
A Comprehensive Investigation into the Distribution of Circulating B Cell Subsets in the Third Trimester of Pregnancy






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
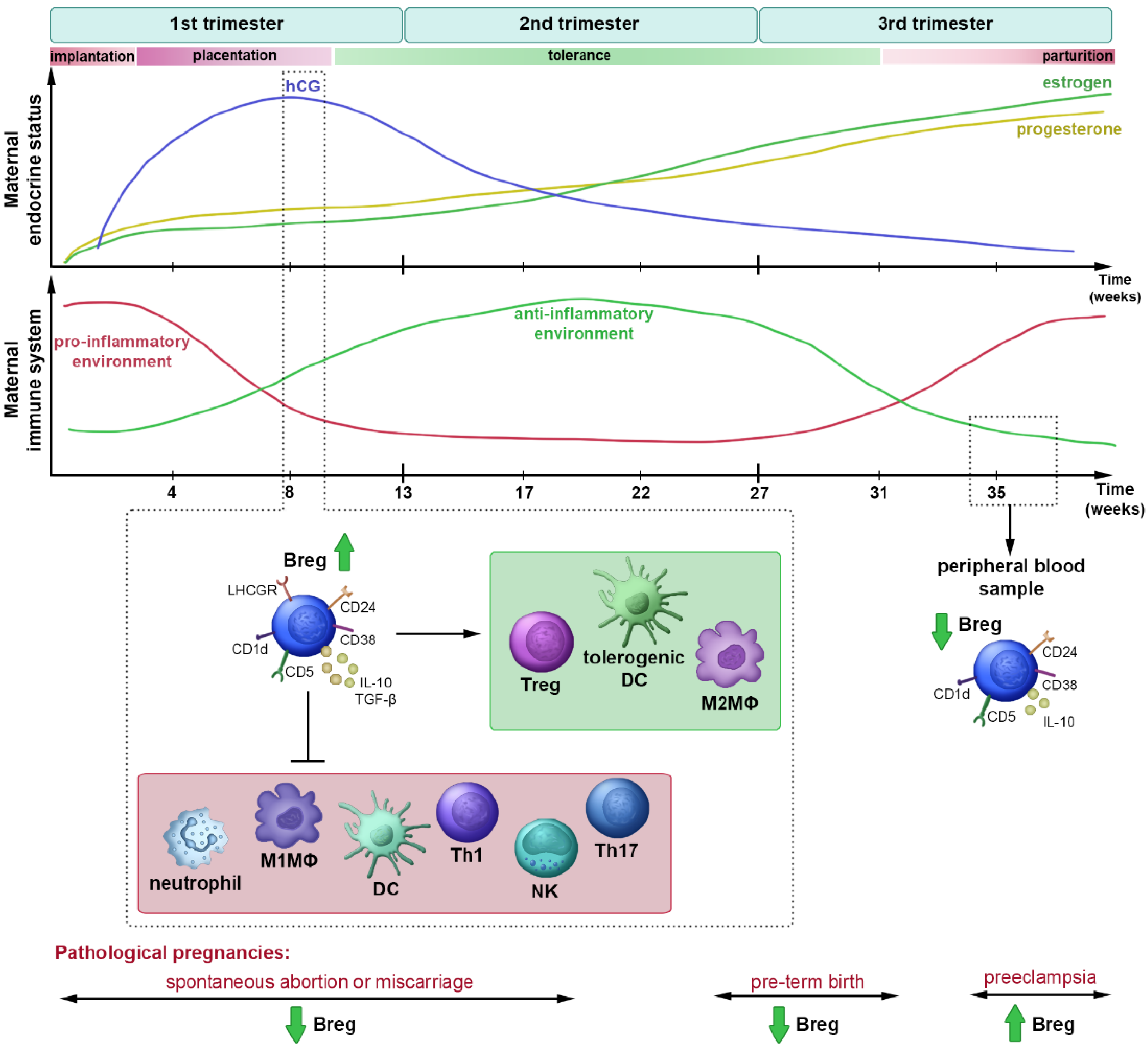
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Flow Cytometric Analysis of Different B Cell Subsets
2.3. Intracellular Cytokine Analysis by Flow Cytometry
2.4. Statistical Analysis
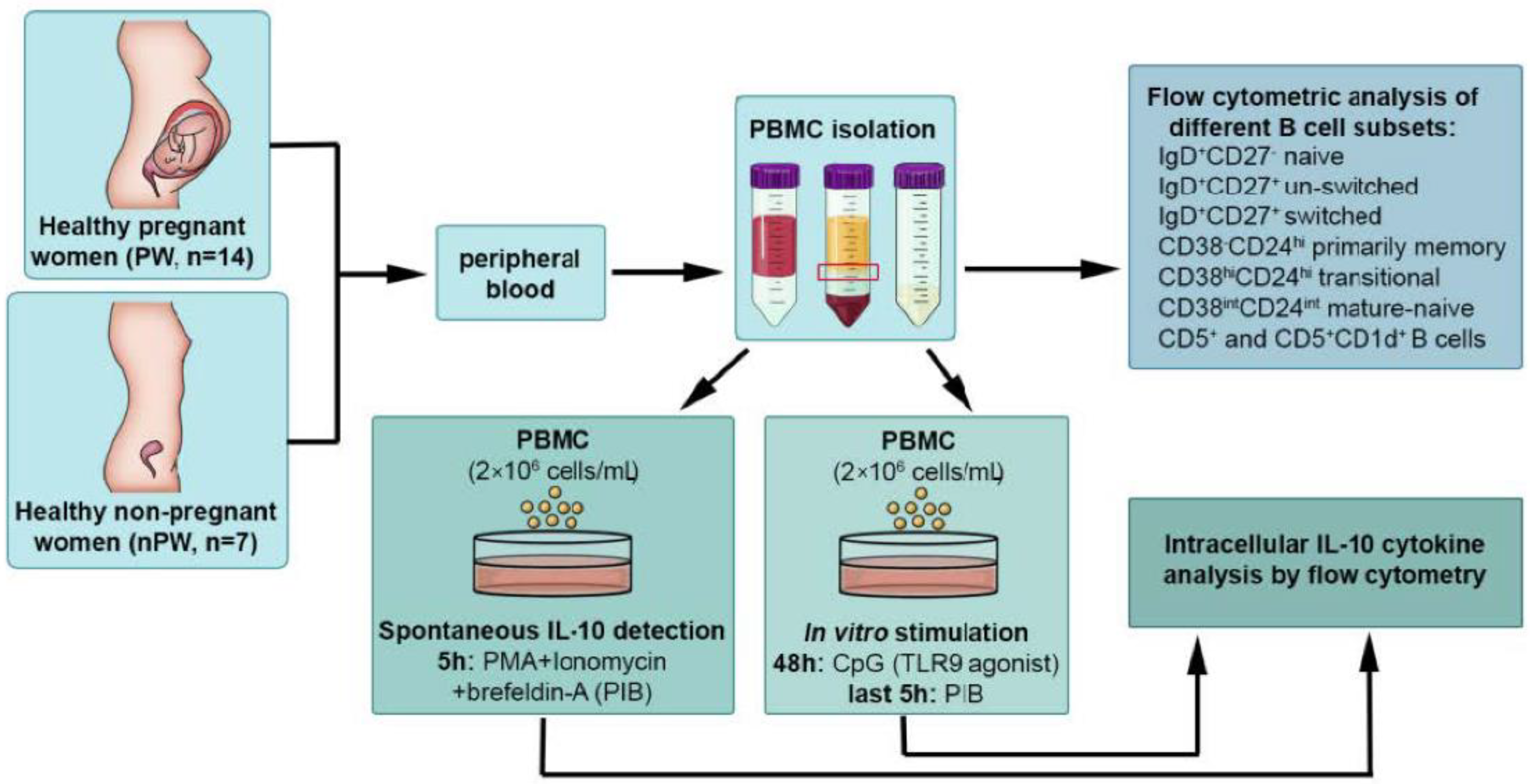
3. Results
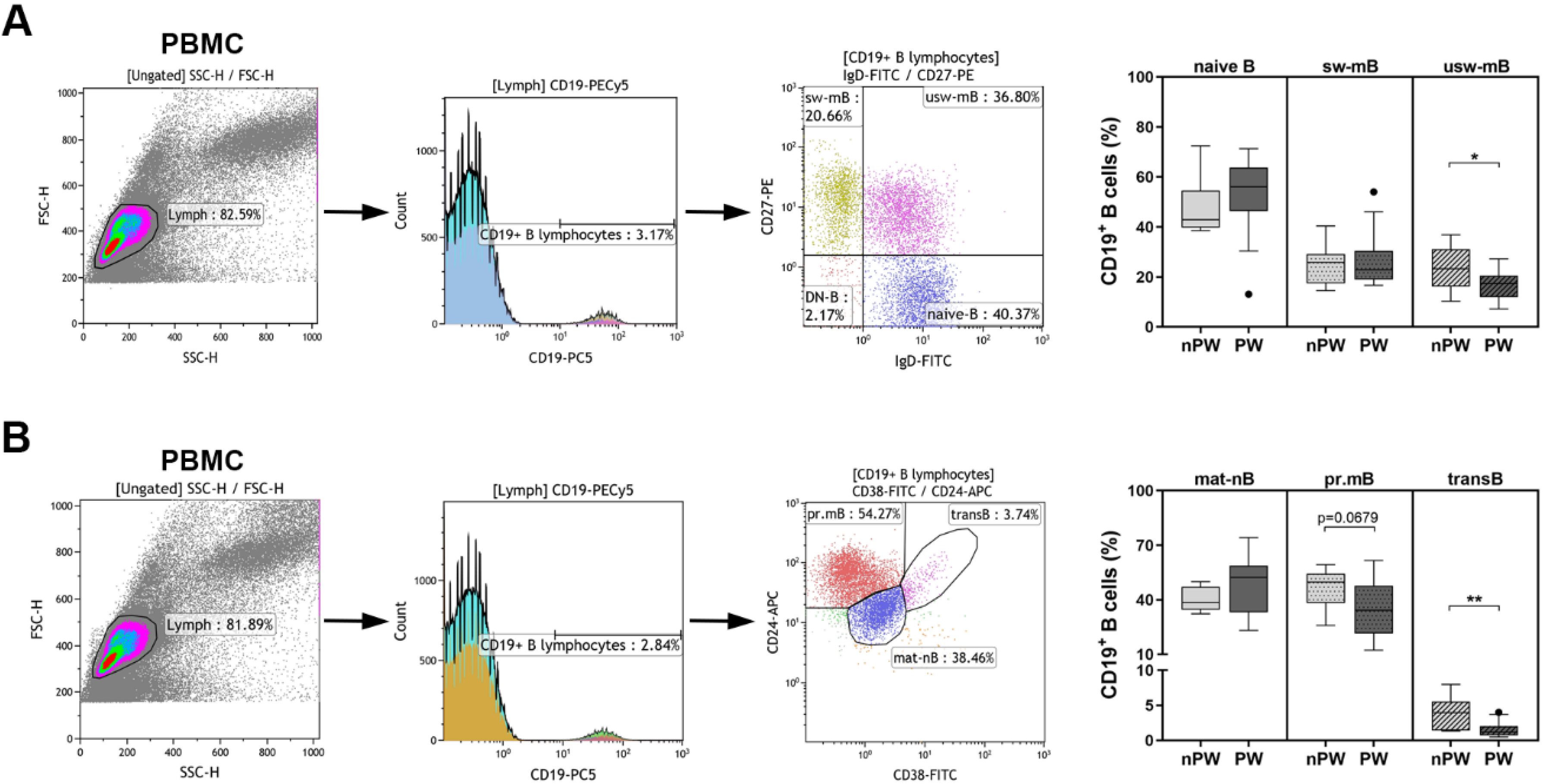
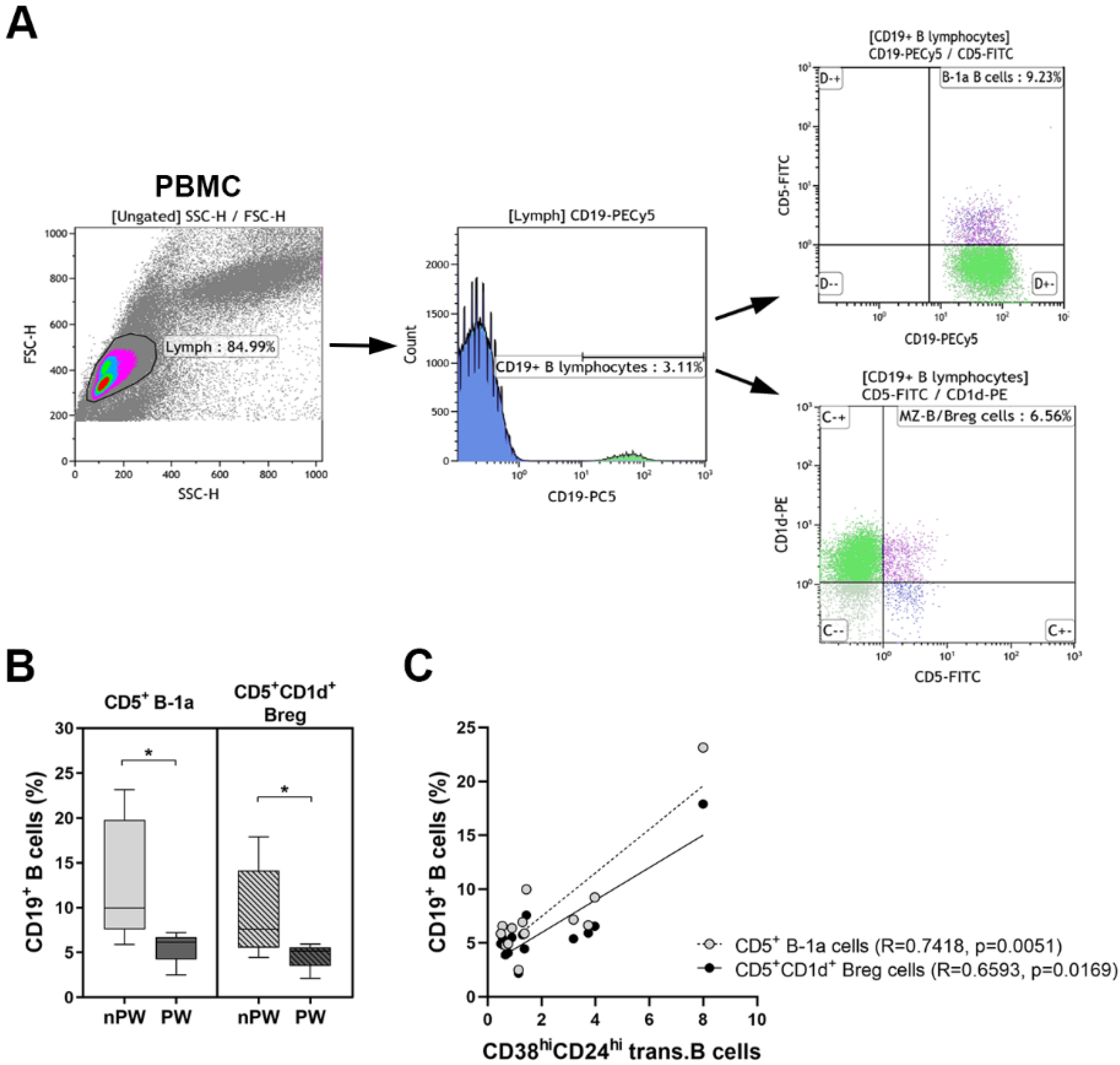
3.1. The Distribution of Peripheral B Cell Subpopulations
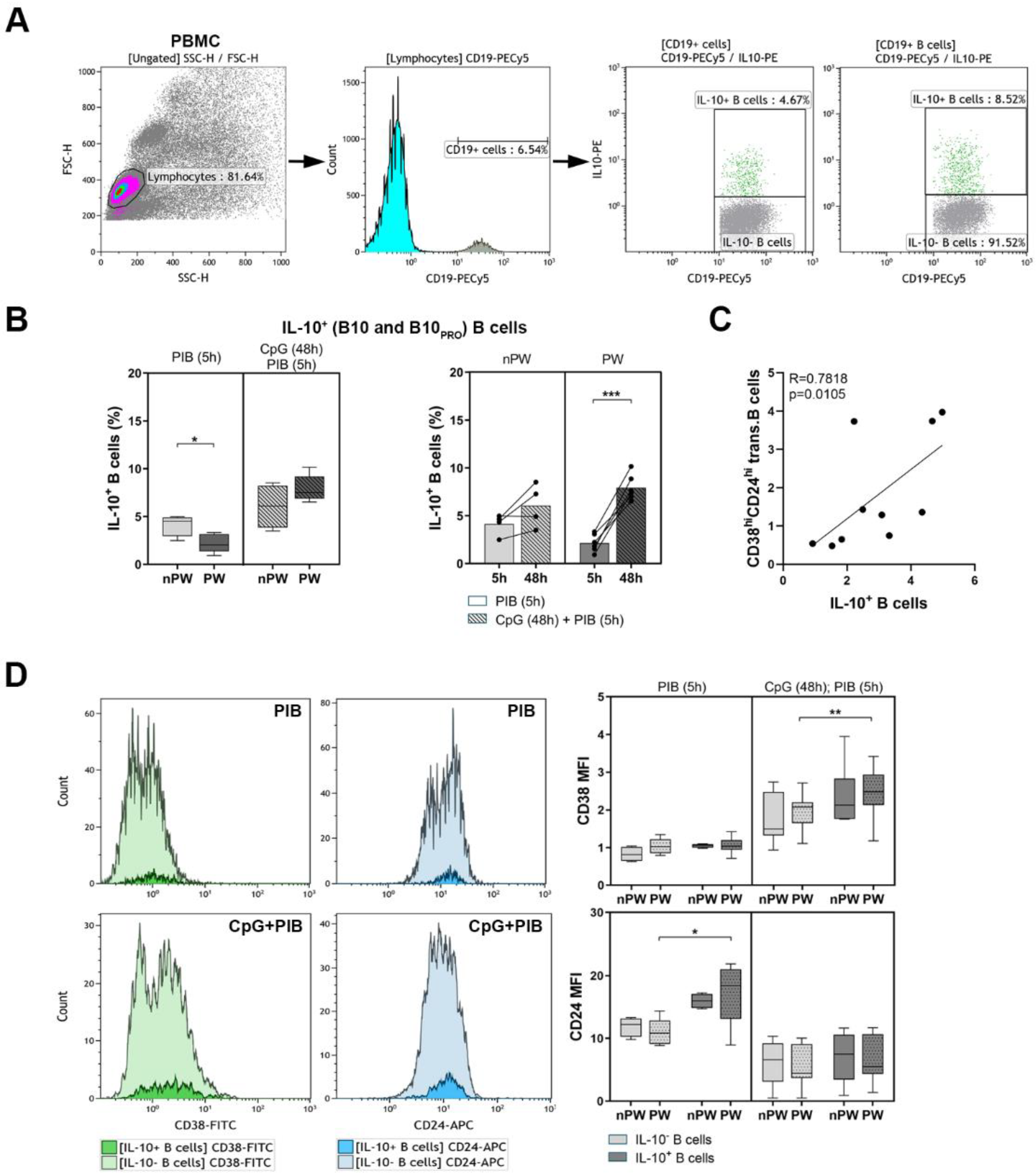
3.2. The Determination of IL-10-Producing B Cells
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lampé, R.; Szűcs, S.; Ádány, R.; Póka, R. Granulocyte superoxide anion production and regulation by plasma factors in normal and preeclamptic pregnancy. J. Reprod. Immunol. 2011, 89, 199–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geldenhuys, J.; Rossouw, T.M.; Lombaard, H.A.; Ehlers, M.M.; Kock, M.M. Disruption in the Regulation of Immune Responses in the Placental Subtype of Preeclampsia. Front. Immunol. 2018, 9, 1659. [Google Scholar] [CrossRef] [PubMed]
- Ghaebi, M.; Nouri, M.; Ghasemzadeh, A.; Farzadi, L.; Jadidi-Niaragh, F.; Ahmadi, M.; Yousefi, M. Immune regulatory network in successful pregnancy and reproductive failures. Biomed. Pharmacother. 2017, 88, 61–73. [Google Scholar] [CrossRef]
