
The Effect of Aerobic Exercise and Low-Impact Pilates Workout on the Adaptive Immune System
 , , ,
, , , 

Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Exercise Protocols
2.3. Blood Sampling
2.4. Assessment of Lymphocyte Subpopulations by Flow Cytometry
2.5. Determination of T Helper and T Cytotoxic Cells by Immunofluorescent Staining of Intracellular Cytokines
2.6. Statistical Analysis
3. Results
3.1. Data Describing Study Population
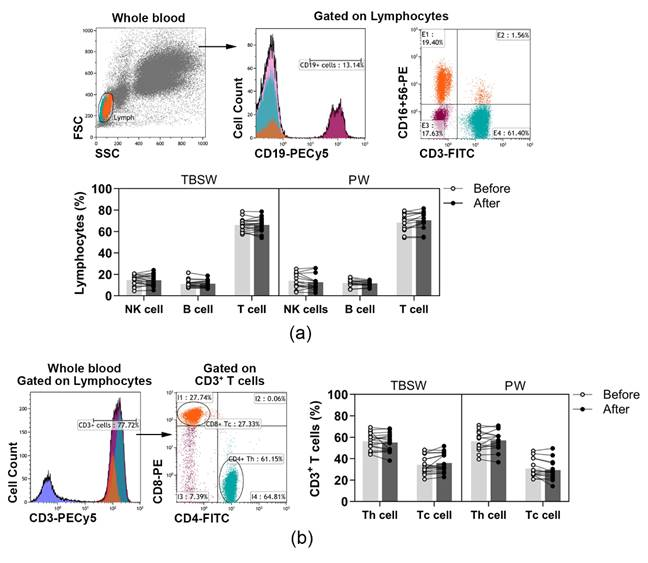
3.2. Quantification of Different Lymphocyte Subpopulations before and after the Exercise Routine
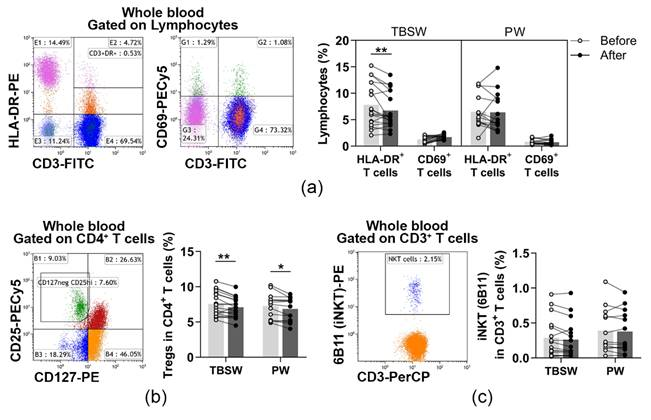
3.3. Determination of Cells with a Regulatory Function in the Innate and Adaptive Immune Response after a 14-Week Exercise Routine
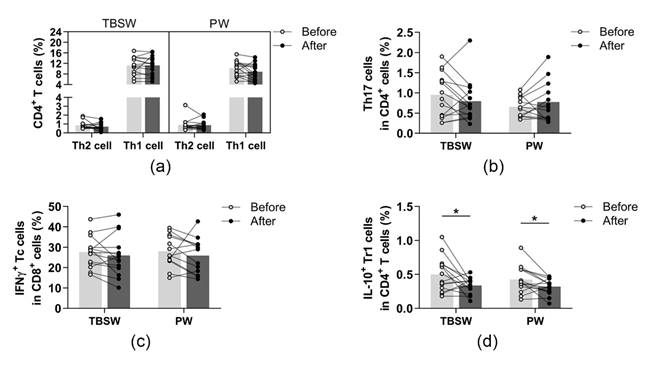
3.4. Assessment of Peripheral T Helper Subsets and Cytotoxic T Cells before and after the 14-Week Training
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Booth, F.W.; Roberts, C.K.; Thyfault, J.P.; Ruegsegger, G.N.; Toedebusch, R.G. Role of Inactivity in Chronic Diseases: Evolutionary Insight and Pathophysiological Mechanisms. Physiol. Rev. 2017, 97, 1351–1402. [Google Scholar] [CrossRef] [PubMed]
- Laaksonen, D.E.; Lakka, H.M.; Salonen, J.T.; Niskanen, L.K.; Rauramaa, R.; Lakka, T.A. Low levels of leisure-time physical activity and cardiorespiratory fitness predict development of the metabolic syndrome. Diabetes Care 2002, 25, 1612–1618. [Google Scholar] [CrossRef] [PubMed]
- Warburton, D.E.R.; Bredin, S.S.D. Health benefits of physical activity: A systematic review of current systematic reviews. Curr. Opin. Cardiol. 2017, 32, 541–556. [Google Scholar] [CrossRef]
- Sharif, K.; Watad, A.; Bragazzi, N.L.; Lichtbroun, M.; Amital, H.; Shoenfeld, Y. Physical activity and autoimmune diseases: Get moving and manage the disease. Autoimmun. Rev. 2018, 17, 53–72. [Google Scholar] [CrossRef] [PubMed]
- Pape, K.; Ryttergaard, L.; Rotevatn, T.A.; Nielsen, B.J.; Torp-Pedersen, C.; Overgaard, C.; Boggild, H. Leisure-Time Physical Activity and the Risk of Suspected Bacterial Infections. Med. Sci. Sports Exerc. 2016, 48, 1737–1744. [Google Scholar] [CrossRef] [PubMed]
- Kostka, T.; Berthouze, S.E.; Lacour, J.; Bonnefoy, M. The symptomatology of upper respiratory tract infections and exercise in elderly people. Med. Sci. Sports Exerc. 2000, 32, 46–51. [Google Scholar] [CrossRef]
- McCarthy, D.A.; Dale, M.M. The leucocytosis of exercise. A review and model. Sports Med. 1988, 6, 333–363. [Google Scholar] [CrossRef]
- Navalta, J.W.; Sedlock, D.A.; Park, K.S. Effect of exercise intensity on exercise-induced lymphocyte apoptosis. Int. J. Sports Med. 2007, 28, 539–542. [Google Scholar] [CrossRef]
- Steensberg, A.; Toft, A.D.; Bruunsgaard, H.; Sandmand, M.; Halkjaer-Kristensen, J.; Pedersen, B.K. Strenuous exercise decreases the percentage of type 1 T cells in the circulation. J. Appl. Physiol. 2001, 91, 1708–1712. [Google Scholar] [CrossRef]
- Pedersen, B.K.; Hoffman-Goetz, L. Exercise and the immune system: Regulation, integration, and adaptation. Physiol. Rev. 2000, 80, 1055–1081. [Google Scholar] [CrossRef]
