

Mechanosensitive Stem-Cell Genes and Klotho in Atherosclerotic Aortas: Regulating Spatially Deranged Expression Patterns Using Colchicine Regimens

 ,
,  ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
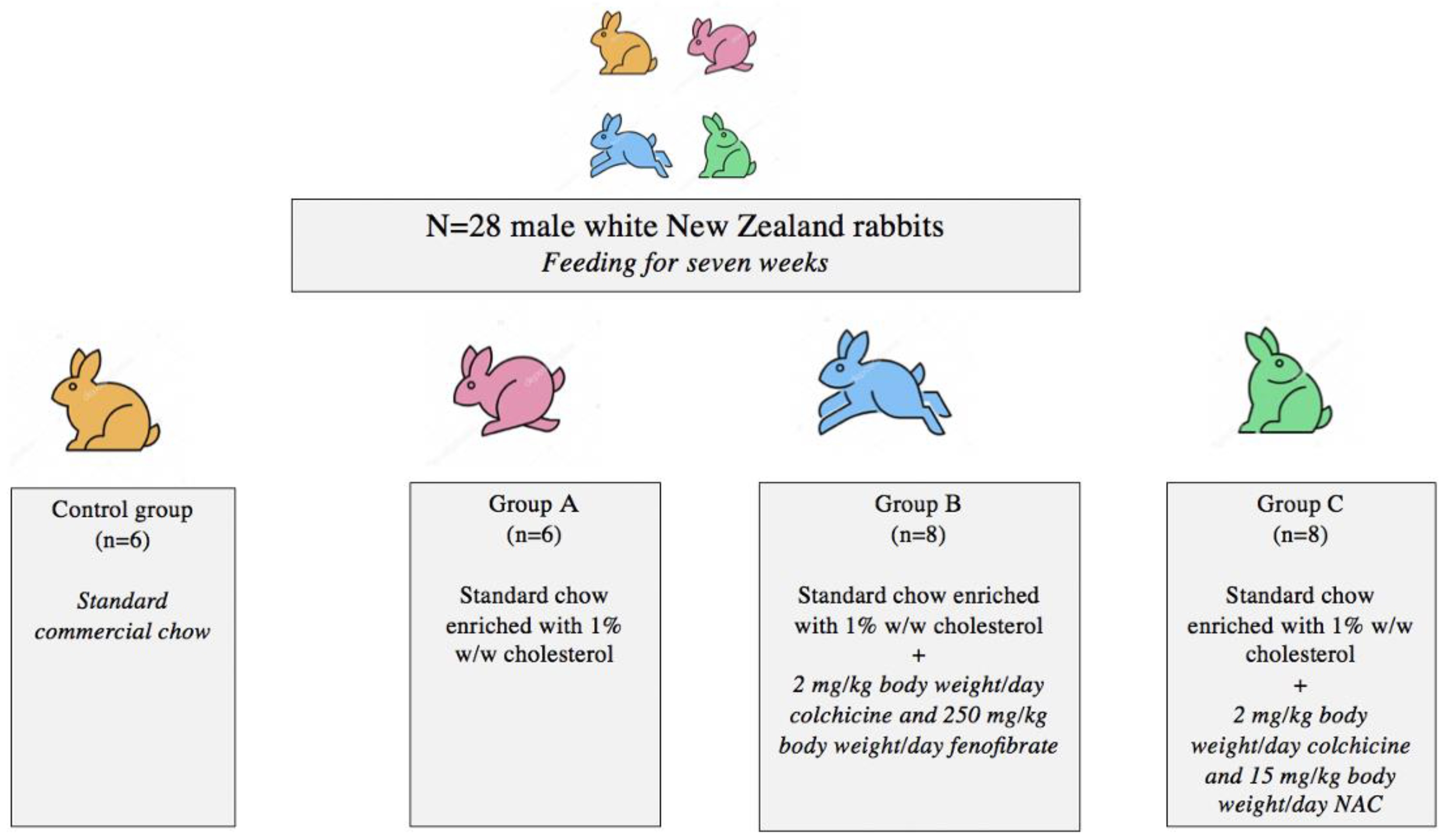
2.1. Animal Model and Experimental Design
2.2. Tissue Preparation
2.3. Histology
2.4. Morphometric Analysis
2.5. Genes of Interest
2.6. RNA Extraction—cDNA Synthesis
2.7. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.8. Statistical Analysis
3. Results
3.1. Atherosclerotic Burden in Thoracic and Abdominal Aortic Specimens
3.2. α-Klotho Expression
3.2.1. α-Klotho in Thoracic Aortic Specimens
3.2.2. α-Klotho in Abdominal Aortic Specimens
3.3. NANOG Expression
3.3.1. NANOG in Thoracic Aortic Specimens
3.3.2. NANOG in Abdominal Aortic Specimens
3.4. NOTCH1 Expression
3.4.1. NOTCH1 in Thoracic Aortic Specimens
3.4.2. NOTCH1 in Abdominal Aortic Specimens
3.5. HIF1a Expression
3.5.1. HIF1a in Thoracic Aortic Specimens
3.5.2. HIF1a in Abdominal Aortic Specimens
3.6. HOXA5 Expression
3.6.1. HOXA5 in Thoracic Aortic Specimens
3.6.2. HOXA5 in Abdominal Aortic Specimens
3.7. BMP4, SOX2, and OCT4 Expression
3.8. Summary of Findings
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Davies, P.F.; Civelek, M.; Fang, Y.; Fleming, I. The atherosusceptible endothelium: Endothelial phenotypes in complex haemodynamic shear stress regions in vivo. Cardiovasc. Res. 2013, 99, 315–327. [Google Scholar] [CrossRef] [PubMed]
- Davies, P.F. Hemodynamic shear stress and the endothelium in cardiovascular pathophysiology. Nat. Clin. Pract. Cardiovasc. Med. 2009, 6, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Libby, P.; Hansson, G.K. Taming Immune and Inflammatory Responses to Treat Atherosclerosis. J. Am. Coll. Cardiol. 2018, 71, 173–176. [Google Scholar] [CrossRef] [PubMed]
- Nidorf, S.M.; Eikelboom, J.W.; Budgeon, C.A.; Thompson, P.L. Low-dose colchicine for secondary prevention of cardiovascular disease. J. Am. Coll. Cardiol. 2013, 61, 404–410. [Google Scholar] [CrossRef]
- Tardif, J.C.; Kouz, S.; Waters, D.D.; Bertrand, O.F.; Diaz, R.; Maggioni, A.P.; Pinto, F.J.; Ibrahim, R.; Gamra, H.; Kiwan, G.S.; et al. Efficacy and Safety of Low-Dose Colchicine after Myocardial Infarction. N. Engl. J. Med. 2019, 381, 2497–2505. [Google Scholar] [CrossRef]
- Ravelli, R.B.; Gigant, B.; Curmi, P.A.; Jourdain, I.; Lachkar, S.; Sobel, A.; Knossow, M. Insight into tubulin regulation from a complex with colchicine and a stathmin-like domain. Nature 2004, 428, 198–202. [Google Scholar] [CrossRef]
- Perico, N.; Ostermann, D.; Bontempeill, M.; Morigi, M.; Amuchastegui, C.S.; Zoja, C.; Akalin, E.; Sayegh, M.H.; Remuzzi, G. Colchicine interferes with L-selectin and leukocyte function-associated antigen-1 expression on human T lymphocytes and inhibits T cell activation. J. Am. Soc. Nephrol. JASN 1996, 7, 594–601. [Google Scholar] [CrossRef]
