
Hypothermia after Perinatal Asphyxia Does Not Affect Genes Responsible for Amyloid Production in Neonatal Peripheral Lymphocytes

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Setting and Design
- Newborns (full-term and preterm) > 31 weeks of gestational age.
- Metabolic acidosis with pH < 7.0 (in umbilical cord or newborn blood sample obtained during 60 min after birth),
- or Base deficit > −12,
- or Apgar score of 0–5 at 10 min or continued need for resuscitation at 10 min of age.
- Presence of multiple organ-system failures,
- Clinical evidence of encephalopathy: periodic breathing/apnea, abnormal oculomotor or pupillary movements, weak or absent suck, or clinical seizures.
- Neurologic findings cannot be attributed to other cause.
2.2. Therapeutic Protocol for Hypothermia
- (1)
- Inclusion criteria for perinatal asphyxia treated by hypothermia:
- ▪
- Newborns born after ≥35 weeks of gestation with severe or moderate perinatal asphyxia,
- ▪
- or Apgar ≤ 5 point at 10th minute of life,
- ▪
- or mechanical ventilation 10 min after birth,
- ▪
- or acidosis pH < 7.0 in umbilical or arterial blood 1 h after birth,
- ▪
- or base deficit of at least −16 mmol/L in umbilical or arterial blood 1 h after birth.
- ▪
- or Neurological deficits: coma, lethargy, decreased muscle tone, abnormal response to stimuli, no or reduced sucking reflex, convulsions, increased muscle tone.
- ▪
- Changes in amplitude—integrated EEG recordings.
- (2)
- Exclusion criteria:
- ▪
- Gestational age < 35 weeks.
- ▪
- Birth weight < 1800 g.
- ▪
- Severe birth defects with poor prognosis.
- ▪
- Severe traumatic head injury.
- ▪
- Rectal obstruction which prevents deep body temperature measurements.
- ▪
- Lack of consent of parents/legal guardians.
- ▪
- Over 6 h of life on admission to the intensive care unit.
2.3. Study Population and Sample Size
2.4. Study of Gene Expression, Including Amyloid Protein Precursor, β-Secretase, and Presenilin 1 and 2
3. Results
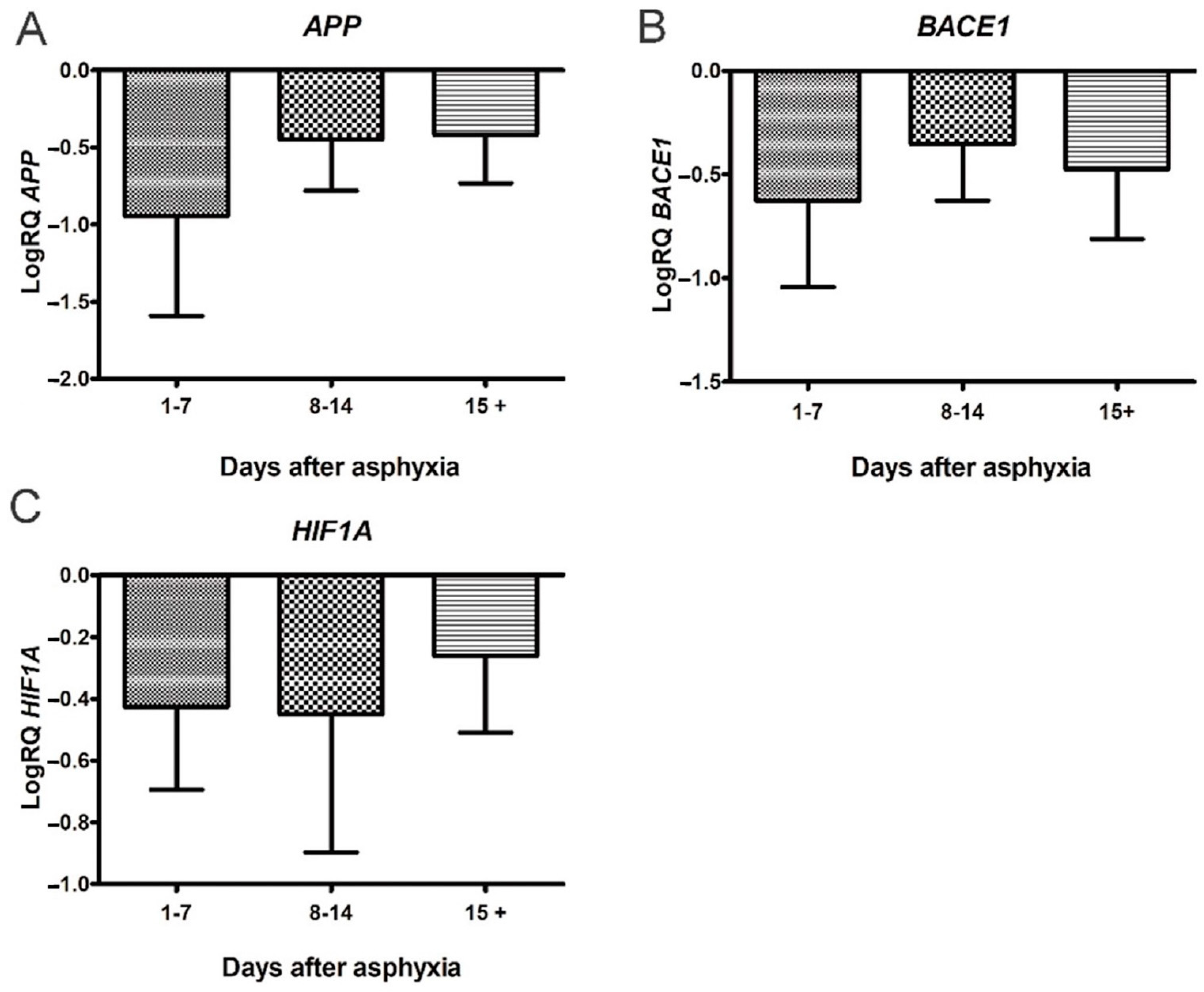
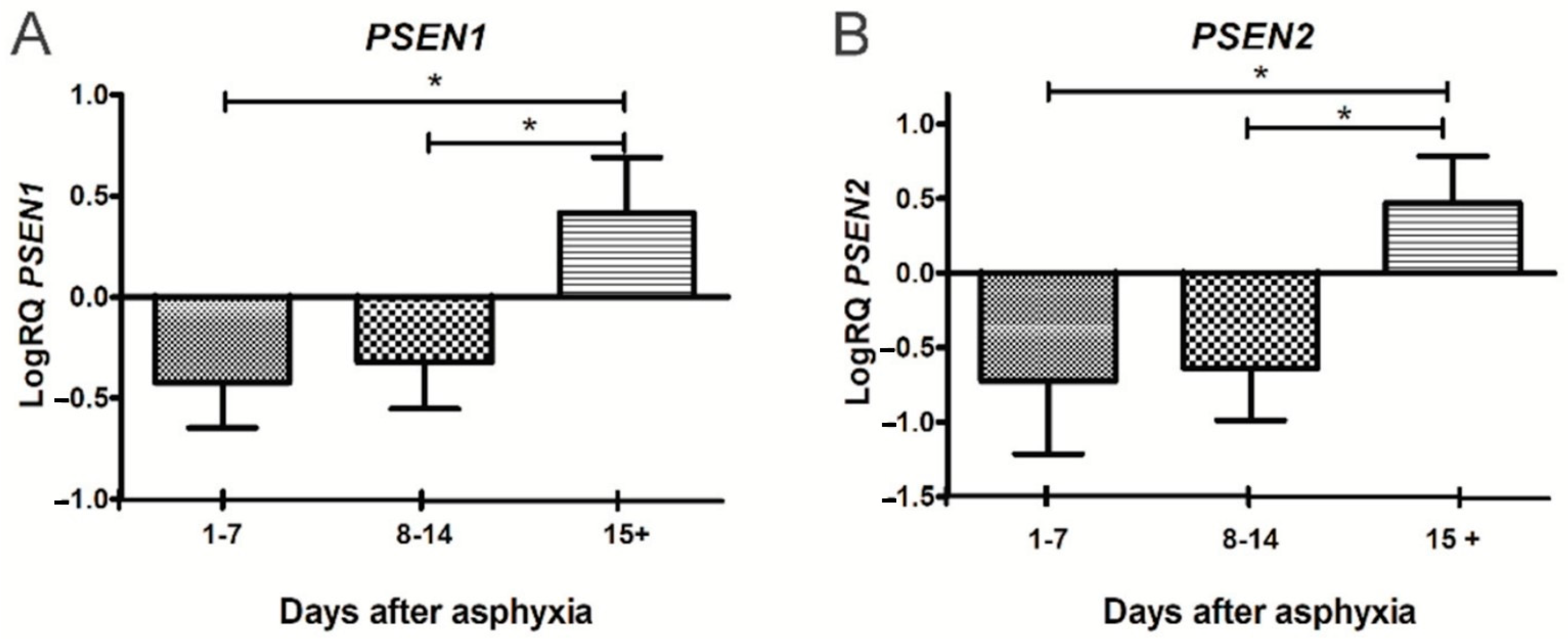
3.1. Expression of Genes Metabolizing Amyloid Protein Precursor in Lymphocytes after Asphyxia
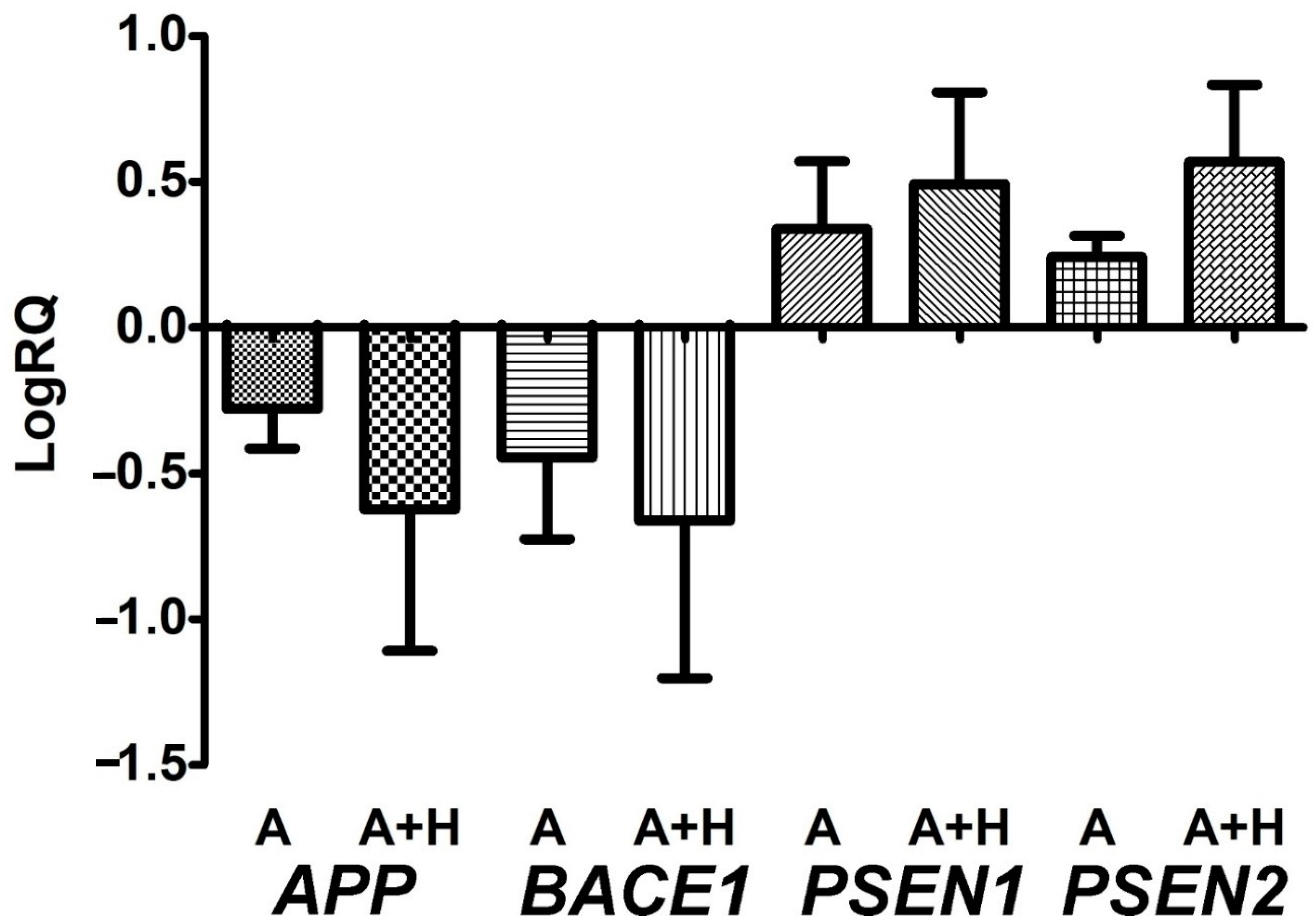
3.2. Expression of Genes Metabolizing Amyloid Protein Precursor in Lymphocytes after Asphyxia Treated with Hypothermia
