningham-Rundles, C.; Feuille, E. Lymphoproliferative Disease in CVID: A Report of Types and Frequencies from a US Patient Registry. J. Clin. Immunol. 2020, 40, 524–530. [Google Scholar] [CrossRef]
- Gathmann, B.; Mahlaoui, N.; Gérard, L.; Oksenhendler, E.; Warnatz, K.; Schulze, I.; Kindle, G.; Kuijpers, T.W.; van Beem, R.T.; Guzman, D.; et al. Clinical picture and treatment of 2212 patients with common variable immunodeficiency. J. Allergy Clin. Immunol. 2014, 134, 116–126.e11. [Google Scholar] [CrossRef]
- Pathogenesis of Common Variable Immunodeficiency-UpToDate. Available online: https://www.uptodate.com/contents/pathogenesis-of-common-variable-immunodeficiency (accessed on 30 December 2021).
- Vořechovský, I.; Cullen, M.; Carrington, M.; Hammarström, L.; Webster, A.D.B. Fine Mapping of IGAD1 in IgA Deficiency and Common Variable Immunodeficiency: Identification and Characterization of Haplotypes Shared by Affected Members of 101 Multiple-Case Families. J. Immunol. 2000, 164, 4408–4416. [Google Scholar] [CrossRef]
- Patuzzo, G.; Mazzi, F.; Vella, A.; Ortolani, R.; Barbieri, A.; Tinazzi, E.; Marchi, G.; Codella, O.; Beri, R.; Puccetti, A.; et al. Immunophenotypic Analysis of B Lymphocytes in Patients with Common Variable Immunodeficiency: Identification of CD23 as a Useful Marker in the Definition of the Disease. ISRN Immunol. 2013, 2013, 512527. [Google Scholar] [CrossRef]
- Matson, E.M.; Abyazi, M.L.; Bell, K.A.; Hayes, K.M.; Maglione, P.J. B Cell Dysregulation in Common Variable Immunodeficiency Interstitial Lung Disease. Front. Immunol. 2021, 11, 3842. [Google Scholar] [CrossRef]
- López-Herrera, G.; Segura-Méndez, N.H.; O’Farril-Romanillos, P.; Nez, M.E.N.-N.; Zarate-Hernández, M.C.; Mogica-Martínez, D.; Yamazaki-Nakashimada, M.A.; Staines-Boone, A.T.; Santos-Argumedo, L.; Berrón-Ruiz, L. Low Percentages of Regulatory T Cells in Common Variable Immunodeficiency (CVID) Patients with Autoimmune Diseases and Its Association with Increased Numbers of CD4+CD45RO+ T and CD21 Low B Cells. Allergol. Immunopathol. 2019, 47, 457–466. [Google Scholar] [CrossRef]
- Ghafoor, A.; Joseph, S.M. Making a Diagnosis of Common Variable Immunodeficiency: A Review. Cureus 2020, 12, e6711. [Google Scholar] [CrossRef]
- Salzer, U.; Warnatz, K.; Peter, H.H. Common variable immunodeficiency-an update. Arthritis Res. Ther. 2012, 14, 223. [Google Scholar] [CrossRef] [PubMed]
- Kutukculer, N.; Azarsiz, E.; Aksu, G.; Karaca, N.E. CD4+CD25+Foxp3+ T regulatory cells, Th1 (CCR5, IL-2, IFN-γ) and Th2 (CCR4, IL-4, Il-13) type chemokine receptors and intracellular cytokines in children with common variable immunodeficiency. Int. J. Immunopathol. Pharmacol. 2015, 29, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Horn, J.; Manguiat, A.; Berglund, L.J.; Knerr, V.; Tahami, F.; Grimbacher, B.; Fulcher, D.A. Decrease in phenotypic regulatory T cells in subsets of patients with common variable immunodeficiency. Clin. Exp. Immunol. 2009, 156, 446–454. [Google Scholar] [CrossRef] [PubMed]
- Genre, J.; Errante, P.R.; Kokron, C.M.; Toledo-Barros, M.; Câmara, N.; Rizzo, L.V. Reduced frequency of CD4+CD25HIGHFOXP3+ cells and diminished FOXP3 expression in paO.S.tients with Common Variable Immunodeficiency: A link to autoimmunity? Clin. Immunol. 2009, 132, 215–221. [Google Scholar] [CrossRef]
- Arandi, N.; Mirshafiey, A.; Abolhassani, H.; Jeddi-Tehrani, M.; Edalat, R.; Sadeghi, B.; Shaghaghi, M.; Aghamohammadi, A. Frequency and Expression of Inhibitory Markers of CD4+CD25+FOXP3+Regulatory T Cells in Patients with Common Variable Immunodeficiency. Scand. J. Immunol. 2013, 77, 405–412. [Google Scholar] [CrossRef]
- UniProtKB. Available online: http://www.uniprot.org (accessed on 30 December 2021).
- Kozlowski, L.P. IPC–Isoelectric Point Calculator. Biol. Direct 2016, 11, 55. [Google Scholar] [CrossRef] [PubMed]
- Services. Available online: https://services.healthtech.dtu.dk (accessed on 30 December 2021).