- Prasad, G.S.; Govardhan, P.; Deepika, G.; Vakdevi, V.; Sashidhar, R.B. Anti-inflammatory activity of anti-hyperlipidemic drug, fenofibrate, and its phase-I metabolite fenofibric acid: In silico, in vitro, and in vivo studies. Inflammopharmacology 2018, 26, 973–981. [Google Scholar] [CrossRef]
- Duez, H.; Chao, Y.S.; Hernandez, M.; Torpier, G.; Poulain, P.; Mundt, S.; Mallat, Z.; Teissier, E.; Burton, C.A.; Tedgui, A.; et al. Reduction of atherosclerosis by the peroxisome proliferator-activated receptor alpha agonist fenofibrate in mice. J. Biol. Chem. 2002, 277, 48051–48057. [Google Scholar] [CrossRef]
- Lee, J.N.; Dutta, R.K.; Kim, S.G.; Lim, J.Y.; Kim, S.J.; Choe, S.K.; Yoo, K.W.; Song, S.R.; Park, D.S.; So, H.S.; et al. Fenofibrate, a peroxisome proliferator-activated receptor α ligand, prevents abnormal liver function induced by a fasting-refeeding process. Biochem. Biophys. Res. Commun. 2013, 442, 22–27. [Google Scholar] [CrossRef]
- Oliveira, A.C.; Bertollo, C.M.; Rocha, L.T.; Nascimento, E.B., Jr.; Costa, K.A.; Coelho, M.M. Antinociceptive and antiedematogenic activities of fenofibrate, an agonist of PPAR alpha, and pioglitazone, an agonist of PPAR gamma. Eur. J. Pharmacol. 2007, 561, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Pieralisi, A.; Martini, C.; Soto, D.; Vila, M.C.; Calvo, J.C.; Guerra, L.N. N-acetylcysteine inhibits lipid accumulation in mouse embryonic adipocytes. Redox Biol. 2016, 9, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.; Spagnoli, V.; Tardif, J.C.; L’Allier, P.L. Novel anti-inflammatory therapies for the treatment of atherosclerosis. Atherosclerosis 2015, 240, 497–509. [Google Scholar] [CrossRef]
- Maekawa, Y.; Ishikawa, K.; Yasuda, O.; Oguro, R.; Hanasaki, H.; Kida, I.; Takemura, Y.; Ohishi, M.; Katsuya, T.; Rakugi, H. Klotho suppresses TNF-alpha-induced expression of adhesion molecules in the endothelium and attenuates NF-kappaB activation. Endocrine 2009, 35, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Mack, J.J.; Mosqueiro, T.S.; Archer, B.J.; Jones, W.M.; Sunshine, H.; Faas, G.C.; Briot, A.; Aragón, R.L.; Su, T.; Romay, M.C.; et al. NOTCH1 is a mechanosensor in adult arteries. Nat. Commun. 2017, 8, 1620. [Google Scholar] [CrossRef]
- An, Z.; Liu, Y.; Song, Z.G.; Tang, H.; Yuan, Y.; Xu, Z.Y. Mechanisms of aortic dissection smooth muscle cell phenotype switch. J. Thorac. Cardiovasc. Surg. 2017, 154, 1511–1521.e1516. [Google Scholar] [CrossRef]
- Jing, Y.; Gao, B.; Han, Z.; Xin, S. HOXA5 induces M2 macrophage polarization to attenuate carotid atherosclerosis by activating MED1. IUBMB Life 2021, 73, 1142–1152. [Google Scholar] [CrossRef]