- Elenkov, I.J.; Chrousos, G.P. Stress hormones, Th1/Th2 patterns, pro/anti-inflammatory cytokines, and susceptibility to disease. Trends Endocrinol. Metab. 1999, 10, 359–368. [Google Scholar] [CrossRef]
- Kruger, K.; Lechtermann, A.; Fobker, M.; Volker, K.; Mooren, F.C. Exercise-induced redistribution of T lymphocytes is regulated by adrenergic mechanisms. Brain Behav. Immun. 2008, 22, 324–338. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.P.; Turner, J.E. Debunking the Myth of Exercise-Induced Immune Suppression: Redefining the Impact of Exercise on Immunological Health Across the Lifespan. Front. Immunol. 2018, 9, 648. [Google Scholar] [CrossRef]
- Campbell, J.P.; Turner, J.E. There is limited existing evidence to support the common assumption that strenuous endurance exercise bouts impair immune competency. Expert Rev. Clin. Immunol. 2019, 15, 105–109. [Google Scholar] [CrossRef] [PubMed]
- Nieman, D.C.; Wentz, L.M. The compelling link between physical activity and the body’s defense system. J. Sport Health Sci. 2019, 8, 201–217. [Google Scholar] [CrossRef]
- Simpson, R.J.; Campbell, J.P.; Gleeson, M.; Krüger, K.; Nieman, D.C.; Pyne, D.B.; Turner, J.E.; Walsh, N.P. Can exercise affect immune function to increase susceptibility to infection? Exerc. Immunol. Rev. 2020, 26, 8–22. [Google Scholar]
- Simpson, R.J.; Kunz, H.; Agha, N.; Graff, R. Exercise and the Regulation of Immune Functions. Prog. Mol. Biol. Transl. Sci 2015, 135, 355–380. [Google Scholar] [CrossRef]
- Woods, J.A.; Keylock, K.T.; Lowder, T.; Vieira, V.J.; Zelkovich, W.; Dumich, S.; Colantuano, K.; Lyons, K.; Leifheit, K.; Cook, M.; et al. Cardiovascular exercise training extends influenza vaccine seroprotection in sedentary older adults: The immune function intervention trial. J. Am. Geriatr. Soc. 2009, 57, 2183–2191. [Google Scholar] [CrossRef]
- Simpson, R.J. Aging, persistent viral infections, and immunosenescence: Can exercise “make space”? Exerc. Sport Sci. Rev. 2011, 39, 23–33. [Google Scholar] [CrossRef]
- Hagner-Derengowska, M.; Kałużny, K.; Kochański, B.; Hagner, W.; Borkowska, A.; Czamara, A.; Budzyński, J. Effects of Nordic Walking and Pilates exercise programs on blood glucose and lipid profile in overweight and obese postmenopausal women in an experimental, nonrandomized, open-label, prospective controlled trial. Menopause 2015, 22, 1215–1223. [Google Scholar] [CrossRef]
- Pilates, J.H.; Miller, W.J. Pilates’ Return to Life through Contrology; Presentation Dynamics Inc.: Incline Village, NV, USA, 1998. [Google Scholar]
- Gronesova, P.; Cholujova, D.; Kozic, K.; Korbuly, M.; Vlcek, M.; Penesova, A.; Imrich, R.; Sedlak, J.; Hunakova, L. Effects of short-term Pilates exercise on selected blood parameters. Gen. Physiol. Biophys. 2018, 37, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Wilson, L.D.; Zaldivar, F.P.; Schwindt, C.D.; Wang-Rodriguez, J.; Cooper, D.M. Circulating T-regulatory cells, exercise and the elite adolescent swimmer. Pediatr. Exerc. Sci. 2009, 21, 305–317. [Google Scholar] [CrossRef]