- Align. Available online: https://www.uniprot.org/align (accessed on 30 December 2021).






| Name | Amino Acid Length | Mass [kDa] | Isoelectric Point | Amino Acid Composition | Secondary Structure | Protein ID | ||
|---|---|---|---|---|---|---|---|---|
| Hydrophobic Amino Acids [%] | Hydrophilic Amino Acids [%] | α-Helix | β-Starnad | |||||
| Foxp3 | 431 | 47.24 | 8.62 | 53.60 | 46.40 | 12 (26.15%) | 10 (6.26%) | Q9BZS1 |
| Isoform2 | 396 | 43.41 | 8.53 | 52.27 | 47.73 | 10 (25.25%) | 9 (5.55%) | Q9BZS1-2 |
| Isoform 3 | 456 | 49.84 | 8.00 | 53.51 | 46.49 | 14 (27.41%) | 10 (6.56%) | Q9BZS1-3 |
| Isoform 4 | 404 | 44.41 | 8.52 | 53.47 | 46.53 | 12 (24.50%) | 11 (7.43%) | Q9BZS1-4 |
| Foxp3 | Isoform 2 | Isoform 3 | Isoform 4 | |
|---|---|---|---|---|
| Foxp3 | - | 91.88% | 80.65% | 93.73% |
| Isoform 2 | 91.88% | - | 86.84% | 85.61% |
| Isoform 3 | 80.65% | 86.84% | - | 75.15% |
| Isoform 4 | 93.73% | 85.61% | 75.15% | - |
| Name of the Subpopulation | Characteristics | |
|---|---|---|
| Expressing Foxp3 | T lymphocytes CD4+CD25+Foxp3+ | The most widely studied and characterized sub-population of regulatory cells. Characterized by the expression of the Foxp3 transcription factor and accompanied by a high expression of the CD25 surface molecule. |
| T lymphocytes CD8+CD25+Foxp3+ | A subpopulation of CD8+ T cells that are much less well understood than CD4+Foxp3+ cells. | |
| Not expressing Foxp3 | Type 1 regulatory T cells (Tr1) | Cells with the CD4+Foxp3− phenotype secreting significant amounts of IL-10. |
| Th3 lymphocytes | CD4+Foxp3− cells secreting significant amounts of TGF-β. | |
| CD8+CD28− lymphocytes | identified with pre-Ts lymphocytes that also do not express Foxp3. | |
| Type of Research | Type of Examination | The Importance of the Examination | Reference |
|---|---|---|---|
| Basic examination | Complete blood count with a smear | Presence of eosinophilia, neutropenia, anemia, or thrombocytopenia | [177] |
| Serum glucose concentration | Glucose monitoring can help to detect the presence of type 1 diabetes | [177] | |
| Functioning of the thyroid gland | Elevated levels of anti-thyroid antibodies | [177] | |
| Concentration of immunoglobulins | Increase in IgE level in most patients, increase in IgA level in half of the patients, normal IgG and IgM | [178,179,180,181,182] | |
| Food hypersensitivity test | Presence of IgE-dependent food allergy | [181,183] | |
| Determination of the percentage of T and B lymphocytes | T and B cell subsets are usually normal | [177] | |
| Specialized research | Endoscopy with intestinal biopsy | Necessary to characterize the presence of enteropathy of the small intestine, combined with the performance of Foxp3 staining | [184] |
| Skin biopsy | The presence of lymphocyte infiltrates in biopsy samples as an autoimmune process | [177] | |
| Treg lymphocyte immunophenotyping | Determines the amount of Treg and the expression level of Foxp3 | [177] | |
| Sequencing of the FOXP3 gene | Evaluates mutations within the FOXP3 gene to confirm the clinical picture | [177] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mertowska, P.; Mertowski, S.; Podgajna, M.; Grywalska, E. The Importance of the Transcription Factor Foxp3 in the Development of Primary Immunodeficiencies. J. Clin. Med. 2022, 11, 947. https://doi.org/10.3390/jcm11040947
Mertowska P, Mertowski S, Podgajna M, Grywalska E. The Importance of the Transcription Factor Foxp3 in the Development of Primary Immunodeficiencies. Journal of Clinical Medicine. 2022; 11(4):947. https://doi.org/10.3390/jcm11040947
Chicago/Turabian StyleMertowska, Paulina, Sebastian Mertowski, Martyna Podgajna, and Ewelina Grywalska. 2022. "The Importance of the Transcription Factor Foxp3 in the Development of Primary Immunodeficiencies" Journal of Clinical Medicine 11, no. 4: 947. https://doi.org/10.3390/jcm11040947
APA StyleMertowska, P., Mertowski, S., Podgajna, M., & Grywalska, E. (2022). The Importance of the Transcription Factor Foxp3 in the Development of Primary Immunodeficiencies. Journal of Clinical Medicine, 11(4), 947. https://doi.org/10.3390/jcm11040947