- Lu, H.Q.; Hu, R. The role of immunity in the pathogenesis and development of pre-eclampsia. Scand. J. Immunol. 2019, 90, e12756. [Google Scholar] [CrossRef] [PubMed]
- Darmochwal-Kolarz, D.; Leszczynska-Gorzelak, B.; Rolinski, J.; Oleszczuk, J. The immunophenotype of patients with recurrent pregnancy loss. Eur. J. Obstet. Gynecol. Reprod. Biol. 2002, 103, 53–57. [Google Scholar] [CrossRef]
- Sacks, G.P.; Studena, K.; Sargent, K.; Redman, C.W. Normal pregnancy and preeclampsia both produce inflammatory changes in peripheral blood leukocytes akin to those of sepsis. Am. J. Obstet. Gynecol. 1998, 179, 80–86. [Google Scholar] [CrossRef]
- Bonney, E.A. Immune Regulation in Pregnancy: A Matter of Perspective? Obstet. Gynecol. Clin. N. Am. 2016, 43, 679–698. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Dunk, C.E.; Lye, S.J. Sphingosine signalling regulates decidual NK cell angiogenic phenotype and trophoblast migration. Hum. Reprod. 2013, 28, 3026–3037. [Google Scholar] [CrossRef] [Green Version]
- Cervar, M.; Desoye, G.; Blaschitz, A.; Dohr, G. Paracrine regulation of distinct trophoblast functions in vitro by placental macrophages. Cell Tissue Res. 1999, 295, 297–305. [Google Scholar] [CrossRef]
- Nair, R.R.; Verma, P.; Singh, K. Immune-endocrine crosstalk during pregnancy. Gen. Comp. Endocrinol. 2017, 242, 18–23. [Google Scholar] [CrossRef]
- Matalka, K.Z. The effect of estradiol, but not progesterone, on the production of cytokines in stimulated whole blood, is concentration-dependent. Neuro Endocrinol. Lett. 2003, 24, 185–191. [Google Scholar] [PubMed]