- Hulmi, J.J.; Myllymäki, T.; Tenhumäki, M.; Mutanen, N.; Puurtinen, R.; Paulsen, G.; Mero, A.A. Effects of resistance exercise and protein ingestion on blood leukocytes and platelets in young and older men. Eur. J. Appl. Physiol. 2010, 109, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Hwang, Y.; Park, J.; Lim, K. Effects of Pilates Exercise on Salivary Secretory Immunoglobulin A Levels in Older Women. J. Aging Phys. Act. 2016, 24, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Sellami, M.; Bragazzi, N.L.; Aboghaba, B.; Elrayess, M.A. The Impact of Acute and Chronic Exercise on Immunoglobulins and Cytokines in Elderly: Insights From a Critical Review of the Literature. Front. Immunol. 2021, 12, 631873. [Google Scholar] [CrossRef] [PubMed]
- Martins, R.A.; Cunha, M.R.; Neves, A.P.; Martins, M.; Teixeira-Veríssimo, M.; Teixeira, A.M. Effects of aerobic conditioning on salivary IgA and plasma IgA, IgG and IgM in older men and women. Int. J. Sports Med. 2009, 30, 906–912. [Google Scholar] [CrossRef] [PubMed]
- Kogut, I.; Scholz, J.L.; Cancro, M.P.; Cambier, J.C. B cell maintenance and function in aging. Semin. Immunol. 2012, 24, 342–349. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.E.; Spielmann, G.; Wadley, A.J.; Aldred, S.; Simpson, R.J.; Campbell, J.P. Exercise-induced B cell mobilisation: Preliminary evidence for an influx of immature cells into the bloodstream. Physiol. Behav. 2016, 164, 376–382. [Google Scholar] [CrossRef]
- Palm, A.E.; Henry, C. Remembrance of Things Past: Long-Term B Cell Memory after Infection and Vaccination. Front. Immunol. 2019, 10, 1787. [Google Scholar] [CrossRef]
- Papp, G.; Szabó, K.; Jámbor, I.; Mile, M.; Berki, A.R.; Arany, A.C.; Makra, G.; Szodoray, P.; Csiki, Z.; Balogh, L. Regular Exercise May Restore Certain Age-Related Alterations of Adaptive Immunity and Rebalance Immune Regulation. Front. Immunol. 2021, 12, 639308. [Google Scholar] [CrossRef]
- Mittelbrunn, M.; Kroemer, G. Hallmarks of T cell aging. Nat. Immunol. 2021, 22, 687–698. [Google Scholar] [CrossRef] [PubMed]
- Mathot, E.; Liberman, K.; Dinh, H.C.; Njemini, R.; Bautmans, I. Systematic review on the effects of physical exercise on cellular immunosenescence-related markers—An update. Exp. Gerontol. 2021, 149, 111318. [Google Scholar] [CrossRef] [PubMed]
- Arruvito, L.; Payaslián, F.; Baz, P.; Podhorzer, A.; Billordo, A.; Pandolfi, J.; Semeniuk, G.; Arribalzaga, E.; Fainboim, L. Identification and clinical relevance of naturally occurring human CD8 + HLA-DR+ regulatory T cells. J. Immunol. 2014, 193, 4469–4476. [Google Scholar] [CrossRef] [PubMed]
- Papp, G.; Boros, P.; Nakken, B.; Szodoray, P.; Zeher, M. Regulatory immune cells and functions in autoimmunity and transplantation immunology. Autoimmun. Rev. 2017, 16, 435–444. [Google Scholar] [CrossRef]
- Lowder, T.; Dugger, K.; Deshane, J.; Estell, K.; Schwiebert, L.M. Repeated bouts of aerobic exercise enhance regulatory T cell responses in a murine asthma model. Brain Behav. Immun. 2010, 24, 153–159. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibodies | Clone | Fluorophore | Company | Lymphocyte Subsets |