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Boldingh, A.M.; Solevåg, A.L.; Nakstad, B. Outcomes following neonatal cardiopulmonary resuscitation. Tidsskr. Den Nor. Legeforening 2018, 138. [Google Scholar] [CrossRef]
- Dzikienė, R.; Lukoševičius, S.; Laurynaitienė, J.; Marmienė, V.; Nedzelskienė, I.; Tamelienė, R.; Rimdeikienė, I.; Kudrevičienė, A. Long-term outcomes of perinatal hypoxia and asphyxia at an early school age. Medicina 2021, 57, 988. [Google Scholar] [CrossRef] [PubMed]
- Tarkowska, A.; Furmaga-Jabłońska, W.; Bogucki, J.; Kocki, J.; Pluta, R. Alzheimer’s Disease Associated Presenilin 1 and 2 Genes Dysregulation in Neonatal Lymphocytes Following Perinatal Asphyxia. Int. J. Mol. Sci. 2021, 22, 5140. [Google Scholar] [CrossRef] [PubMed]
- Ahearne, C.E.; Boylan, G.; Murray, D.M. Short and long term prognosis in perinatal asphyxia: An update. World J. Clin. Pediatr. 2016, 5, 67–74. [Google Scholar] [CrossRef]
- Balada, R.; Tebé, C.; León, M.; Arca, G.; Alsina, M.; Castells, A.A.; Alcántara, S.; Garcia-Alix, A. Enquiring beneath the surface: Can a gene expression assay shed light into the heterogeneity among newborns with neonatal encephalopathy? Pediatr. Res. 2020, 88, 451–458. [Google Scholar] [CrossRef]
- Shankaran, S.; Laptook, A.R.; Pappas, A.; McDonald, S.A.; Das, A.; Tyson, J.E.; Poindexter, B.B.; Schibler, K.; Bell, E.F.; Heyne, R.J.; et al. Eunice Kennedy Shriver National Institute of Child Health and Human Development Neonatal Research Network. Effect of depth and duration of cooling on death or disability at age 18 months among neonates with hypoxic-ischemic encephalopathy: A randomized clinical trial. JAMA 2017, 318, 57–67. [Google Scholar]
- Chin, E.M.; Jayakumar, S.; Ramos, E.; Gerner, G.; Soares, B.P.; Cristofalo, E.; Leppert, M.; Allen, M.; Parkinson, C.; Johnston, M.; et al. Preschool Language Outcomes following Perinatal Hypoxic-Ischemic Encephalopathy in the Age of Therapeutic Hypothermia. Dev. Neurosci. 2018, 40, 627–637. [Google Scholar] [CrossRef]
- Edmonds, C.J.; Helps, S.K.; Hart, D.; Zatorska, A.; Gupta, N.; Cianfaglione, R.; Vollmer, B. Minor neurological signs and behavioural function at age 2 years in neonatal hypoxic ischemic encephalopathy (HIE). Eur. J. Paediatr. Neurol. 2020, 27, 78–85. [Google Scholar] [CrossRef]