- Feng, S.; Bowden, N.; Fragiadaki, M.; Souilhol, C.; Hsiao, S.; Mahmoud, M.; Allen, S.; Pirri, D.; Ayllon, B.T.; Akhtar, S.; et al. Mechanical Activation of Hypoxia-Inducible Factor 1α Drives Endothelial Dysfunction at Atheroprone Sites. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 2087–2101. [Google Scholar] [CrossRef]
- Mylonas, K.S.; Karangelis, D.; Androutsopoulou, V.; Chalikias, G.; Tziakas, D.; Mikroulis, D.; Iliopoulos, D.C.; Nikiteas, N.; Schizas, D. Stem cell genes in atheromatosis: The role of Klotho, HIF1α, OCT4, and BMP4. IUBMB Life 2022, 74, 1003–1011. [Google Scholar] [CrossRef]
- Mylonas, K.S.; Kapelouzou, A.; Spartalis, M.; Mastrogeorgiou, M.; Spartalis, E.; Bakoyiannis, C.; Liakakos, T.; Schizas, D.; Iliopoulos, D.; Nikiteas, N. KLF4 Upregulation in Atherosclerotic Thoracic Aortas: Exploring the Protective Effect of Colchicine-based Regimens in a Hyperlipidemic Rabbit Model. Ann. Vasc. Surg. 2022, 78, 328–335. [Google Scholar] [CrossRef]
- Hollander, W.; Paddock, J.; Nagraj, S.; Colombo, M.; Kirkpatrick, B. Effects of anticalcifying and antifibrobrotic drugs on pre-established atherosclerosis in the rabbit. Atherosclerosis 1979, 33, 111–123. [Google Scholar] [CrossRef]
- Kaminiotis, V.V.; Agrogiannis, G.; Konstantopoulos, P.; Androutsopoulou, V.; Korou, L.M.; Vlachos, I.S.; Dontas, I.A.; Perrea, D.; Iliopoulos, D.C. Per os colchicine administration in cholesterol fed rabbits: Triglycerides lowering effects without affecting atherosclerosis progress. Lipids Health Dis. 2017, 16, 184. [Google Scholar] [CrossRef] [PubMed]
- Jeanpierre, E.; Le Tourneau, T.; Zawadzki, C.; Van Belle, E.; Mouquet, F.; Susen, S.; Ezekowitz, M.D.; Staels, B.; Jude, B.; Corseaux, D. Beneficial effects of fenofibrate on plaque thrombogenicity and plaque stability in atherosclerotic rabbits. Cardiovasc. Pathol. Off. J. Soc. Cardiovasc. Pathol. 2009, 18, 140–147. [Google Scholar] [CrossRef]
- Cavallero, C.; Di Tondo, U.; Mingazzini, P.L.; Pesando, P.C.; Spagnoli, L.G. Cell proliferation in the atherosclerotic lesions of cholesterol-fed rabbits. 2. Histological, ultrastructural and radioautographic observations on epinephrine-treated rabbits. Atherosclerosis 1973, 17, 49–62. [Google Scholar] [CrossRef]
- Yanni, A.E.; Agrogiannis, G.; Nomikos, T.; Fragopoulou, E.; Pantopoulou, A.; Antonopoulou, S.; Perrea, D. Oral supplementation with L-aspartate and L-glutamate inhibits atherogenesis and fatty liver disease in cholesterol-fed rabbit. Amino Acids 2010, 38, 1323–1331. [Google Scholar] [CrossRef]
- Kapelouzou, A.; Giaglis, S.; Peroulis, M.; Katsimpoulas, M.; Moustardas, P.; Aravanis, C.V.; Kostakis, A.; Karayannakos, P.E.; Cokkinos, D.V. Overexpression of Toll-Like Receptors 2, 3, 4, and 8 Is Correlated to the Vascular Atherosclerotic Process in the Hyperlipidemic Rabbit Model: The Effect of Statin Treatment. J. Vasc. Res. 2017, 54, 156–169. [Google Scholar] [CrossRef]