- Hughes, G.C.; Clark, E.A.; Wong, A.H. The intracellular progesterone receptor regulates CD4+ T cells and T cell-dependent antibody responses. J. Leukoc. Biol. 2013, 93, 369–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zenclussen, A.C. Adaptive Immune Responses during Pregnancy. Am. J. Reprod. Immunol. 2013, 69, 291–303. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Zhu, X.; Chen, R.; Liu, J.; Liu, P.; Hu, A.; Wu, L.; Hua, H.; Yuan, H. Early Pregnancy Factor Enhances the Generation and Function of CD4+CD25+ Regulatory T Cells. J. Exp. Med. 2016, 240, 215–220. [Google Scholar] [CrossRef] [Green Version]
- Huang, N.; Chi, H.; Qiao, J. Role of Regulatory T Cells in Regulating Fetal-Maternal Immune Tolerance in Healthy Pregnancies and Reproductive Diseases. Front. Immunol. 2020, 11, 1023. [Google Scholar] [CrossRef]
- Adar, T.; Grisaru-Granovsky, S.; Ben Ya’acov, A.; Goldin, E.; Bar-Gil Shitrit, A.; Ben Ya’acov, A. Pregnancy and the Immune System: General Overview and the Gastroenterological Perspective. Dig. Dis. Sci. 2015, 60, 2581–2589. [Google Scholar] [CrossRef]
- Dutta, S.; Sengupta, P.; Haque, N. Reproductive immunomodulatory functions of B cells in pregnancy. Int. Rev. Immunol. 2020, 39, 53–66. [Google Scholar] [CrossRef]
- Nguyen, T.G.; Ward, C.M.; Morris, J.M. To B or not to B cells-mediate a healthy start to life. Clin. Exp. Immunol. 2013, 171, 124–134. [Google Scholar] [CrossRef]