- Grass, B.; Scheidegger, S.; Latal, B.; Hagmann, C.; Held, U.; Brotschi, B. Short-term neurological improvement in neonates with hypoxic-ischemic encephalopathy predicts neurodevelopmental outcome at 18–24 months. J. Périnat. Med. 2020, 48, 296–303. [Google Scholar] [CrossRef]
- Monzani, A.; Cosi, G.; Genoni, G.; Lavrano, M.; Peruzzi, C.; Pomella, R.; Ferrero, F. Early instrumental predictors of long-term neurodevelopmental impairment in newborns with perinatal asphyxia treated with therapeutic hypothermia. Signa Vitae 2018, 14, 81–85. [Google Scholar]
- Reiss, J.; Sinha, M.; Gold, J.; Bykowski, J.; Lawrence, S.M. Outcomes of Infants with Mild Hypoxic Ischemic Encephalopathy Who Did Not Receive Therapeutic Hypothermia. Biomed. Hub 2019, 4, 502936. [Google Scholar] [CrossRef] [PubMed]
- Schreglmann, M.; Ground, A.; Vollmer, B.; Johnson, M.J. Systematic review: Long-term cognitive and behavioural outcomes of neonatal hypoxic–ischaemic encephalopathy in children without cerebral palsy. Acta Paediatr. 2019, 109, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Li, L.; Zhang, X.; Xie, W.; Li, L.; Yang, D.; Heng, X.; Du, Y.; Doody, R.S.; Le, W. Prenatal hypoxia may aggravate the cognitive impairment and Alzheimer’s disease neuropathology in APPSwe/PS1A246E transgenic mice. Neurobiol. Aging 2013, 34, 663–678. [Google Scholar] [CrossRef] [PubMed]
- Chavez-Valdez, R.; Lechner, C.; Emerson, P.; Northington, F.J.; Martin, L.J. Accumulation of PSA-NCAM marks nascent neurodegeneration in the dorsal hippocampus after neonatal hypoxic-ischemic brain injury in mice. J. Cereb. Blood Flow Metab. 2020, 41, 1039–1057. [Google Scholar] [CrossRef] [PubMed]
- Tarkowska, A. Hypoxic-ischemic brain injury after perinatal asphyxia as a possible factor in the pathology of Alzheimer’s disease. In Cerebral Ischemia; Pluta, R., Ed.; Exon Publications: Brisbane, Australia, 2021; pp. 45–59. [Google Scholar]
- Schiefecker, A.J.; Putzer, G.; Braun, P.; Martini, J.; Strapazzon, G.; Antunes, A.P.; Mulino, M.; Pinggera, D.; Glodny, B.; Brugger, H.; et al. Total TauProtein as Investigated by Cerebral Microdialysis Increases in Hypothermic Cardiac Arrest: A Pig Study. Ther. Hypothermia Temp. Manag. 2021, 11, 28–34. [Google Scholar] [CrossRef]