- Martín-Núñez, E.; Donate-Correa, J.; López-Castillo, Á.; Delgado-Molinos, A.; Ferri, C.; Rodríguez-Ramos, S.; Cerro, P.; Pérez-Delgado, N.; Castro, V.; Hernández-Carballo, C.; et al. Soluble levels and endogenous vascular gene expression of KLOTHO are related to inflammation in human atherosclerotic disease. Clinical Sci. 2017, 131, 2601–2609. [Google Scholar] [CrossRef]
- Liu, F.; Wu, S.; Ren, H.; Gu, J. Klotho suppresses RIG-I-mediated senescence-associated inflammation. Nat. Cell Biol. 2011, 13, 254–262. [Google Scholar] [CrossRef]
- Wang, Y.; Kuro-o, M.; Sun, Z. Klotho gene delivery suppresses Nox2 expression and attenuates oxidative stress in rat aortic smooth muscle cells via the cAMP-PKA pathway. Aging Cell 2012, 11, 410–417. [Google Scholar] [CrossRef]
- Maltese, G.; Psefteli, P.M.; Rizzo, B.; Srivastava, S.; Gnudi, L.; Mann, G.E.; Siow, R.C. The anti-ageing hormone klotho induces Nrf2-mediated antioxidant defences in human aortic smooth muscle cells. J. Cell. Mol. Med. 2017, 21, 621–627. [Google Scholar] [CrossRef]
- Moe, S.M. Klotho. Circulation 2012, 125, 2181–2183. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.C.; Shi, M.; Zhang, J.; Quiñones, H.; Griffith, C.; Kuro-o, M.; Moe, O.W. Klotho deficiency causes vascular calcification in chronic kidney disease. J. Am. Soc. Nephrol. JASN 2011, 22, 124–136. [Google Scholar] [CrossRef] [PubMed]
- Lim, K.; Lu, T.S.; Molostvov, G.; Lee, C.; Lam, F.T.; Zehnder, D.; Hsiao, L.L. Vascular Klotho deficiency potentiates the development of human artery calcification and mediates resistance to fibroblast growth factor 23. Circulation 2012, 125, 2243–2255. [Google Scholar] [CrossRef] [PubMed]
- Keles, N.; Caliskan, M.; Dogan, B.; Keles, N.N.; Kalcik, M.; Aksu, F.; Kostek, O.; Aung, S.M.; Isbilen, B.; Oguz, A. Low Serum Level of Klotho Is an Early Predictor of Atherosclerosis. Tohoku J. Exp. Med. 2015, 237, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Navarro-González, J.F.; Donate-Correa, J.; Muros de Fuentes, M.; Pérez-Hernández, H.; Martínez-Sanz, R.; Mora-Fernández, C. Reduced Klotho is associated with the presence and severity of coronary artery disease. Heart 2014, 100, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Briot, A.; Civelek, M.; Seki, A.; Hoi, K.; Mack, J.J.; Lee, S.D.; Kim, J.; Hong, C.; Yu, J.; Fishbein, G.A.; et al. Endothelial NOTCH1 is suppressed by circulating lipids and antagonizes inflammation during atherosclerosis. J. Exp. Med. 2015, 212, 2147–2163. [Google Scholar] [CrossRef]
- Noseda, M.; Chang, L.; McLean, G.; Grim, J.E.; Clurman, B.E.; Smith, L.L.; Karsan, A. Notch activation induces endothelial cell cycle arrest and participates in contact inhibition: Role of p21Cip1 repression. Mol. Cell. Biol. 2004, 24, 8813–8822. [Google Scholar] [CrossRef]