- Muzzio, D.; Zygmunt, M.; Jensen, F. The role of pregnancy-associated hormones in the development and function of regulatory B cells. Front. Endocrinol. 2014, 5, 39. [Google Scholar] [CrossRef] [Green Version]
- Lima, J.; Martins, C.; Leandro, M.J.; Nunes, G.; Sousa, M.J.; Branco, J.C. Characterization of B cells in healthy pregnant women from late pregnancy to post-partum: A prospective observational study. BMC Pregnancy Childbirth 2016, 16, 139. [Google Scholar] [CrossRef] [Green Version]
- Faul, F.; Erdfelder, E.; Lang, A.G.; Buchner, A. G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behav. Res. Methods 2007, 39, 175–191. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, K.B.; Muzzio, D.O.; Matzner, F.; Bommer, I.; Ventimiglia, M.S.; Malinowsky, K.; Ehrhardt, J.; Zygmunt, M.; Jensen, F. Human pregnancy is accompanied by modifications in B cell development and immunoglobulin profile. J. Reprod. Immunol. 2018, 129, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Jensen, F.; Wallukat, G.; Herse, F.; Budner, O.; El-Mousleh, T.; Costa, S.D.; Dechend, R.; Zenclussen, A.C. CD19+CD5+ cells as indicators of preeclampsia. Hypertension 2012, 59, 861–868. [Google Scholar] [CrossRef] [Green Version]
- Abu-Raya, B.; Michalski, C.; Sadarangani, M.; Lavoie, P.M. Maternal Immunological Adaptation during Normal Pregnancy. Front. Immunol. 2020, 11, 575197. [Google Scholar] [CrossRef] [PubMed]