- Takahashi, K.; Hasegawa, S.; Maeba, S.; Fukunaga, S.; Motoyama, M.; Hamano, H.; Ichiyama, T. Serum tau protein level serves as a predictive factor for neurological prognosis in neonatal asphyxia. Brain Dev. 2014, 36, 670–675. [Google Scholar] [CrossRef]
- Benterud, T.; Pankratov, L.; Solberg, R.; Bolstad, N.; Skinningsrud, A.; Baumbusch, L.; Sandvik, L.; Saugstad, O.D. Perinatal Asphyxia May Influence the Level of Beta-Amyloid (1-42) in Cerebrospinal Fluid: An Experimental Study on Newborn Pigs. PLoS ONE 2015, 10, e0140966. [Google Scholar] [CrossRef]
- Karran, E.; Mercken, M.; De Strooper, B. The amyloid cascade hypothesis for Alzheimer’s disease: An appraisal for the development of therapeutics. Nat. Rev. Drug Discov. 2011, 10, 698–712. [Google Scholar] [CrossRef]
- Hansson, O.; Zetterberg, H.; Vanmechelen, E.; Vanderstichele, H.; Andreasson, U.; Londos, E.; Wallin, A.; Minthon, L.; Blennow, K. Evaluation of plasma Abeta(40) and Abeta(42) as predictors of conversion to Alzheimer’s disease in patients with mild cognitive impairment. Neurobiol. Aging 2010, 31, 357–367. [Google Scholar] [CrossRef]
- Jacobs, S.E.; Berg, M.; Hunt, R.; Tarnow-Mordi, W.O.; Inder, T.E.; Davis, P.G. Cooling for newborns with hypoxic ischaemic encephalopathy. Cochrane Database Syst. Rev. 2013, 1, CD003311. [Google Scholar] [CrossRef]
- Martinello, K.; Hart, A.R.; Yap, S.; Mitra, S.; Robertson, N.J. Management and investigation of neonatal encephalopathy: 2017 update. Arch. Dis. Child.-Fetal Neonatal Ed. 2017, 102, F346–F358. [Google Scholar] [CrossRef] [PubMed]
- Michniewicz, B.; Szpecht, D.; Sowińska, A.; Sibiak, R.; Szymankiewicz, M.; Gadzinowski, J. Biomarkers in newborns with hypoxic-ischemic encephalopathy treated with therapeutic hypothermia. Child’s Nerv. Syst. 2020, 36, 2981–2988. [Google Scholar] [CrossRef] [PubMed]
- Gano, D.; Orbach, S.A.; Bonifacio, S.L.; Glass, H.C. Neonatal seizures and therapeutic hypothermia for hypoxic-ischemic encephalopathy. Mol. Cell. Epilepsy 2014, 1, e88. [Google Scholar] [CrossRef]
- Natarajan, G.; Pappas, A.; Shankaran, S. Outcomes in childhood following therapeutic hypothermia for neonatal hypoxic-ischemic encephalopathy (HIE). Semin. Perinatol. 2016, 40, 549–555. [Google Scholar] [CrossRef] [Green Version]
