
Chemically Induced Colitis-Associated Cancer Models in Rodents for Pharmacological Modulation: A Systematic Review

 , , , ,
, , , , 
Abstract
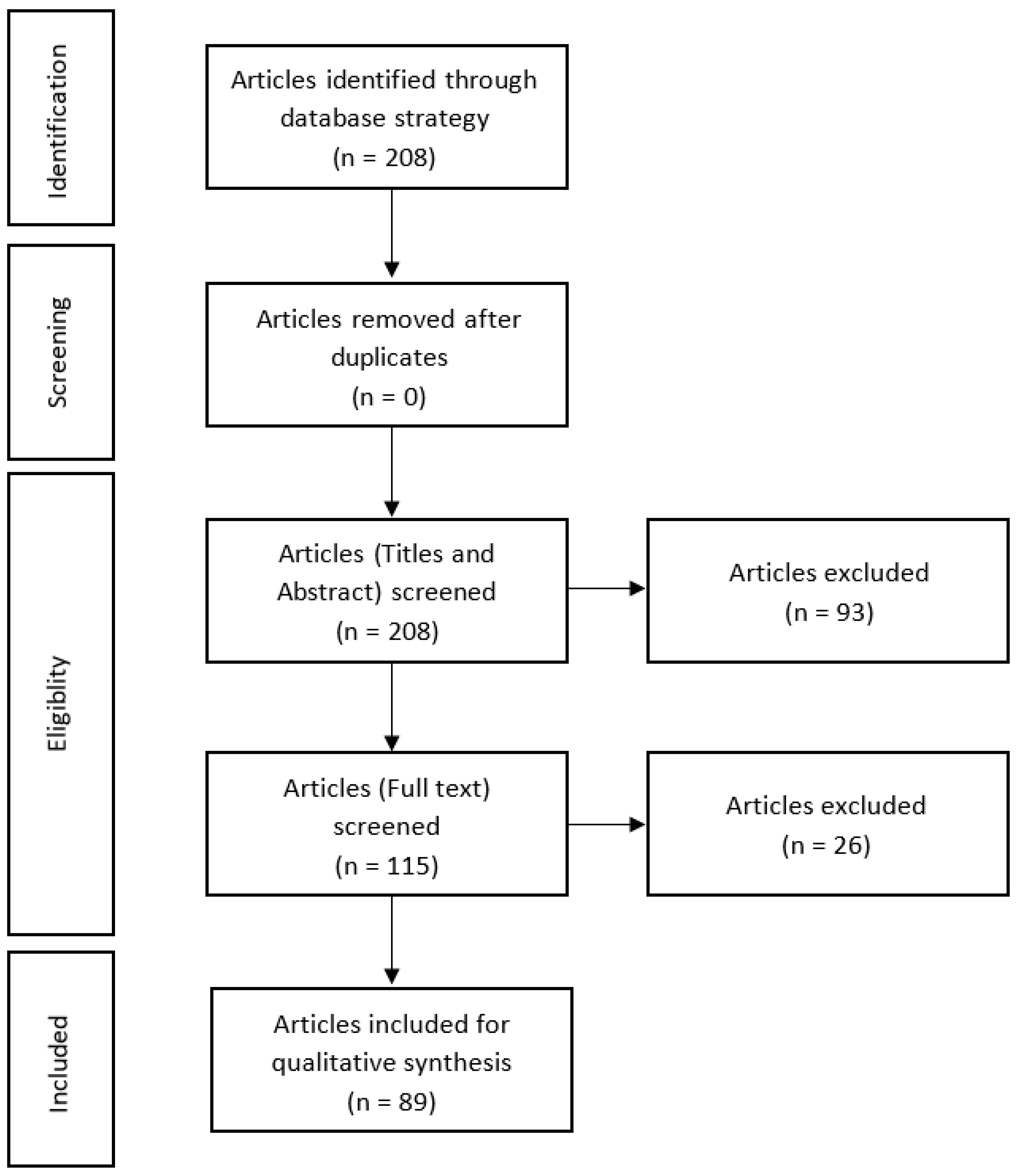
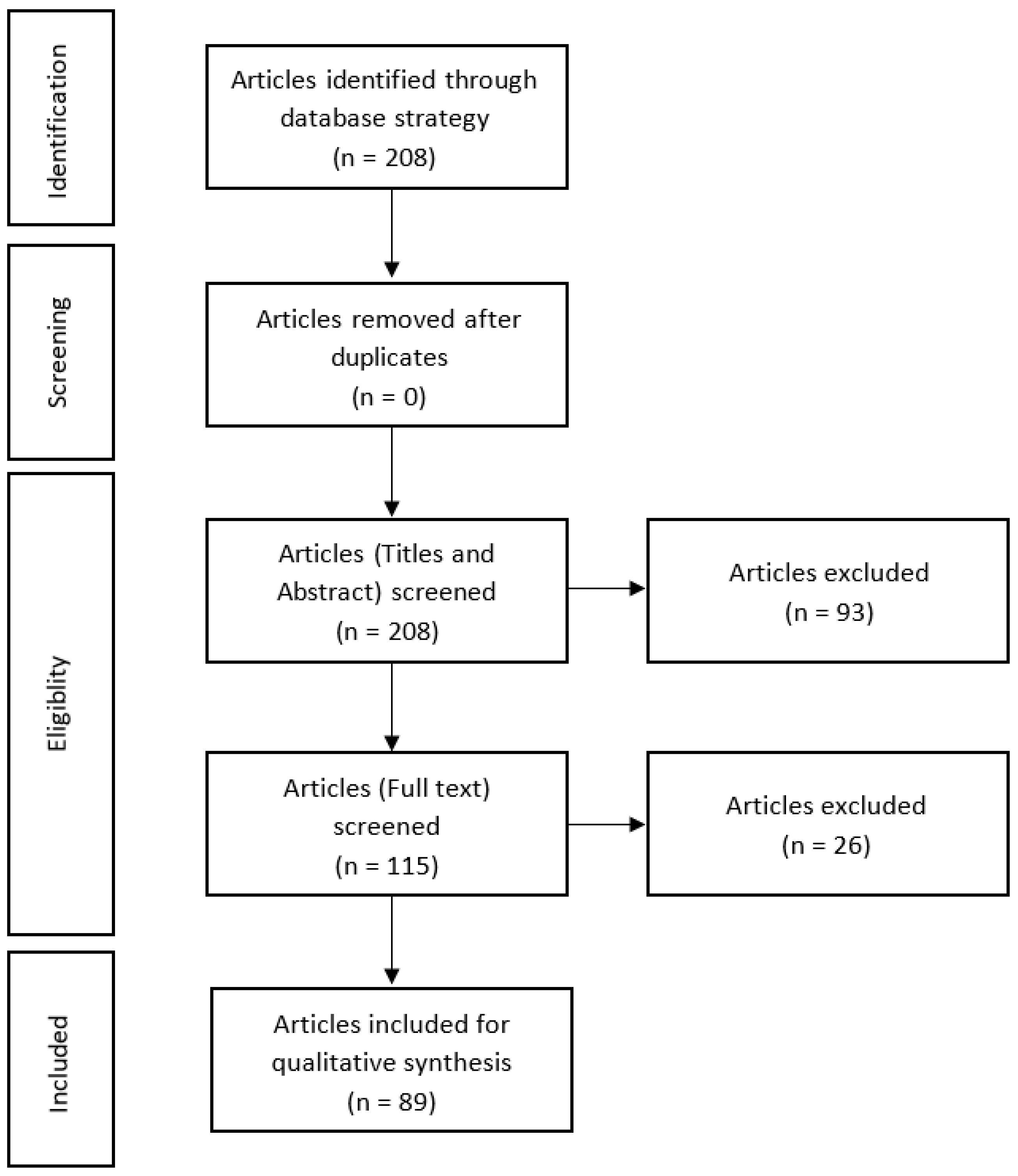
:1. Introduction
2. Materials and Methods
2.1. Search Strategy
2.2. Selection of Studies
2.3. Data Extraction
2.4. Articles Eligibility
3. Results
3.1. Pro-Inflammatory Reagent Related Parameters
3.1.1. Number of TNBS Administrations
3.1.2. TNBS Dose
3.1.3. TNBS Volume
3.1.4. TNBS Vehicle
3.1.5. Number of DSS Administrations
3.1.6. DSS Dose
3.2. Procarcinogen Reagent Related Parameters
3.2.1. Number of AOM Administrations
3.2.2. AOM Dose
3.3. Rodent-Related Parameters
3.3.1. Strain
3.3.2. Gender
3.3.3. Age
3.4. Model Characterization
3.4.1. Clinical Signs and Symptoms
3.4.2. Biochemical Markers
3.4.3. Pro-Inflammatory Markers
3.4.4. Carcinogenesis Markers
3.4.5. Macroscopic Evaluation
3.4.6. Histological Evaluation
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dekker, E.; Tanis, P.J.; Vleugels, J.L.A.; Kasi, P.M.; Wallace, M.B. Colorectal cancer. Lancet 2019, 394, 1467–1480. [Google Scholar] [CrossRef]
- IARC International Agency for Research on Cancer. Cancer Fact Sheets. Available online: http://gco.iarc.fr/today/fact-sheets-cancers (accessed on 3 June 2021).