- Martins, C.; Lima, J.; Nunes, G.; Borrego, L.M. Pregnancy alters the circulating B-cell compartment in atopic asthmatic women, and transitional B-cells are positively associated with the development of allergy manifestations in their progeny. Am. J. Reprod. Immunol. 2016, 76, 465–474. [Google Scholar] [CrossRef]
- Ota, K.; Dambaeva, S.; Lee, J.; Gilman-Sachs, A.; Beaman, K.; Kwak-Kim, J. Persistent high levels of IgM antiphospholipid antibodies in a patient with recurrent pregnancy losses and rheumatoid arthritis. Am. J. Reprod. Immunol. 2014, 71, 286–292. [Google Scholar] [CrossRef]
- Sung, N.; Byeon, H.J.; Garcia, M.D.S.; Skariah, A.; Wu, L.; Dambaeva, S.; Beaman, K.; Gilman-Sachs, A.; Kwak-Kim, J. Deficiency in memory B cell compartment in a patient with infertility and recurrent pregnancy losses. J. Reprod. Immunol. 2016, 118, 70–75. [Google Scholar] [CrossRef]
- Zhang, L.; Chang, K.K.; Li, M.Q.; Li, D.J.; Yao, X.Y. Mouse endometrial stromal cells and progesterone inhibit the activation and regulate the differentiation and antibody secretion of mouse B cells. Int. J. Clin. Exp. Pathol. 2014, 7, 123–133. [Google Scholar]
- Ding, T.; Yan, F.; Cao, S.; Ren, X. Regulatory B cell: New member of immunosuppressive cell club. Hum. Immunol. 2015, 76, 615–621. [Google Scholar] [CrossRef]
- Mauri, C.; Menon, M. Human regulatory B cells in health and disease: Therapeutic potential. J. Clin. Investig. 2017, 127, 772–779. [Google Scholar] [CrossRef] [Green Version]