- Hakobyan, M.; Dijkman, K.P.; Laroche, S.; Naulaers, G.; Rijken, M.; Steiner, K.; Van Straaten, H.L.; Swarte, R.M.; Ter Horst, H.J.; Zecic, A.; et al. Outcome of Infants with Therapeutic Hypothermia after Perinatal Asphyxia and Early-Onset Sepsis. Neonatology 2018, 115, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Pappas, A.; Shankaran, S.; McDonald, S.; Vohr, B.R.; Hintz, S.R.; Ehrenkranz, R.A.; Tyson, J.E.; Yolton, K.; Das, A.; Bara, R.; et al. Hypothermia Extended Follow-up Subcommittee of the Eunice Kennedy Shriver NICHD Neonatal Research Network. Cognitive outcomes after neonatal encephalopathy. Pediatrics 2015, 135, e624–e634. [Google Scholar] [CrossRef] [Green Version]
- Lee-Kelland, R.; Jary, S.; Tonks, J.; Cowan, F.M.; Thoresen, M.; Chakkarapani, E. School-age outcomes of children without cerebral palsy cooled for neonatal hypoxic–ischaemic encephalopathy in 2008–2010. Arch. Dis. Child.-Fetal Neonatal Ed. 2019, 105, 8–13. [Google Scholar] [CrossRef] [Green Version]
- Chavez-Valdez, R.; Emerson, P.; Goffigan-Holmes, J.; Kirkwood, A.; Martin, L.J.; Northington, F.J. Delayed injury of hippocampal interneurons after neonatal hypoxia-ischemia and therapeutic hypothermia in a murine model. Hippocampus 2018, 28, 617–630. [Google Scholar] [CrossRef]
- Goffigan-Holmes, J.; Sanabria, D.; Diaz, J.; Flock, D. Chavez-Valdez R: Calbindin-1 expression in the hippocampus following neonatal hypoxia-ischemia and therapeutic hypothermia and deficits in spatial memory. Dev. Neurosci. 2018, 40, 508–522. [Google Scholar] [CrossRef]
- Davidson, J.O.; Wassink, G.; van den Heuij, L.G.; Bennet, L.; Gunn, A.J. Therapeutic hypothermia for neonatal hypoxic-ischemic encephalopathy-where to from here? Front. Neurol. 2015, 6, 198. [Google Scholar] [CrossRef] [Green Version]
- Pluta, R.; Kocki, J.; Ułamek-Kozioł, M.; Bogucka-Kocka, A.; Gil-Kulik, P.; Januszewski, S.; Jabłoński, M.; Petniak, A.; Brzozowska, J.; Bogucki, J.; et al. Alzheimer-associated presenilin 2 gene is dysregulated in rat medial temporal lobe cortex after complete brain ischemia due to cardiac arrest. Pharmacol. Rep. 2016, 68, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Pluta, R.; Bogucka-Kocka, A.; Ułamek-Kozioł, M.; Bogucki, J.; Januszewski, S.; Kocki, J.; Czuczwar, S.J. Ischemic tau protein gene induction as an additional key factor driving development of Alzheimer’s phenotype changes in CA1 area of hippocampus in an ischemic model of Alzheimer’s disease. Pharmacol. Rep. 2018, 70, 881–884. [Google Scholar] [CrossRef] [PubMed]
- Robbins, J.R.; Lee, S.M.; Filipovich, A.H.; Szigligeti, P.; Neumeier, L.; Petrovic, M.; Conforti, L. Hypoxia modulates early events in T cell receptor-mediated activation in human T lymphocytes via Kv1.3 channels. J. Physiol. 2005, 564, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Chimote, A.A.; Kuras, Z.; Conforti, L. Disruption of kv1.3 channel forward vesicular trafficking by hypoxia in human T lymphocytes. J. Biol. Chem. 2012, 287, 2055–2067. [Google Scholar]
- Feng, Y.; Liao, S.; Wei, C.; Jia, D.; Wood, K.; Liu, Q.; Wang, X.; Shi, F.-D.; Jin, W.-N. Infiltration and persistence of lymphocytes during late-stage cerebral ischemia in middle cerebral artery occlusion and photothrombotic stroke models. J. Neuroinflamm. 2017, 14, 248. [Google Scholar] [CrossRef] [Green Version]
- Nazmi, A.; Albertsson, A.-M.; Rocha-Ferreira, E.; Zhang, X.; Vontell, R.; Zelco, A.; Rutherford, M.; Zhu, C.; Nilsson, G.; Mallard, C.; et al. Lymphocytes Contribute to the Pathophysiology of Neonatal Brain Injury. Front. Neurol. 2018, 9, 159. [Google Scholar] [CrossRef]
- Pluta, R.; Lossinsky, A.S.; Walski, M.; Wisniewski, H.M.; Mossakowski, M.J. Platelet occlusion phenomenon after short- and long-term survival following complete cerebral ischemia in rats produced by cardiac arrest. J. Hirnforsch. 1994, 35, 463–471. [Google Scholar]
- Pluta, R.; Lossinsky, A.; Wiśniewski, H.; Mossakowski, M. Early blood-brain barrier changes in the rat following transient complete cerebral ischemia induced by cardiac arrest. Brain Res. 1994, 633, 41–52. [Google Scholar] [CrossRef]
- Kucheryavykh, L.Y.; Dávila-Rodríguez, J.; Rivera-Aponte, D.E.; Zueva, L.V.; Washington, A.V.; Sanabria, P.; Inyushin, M.Y. Platelets are responsible for the accumulation of β-amyloid in blood clots inside and around blood vessels in mouse brain after thrombosis. Brain Res. Bull. 2016, 128, 98–105. [Google Scholar] [CrossRef]