- McClain, J.L.; Gulbransen, B.D. The acute inhibition of enteric glial metabolism with fluoroacetate alters calcium signaling, hemichannel function, and the expression of key proteins. J. Neurophysiol. 2017, 117, 365–375. [Google Scholar] [CrossRef]
- Kohler, E.E.; Baruah, J.; Urao, N.; Ushio-Fukai, M.; Fukai, T.; Chatterjee, I.; Wary, K.K. Low-dose 6-bromoindirubin-3’-oxime induces partial dedifferentiation of endothelial cells to promote increased neovascularization. Stem Cells 2014, 32, 1538–1552. [Google Scholar] [CrossRef]
- Jing, Y.; Gao, B.; Han, Z.; Xia, L.; Xin, S. The protective effect of HOXA5 on carotid atherosclerosis occurs by modulating the vascular smooth muscle cell phenotype. Mol. Cell. Endocrinol. 2021, 534, 111366. [Google Scholar] [CrossRef] [PubMed]
- Arderiu, G.; Cuevas, I.; Chen, A.; Carrio, M.; East, L.; Boudreau, N.J. HoxA5 stabilizes adherens junctions via increased Akt1. Cell Adhes. Migr. 2007, 1, 185–195. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Klein, D.; Benchellal, M.; Kleff, V.; Jakob, H.G.; Ergün, S. Hox genes are involved in vascular wall-resident multipotent stem cell differentiation into smooth muscle cells. Sci. Rep. 2013, 3, 2178. [Google Scholar] [CrossRef] [PubMed]
- Biasetti, J.; Spazzini, P.G.; Hedin, U.; Gasser, T.C. Synergy between shear-induced migration and secondary flows on red blood cells transport in arteries: Considerations on oxygen transport. J. R. Soc. Interface 2014, 11, 20140403. [Google Scholar] [CrossRef] [PubMed]
- De Vries, M.R.; Quax, P.H. Plaque angiogenesis and its relation to inflammation and atherosclerotic plaque destabilization. Curr. Opin. Lipidol. 2016, 27, 499–506. [Google Scholar] [CrossRef]
- Akhtar, S.; Hartmann, P.; Karshovska, E.; Rinderknecht, F.A.; Subramanian, P.; Gremse, F.; Grommes, J.; Jacobs, M.; Kiessling, F.; Weber, C.; et al. Endothelial Hypoxia-Inducible Factor-1α Promotes Atherosclerosis and Monocyte Recruitment by Upregulating MicroRNA-19a. Hypertension 2015, 66, 1220–1226. [Google Scholar] [CrossRef]
- Shan, F.; Huang, Z.; Xiong, R.; Huang, Q.Y.; Li, J. HIF1α-induced upregulation of KLF4 promotes migration of human vascular smooth muscle cells under hypoxia. J. Cell. Physiol. 2020, 235, 141–150. [Google Scholar] [CrossRef]
- Aarup, A.; Pedersen, T.X.; Junker, N.; Christoffersen, C.; Bartels, E.D.; Madsen, M.; Nielsen, C.H.; Nielsen, L.B. Hypoxia-Inducible Factor-1α Expression in Macrophages Promotes Development of Atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2016, 36, 1782–1790. [Google Scholar] [CrossRef]
- Yang, P.; Troncone, L.; Augur, Z.M.; Kim, S.S.J.; McNeil, M.E.; Yu, P.B. The role of bone morphogenetic protein signaling in vascular calcification. Bone 2020, 141, 115542. [Google Scholar] [CrossRef]
- Boström, K.I.; Yao, J.; Guihard, P.J.; Blazquez-Medela, A.M.; Yao, Y. Endothelial-mesenchymal transition in atherosclerotic lesion calcification. Atherosclerosis 2016, 253, 124–127. [Google Scholar] [CrossRef]