- Floudas, A.; Amu, S.; Fallon, P.G. New Insights into IL-10 Dependent and IL-10 Independent Mechanisms of Regulatory B Cell Immune Suppression. J. Clin. Immunol. 2016, 36, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Papp, G.; Boros, P.; Nakken, B.; Szodoray, P.; Zeher, M. Regulatory immune cells and functions in autoimmunity and transplantation immunology. Autoimmun. Rev. 2017, 16, 435–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esteve-Solé, A.; Luo, Y.; Vlagea, A.; Deyà-Martínez, Á.; Yagüe, J.; Plaza-Martín, A.M.; Juan, M.; Alsina, L. B Regulatory Cells: Players in Pregnancy and Early Life. Int. J. Mol. Sci. 2018, 19, 2099. [Google Scholar] [CrossRef] [Green Version]
- Bhat, N.M.; Mithal, A.; Bieber, M.M.; Herzenberg, L.A.; Teng, N.N. Human CD5+ B lymphocytes (B-1 cells) decrease in peripheral blood during pregnancy. J. Reprod. Immunol. 1995, 28, 53–60. [Google Scholar] [CrossRef]
- Rolle, L.; Memarzadeh Tehran, M.; Morell-García, A.; Raeva, Y.; Schumacher, A.; Hartig, R.; Costa, S.D.; Jensen, F.; Zenclussen, A.C. Cutting edge: IL-10-producing regulatory B cells in early human pregnancy. Am. J. Reprod. Immunol. 2013, 70, 448–453. [Google Scholar] [CrossRef] [PubMed]
- Jensen, F.; Muzzio, D.; Soldati, R.; Fest, S.; Zenclussen, A.C. Regulatory B10 cells restore pregnancy tolerance in a mouse model. Biol. Reprod. 