- Inyushin, M.; Zayas-Santiago, A.; Rojas, L.; Kucheryavykh, L. On the Role of Platelet-Generated Amyloid Beta Peptides in Certain Amyloidosis Health Complications. Front. Immunol. 2020, 11, 571083. [Google Scholar] [CrossRef]
- Rosenberg, R.N.; Baskin, F.; Fosmire, J.A.; Risser, R.; Adams, P.; Svetlik, D.; Honig, L.S.; Cullum, C.M.; Weiner, M.F. Altered Amyloid Protein Processing in Platelets of Patients with Alzheimer Disease. Arch. Neurol. 1997, 54, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Nataf, S.; Guillen, M.; Pays, L. Common Neurodegeneration-Associated Proteins Are Physiologically Expressed by Human B Lymphocytes and Are Interconnected via the Inflammation/Autophagy-Related Proteins TRAF6 and SQSTM1. Front. Immunol. 2019, 10, 2704. [Google Scholar] [CrossRef] [PubMed]
- Park, M.H.; Choi, D.Y.; Jin, H.W.; Yoo, H.S.; Han, J.-Y.; Oh, K.-W.; Han, S.B.; Hwang, D.Y.; Hong, J.T. Mutant Presenilin 2 Increases β-Secretase Activity Through Reactive Oxygen Species–Dependent Activation of Extracellular Signal–Regulated Kinase. J. Neuropathol. Exp. Neurol. 2012, 71, 130–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galla, L.; Redolfi, N.; Pozzan, T.; Pizzo, P.; Greotti, E. Intracellular Calcium Dysregulation by the Alzheimer’s Disease-Linked Protein Presenilin 2. Int. J. Mol. Sci. 2020, 21, 770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delabio, R.; Rasmussen, L.; Mizumoto, I.; Viani, G.-A.; Chen, E.; Villares, J.; Costa, I.-B.; Turecki, G.; Linde, S.A.; Smith, M.C.; et al. PSEN1 and PSEN2 Gene Expression in Alzheimer’s Disease Brain: A New Approach. J. Alzheimer’s Dis. 2014, 42, 757–760. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, M.; Cai, F.; Song, W. Biological function of Presenilin and its role in AD pathogenesis. Transl. Neurodegener. 2013, 2, 15. [Google Scholar] [CrossRef] [Green Version]
- Qi, J.-P.; Wu, H.; Yang, Y.; Wang, D.-D.; Chen, Y.-X.; Gu, Y.-H.; Liu, T. Cerebral Ischemia and Alzheimer’s Disease: The Expression of Amyloid-β and Apolipoprotein E in Human Hippocampus. J. Alzheimer’s Dis. 2007, 12, 335–341. [Google Scholar] [CrossRef]