- Schechtman, L.M. Implementation of the 3Rs (refinement, reduction, and replacement): Validation and regulatory acceptance considerations for alternative toxicological test methods. ILAR J. 2002, 43, S85–S94. [Google Scholar] [CrossRef]
- Bassiouny, H.S.; Zarins, C.K.; Kadowaki, M.H.; Glagov, S. Hemodynamic stress and experimental aortoiliac atherosclerosis. J. Vasc. Surg. 1994, 19, 426–434. [Google Scholar] [CrossRef]
- Padilla, J.; Jenkins, N.T.; Vieira-Potter, V.J.; Laughlin, M.H. Divergent phenotype of rat thoracic and abdominal perivascular adipose tissues. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 304, R543–R552. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | CD | CD+Col+Fib | CD+Col+NAC |
|---|---|---|---|
| BMP4 | ns | ns | ns |
| SOX2 | ns | ns | ns |
| OCT4 | ns | ns | ns |
| NANOG | ns | ns | ns |
| NOTCH1 | ns | ↑ | ns |
| HIF1a | ns | ns | ns |
| HOXA5 | ns | ns | ns |
| α-Klotho | ↓ | ns | ns |
| Gene | CD | CD+Col+Fib | CD+Col+NAC |
|---|---|---|---|
| BMP4 | ns | ns | ns |
| SOX2 | ns | ns | ns |
| OCT4 | ns | ns | ns |
| NANOG | ns | ns | ns |
| NOTCH1 | ns | ns | ns |
| HIF1a | ns | ↓ | ns |
| HOXA5 | ↓ | ↓ | ns |
| α-Klotho | ns | ns | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mylonas, K.S.; Sarantis, P.; Kapelouzou, A.; Karamouzis, M.V.; Kapetanakis, E.I.; Kontzoglou, K.; Iliopoulos, D.C.; Nikiteas, N.; Schizas, D. Mechanosensitive Stem-Cell Genes and Klotho in Atherosclerotic Aortas: Regulating Spatially Deranged Expression Patterns Using Colchicine Regimens. J. Clin. Med. 2022, 11, 6465. https://doi.org/10.3390/jcm11216465
Mylonas KS, Sarantis P, Kapelouzou A, Karamouzis MV, Kapetanakis EI, Kontzoglou K, Iliopoulos DC, Nikiteas N, Schizas D. Mechanosensitive Stem-Cell Genes and Klotho in Atherosclerotic Aortas: Regulating Spatially Deranged Expression Patterns Using Colchicine Regimens. Journal of Clinical Medicine. 2022; 11(21):6465. https://doi.org/10.3390/jcm11216465
Chicago/Turabian StyleMylonas, Konstantinos S., Panagiotis Sarantis, Alkistis Kapelouzou, Michalis V. Karamouzis, Emmanouil I. Kapetanakis, Konstantinos Kontzoglou, Dimitrios C. Iliopoulos, Nikolaos Nikiteas, and Dimitrios Schizas. 2022. "Mechanosensitive Stem-Cell Genes and Klotho in Atherosclerotic Aortas: Regulating Spatially Deranged Expression Patterns Using Colchicine Regimens" Journal of Clinical Medicine 11, no. 21: 6465. https://doi.org/10.3390/jcm11216465
APA StyleMylonas, K. S., Sarantis, P., Kapelouzou, A., Karamouzis, M. V., Kapetanakis, E. I., Kontzoglou, K., Iliopoulos, D. C., Nikiteas, N., & Schizas, D. (2022). Mechanosensitive Stem-Cell Genes and Klotho in Atherosclerotic Aortas: Regulating Spatially Deranged Expression Patterns Using Colchicine Regimens. Journal of Clinical Medicine, 11(21), 6465. https://doi.org/10.3390/jcm11216465