2013, 89, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koushaeian, L.; Ghorbani, F.; Ahmadi, M.; Eghbal-Fard, S.; Zamani, M.; Danaii, S.; Yousefi, B.; Jadidi-Niaragh, F.; Hamdi, K.; Yousefi, M. The role of IL-10-producing B cells in repeated implantation failure patients with cellular immune abnormalities. Immunol. Lett. 2019, 214, 16–22. [Google Scholar] [CrossRef]
- Danaii, S.; Ghorbani, F.; Ahmadi, M.; Abbaszadeh, H.; Koushaeian, L.; Soltani-Zangbar, M.S.; Mehdizadeh, A.; Hojjat-Farsangi, M.; Kafil, H.S.; Aghebati-Maleki, L.; et al. IL-10-producing B cells play important role in the pathogenesis of recurrent pregnancy loss. Int. Immunopharmacol. 2020, 87, 106806. [Google Scholar] [CrossRef]
- Lighaam, L.C.; Unger, P.A.; Vredevoogd, D.W.; Verhoeven, D.; Vermeulen, E.; Turksma, A.W.; Ten Brinke, A.; Rispens, T.; van Ham, S.M. In vitro-Induced Human IL-10+ B Cells Do Not Show a Subset-Defining Marker Signature and Plastically Co-express IL-10 with Pro-Inflammatory Cytokines. Front. Immunol. 2018, 9, 1913. [Google Scholar] [CrossRef] [Green Version]
- Benner, M.; Feyaerts, D.; García, C.C.; Inci, N.; López, S.C.; Fasse, E.; Shadmanfar, W.; van der Heijden, O.W.H.; Gorris, M.A.J.; Joosten, I.; et al. Clusters of Tolerogenic B Cells Feature in the Dynamic Immunological Landscape of the Pregnant Uterus. Cell Rep. 2020, 32, 108204. [Google Scholar] [CrossRef]