- Yasuno, F.; Kajimoto, K.; Ihara, M.; Taguchi, A.; Yamamoto, A.; Fukuda, T.; Kazui, H.; Iida, H.; Nagatsuka, K. Amyloid β deposition in subcortical stroke patients and effects of educational achievement: A pilot study. Int. J. Geriatr. Psychiatry 2019, 34, 1651–1657. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Group | Age (Days) | Gestationl Age (Weeks) | Birth Weight (g) | Apgar Score (1 min) | RBC (×1000/μL) | WBC (/μL) | Lymphocyte (/μL) | PLT (×1000/μL) | Hct (%) | pH | BE (Mmol/L) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | 20 ± 1 | 37 ± 2 | 3274 ± 542 | 10 ± 0 | 4204 ± 416 | 11,816 ± 1692 | 6256 ± 460 | 539 ± 59 | 39 ± 2 | 7.41 ± 0.01 | 0.2 ± 0.4 |
| Asphyxia | 20 ± 1 | 36 ± 3 | 2746 ± 930 | 2 ± 2 | 3598 ± 949 | 14,844 ± 10,406 | 6422 ± 1354 | 282 ± 106 | 36 ± 11 | 7.23 ± 0.28 | −7.0 ± 4.6 |
| Control | 19 ± 4 | 39 ± 1 | 3492 ± 592 | 9 ± 1 | 4526 ± 838 | 13,596 ± 2400 | 8106 ± 1111 | 464 ± 129 | 41 ± 7 | 7.43 ± 0.04 | 0.8 ± 1.7 |
| Asphyxia + Hypothermia | 19 ± 3 | 39 ± 1 | 3128 ± 151 | 2 ± 1 | 4756 ± 266 | 16,658 ± 7151 | 5766 ± 1625 | 220 ± 39 | 47 ± 4 | 7.09 ± 0.25 | −11 ± 4.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tarkowska, A.; Furmaga-Jabłońska, W.; Bogucki, J.; Kocki, J.; Pluta, R. Hypothermia after Perinatal Asphyxia Does Not Affect Genes Responsible for Amyloid Production in Neonatal Peripheral Lymphocytes. J. Clin. Med. 2022, 11, 3263. https://doi.org/10.3390/jcm11123263
Tarkowska A, Furmaga-Jabłońska W, Bogucki J, Kocki J, Pluta R. Hypothermia after Perinatal Asphyxia Does Not Affect Genes Responsible for Amyloid Production in Neonatal Peripheral Lymphocytes. Journal of Clinical Medicine. 2022; 11(12):3263. https://doi.org/10.3390/jcm11123263
Chicago/Turabian StyleTarkowska, Agata, Wanda Furmaga-Jabłońska, Jacek Bogucki, Janusz Kocki, and Ryszard Pluta. 2022. "Hypothermia after Perinatal Asphyxia Does Not Affect Genes Responsible for Amyloid Production in Neonatal Peripheral Lymphocytes" Journal of Clinical Medicine 11, no. 12: 3263. https://doi.org/10.3390/jcm11123263
APA StyleTarkowska, A., Furmaga-Jabłońska, W., Bogucki, J., Kocki, J., & Pluta, R. (2022). Hypothermia after Perinatal Asphyxia Does Not Affect Genes Responsible for Amyloid Production in Neonatal Peripheral Lymphocytes. Journal of Clinical Medicine, 11(12), 3263. https://doi.org/10.3390/jcm11123263