|---|---|---|---|---|
| CD19 | J3-119 | PE-Cy5 | Beckmann Coulter a | CD3+ T cells, |
| CD3/ | SK7/ | FITC | BD Biosciences b | CD3−CD16+CD56+ NK cells, |
| CD16+CD56 | B73.1+MY31 | PE | CD19+ B cells | |
| CD3/ CD4/ CD8 | UCHT1/ MT310/ DK25 | RPE-Cy5 FITC RPE | Bio-Rad c | CD3+CD4+ Th cells, CD3+CD8+ Tc cells |
| IgD CD27 CD19 | IA6-2 1A4CD27 J3-119 | FITC PE PE-Cy5 | Beckman Coulter | CD19+IgD+CD27− naïve, CD19+IgD+CD27+ UswM, CD19+IgD−CD27+ SwM, CD19+IgD−CD27− DN B cells |
| CD62L | DREG56 | PE-Cy5 | Beckman Coulter | CD45RA+CD62L+ naïve, |
| CD45RA/ | ALB11/ | FITC | CD45RA−CD62L+ CM, | |
| CD4 | 13B8.2 | PE | CD45RA−CD62L− EM, | |
| CD45RA/ | F8-11-13/ | FITC | Bio-Rad | CD45RA+CD62L− EMRA, |
| CD8 | LT8 | RPE | CD4+ Th and CD8+ Tc cells | |
| CD3/ HLA-DR CD69 | UCHT1/ WR18 FN50 | FITC RPE PE-Cy5 | BD Biosciences | CD3+CD69+ T cells, CD3+HLA-DR+ T cells |
| CD3 | SK7 | PerCP | BD Biosciences | CD3+6B11+ NKT cells |
| Inkt d | 6B11 | PE | ||
| CD4 | RPA-T4 | FITC | BD Biosciences | CD4+CD127lo/−CD25bright Treg cells |
| CD25 | B1.49.9 | PE-Cy5 | Beckman Coulter | |
| CD127 | R34.34 | PE |
| Antibodies | Clone | Fluorophore | Company | T Cell Subsets |
|---|---|---|---|---|
| CD4 | 13B8.2 | PE-Cy5 | Beckmann Coulter a | |
| IL-10 | JES3-19F1 | PE | BD Biosciences b | IFN-γ+IL-4− Th1 cells, |
| IFN-γ/ | 25723.11/ | FITC | IFN-γ-IL-4+ Th2 cells, | |
| IL-4 d | 3010.211 | PE | IL-10+ Tr1 cells, | |
| IL-17 | 41802 | PE | R&D Systems c | IFN-γ-IL17+ Th17 cells |
| CD8 | 4S.B3 | FITC | BD Biosciences | |
| CD8 | B9.11 | PE-Cy5 | Beckmann Coulter | IFN-γ+IL-4− Tc cells |
| IFN-γ/ | 25723.11/ | FITC | BD Biosciences | |
| IL-4 | 3010.211 | PE |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balogh, L.; Szabó, K.; Pucsok, J.M.; Jámbor, I.; Gyetvai, Á.; Mile, M.; Barna, L.; Szodoray, P.; Tarr, T.; Csiki, Z.; et al. The Effect of Aerobic Exercise and Low-Impact Pilates Workout on the Adaptive Immune System. J. Clin. Med. 2022, 11, 6814. https://doi.org/10.3390/jcm11226814
Balogh L, Szabó K, Pucsok JM, Jámbor I, Gyetvai Á, Mile M, Barna L, Szodoray P, Tarr T, Csiki Z, et al. The Effect of Aerobic Exercise and Low-Impact Pilates Workout on the Adaptive Immune System. Journal of Clinical Medicine. 2022; 11(22):6814. https://doi.org/10.3390/jcm11226814
Chicago/Turabian StyleBalogh, László, Krisztina Szabó, József Márton Pucsok, Ilona Jámbor, Ágnes Gyetvai, Marianna Mile, Lilla Barna, Peter Szodoray, Tünde Tarr, Zoltán Csiki, and et al. 2022. "The Effect of Aerobic Exercise and Low-Impact Pilates Workout on the Adaptive Immune System" Journal of Clinical Medicine 11, no. 22: 6814. https://doi.org/10.3390/jcm11226814
APA StyleBalogh, L., Szabó, K., Pucsok, J. M., Jámbor, I., Gyetvai, Á., Mile, M., Barna, L., Szodoray, P., Tarr, T., Csiki, Z., & Papp, G. (2022). The Effect of Aerobic Exercise and Low-Impact Pilates Workout on the Adaptive Immune System. Journal of Clinical Medicine, 11(22), 6814. https://doi.org/10.3390/jcm11226814