- Guzman-Genuino, R.M.; Diener, K.R. Regulatory B Cells in Pregnancy: Lessons from Autoimmunity, Graft Tolerance, and Cancer. Front. Immunol. 2017, 8, 172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mor, G.; Aldo, P.; Alvero, A.B. The unique immunological and microbial aspects of pregnancy. Nat. Rev. Immunol. 2017, 17, 469–482. [Google Scholar] [CrossRef] [PubMed]
- Förger, F.; Villiger, P.M. Immunological adaptations in pregnancy that modulate rheumatoid arthritis disease activity. Nat. Rev. Rheumatol. 2020, 16, 113–122. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kövér, Á.; Lampé, R.; Szabó, K.; Tarr, T.; Papp, G. A Comprehensive Investigation into the Distribution of Circulating B Cell Subsets in the Third Trimester of Pregnancy. J. Clin. Med. 2022, 11, 3006. https://doi.org/10.3390/jcm11113006
Kövér Á, Lampé R, Szabó K, Tarr T, Papp G. A Comprehensive Investigation into the Distribution of Circulating B Cell Subsets in the Third Trimester of Pregnancy. Journal of Clinical Medicine. 2022; 11(11):3006. https://doi.org/10.3390/jcm11113006
Chicago/Turabian StyleKövér, Ágnes, Rudolf Lampé, Krisztina Szabó, Tünde Tarr, and Gábor Papp. 2022. "A Comprehensive Investigation into the Distribution of Circulating B Cell Subsets in the Third Trimester of Pregnancy" Journal of Clinical Medicine 11, no. 11: 3006. https://doi.org/10.3390/jcm11113006
APA StyleKövér, Á., Lampé, R., Szabó, K., Tarr, T., & Papp, G. (2022). A Comprehensive Investigation into the Distribution of Circulating B Cell Subsets in the Third Trimester of Pregnancy. Journal of Clinical Medicine, 11(11), 3006. https://doi.org/10.3390/jcm11113006