- Brody, H. Colorectal cancer. Nature 2015, 521, S1. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.; Desantis, C.; Jemal, A. Colorectal cancer statistics, 2014. CA Cancer J. Clin. 2014, 64, 104–117. [Google Scholar] [CrossRef] [PubMed]
- Tommelein, J.; Verset, L.; Boterberg, T.; Demetter, P.; Bracke, M.; De Wever, O. Cancer-Associated Fibroblasts Connect Metastasis-Promoting Communication in Colorectal Cancer. Front. Oncol. 2015, 5, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grizzi, F.; Bianchi, P.; Malesci, A.; Laghi, L. Prognostic value of innate and adaptive immunity in colorectal cancer. World J. Gastroenterol. 2013, 19, 174–184. [Google Scholar] [CrossRef] [PubMed]
- Beaugerie, L.; Itzkowitz, S.H. Cancers Complicating Inflammatory Bowel Disease. N. Engl. J. Med. 2015, 372, 1441–1452. [Google Scholar] [CrossRef]
- Pandey, A.; Shen, C.; Man, S.M. Inflammasomes in colitis and colorectal cancer. Yale J. Biol. Med. 2019, 92, 481–498. [Google Scholar]
- Saxena, A.; Baliga, M.S.; Ponemone, V.; Kaur, K.; Larsen, B.; Fletcher, E.; Greene, J.; Fayad, R. Mucus and adiponectin deficiency: Role in chronic inflammation-induced colon cancer. Int. J. Colorectal Dis. 2013, 28, 1267–1279. [Google Scholar] [CrossRef]
- Weitz, J.; Koch, M.; Debus, J.; Höhler, T.; Galle, P.R.; Büchler, M.W. Colorectal cancer. Lancet 2005, 365, 153–165. [Google Scholar] [CrossRef]
- Kuipers, E.J.; Grady, W.M.; Lieberman, D.; Seufferlein, T.; Sung, J.J.; Boelens, P.G.; van de Velde, C.J.; Watanabe, T. Colorectal cancer. Nat. Rev. Dis. Primers 2015, 1, 15065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De-Souza, A.S.C.; Costa-Casagrande, T.A. Animal Models for Colorectal cancer. ABCD Arq. Bras. Cir. Dig. 2018, 31, e1369. [Google Scholar] [CrossRef] [PubMed]
- Artis, D. Epithelial-cell recognition of commensal bacteria and maintenance of immune homeostasis in the gut. Nat. Rev. Immunol. 2008, 8, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Saxena, A.; Chumanevich, A.; Fletcher, E.; Larsen, B.; Lattwein, K.; Kaur, K.; Fayad, R. Adiponectin deficiency: Role in chronic inflammation induced colon cancer. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2012, 1822, 527–536. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.R.; Chang, D.K. Colorectal cancer in inflammatory bowel disease: The risk, pathogenesis, prevention and diagnosis. World J. Gastroenterol. 2014, 20, 9872–9881. [Google Scholar] [CrossRef] [PubMed]
- Munkholm, P. Review article: The incidence and prevalence of colorectal cancer in inflammatory bowel disease. Aliment. Pharmacol. Ther. 2003, 18, 1–5. [Google Scholar] [CrossRef]
- Derikx, L.A.; Nissen, L.H.; Smits, L.J.; Shen, B.; Hoentjen, F. Risk of Neoplasia After Colectomy in Patients With Inflammatory Bowel Disease: A Systematic Review and Meta-analysis. Clin. Gastroenterol. Hepatol. 2015, 14, 798–806.e20. [Google Scholar] [CrossRef] [Green Version]
- Itzkowitz, S.H.; Harpaz, N. Diagnosis and management of dysplasia in patients with inflammatory bowel diseases. Gastroenterology 2004, 126, 1634–1648. [Google Scholar] [CrossRef]
- Rutter, M.D.; Saunders, B.P.; Wilkinson, K.H.; Rumbles, S.; Schofield, G.; Kamm, M.A.; Williams, C.B.; Price, A.B.; Talbot, I.C.; Forbes, A. Thirty-Year Analysis of a Colonoscopic Surveillance Program for Neoplasia in Ulcerative Colitis. Gastroenterology 2006, 130, 1030–1038. [Google Scholar] [CrossRef]
- Eaden, J.A.; Abrams, K.R.; Mayberry, J.F. The risk of colorectal cancer in ulcerative colitis: A meta-analysis. Gut 2001, 48, 526–535. [Google Scholar] [CrossRef] [Green Version]
- Man, S.M. Inflammasomes in the gastrointestinal tract: Infection, cancer and gut microbiota homeostasis. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 721–737. [Google Scholar] [CrossRef] [PubMed]
- Modest, D.P.; Pant, S.; Sartore-Bianchi, A. Treatment sequencing in metastatic colorectal cancer. Eur. J. Cancer 2019, 109, 70–83. [Google Scholar] [CrossRef] [PubMed]
- Mittal, V.K.; Bhullar, J.S.; Jayant, K. Animal models of human colorectal cancer: Current status, uses and limitations. World J. Gastroenterol. 2015, 21, 11854–11861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanneganti, M.; Mino-Kenudson, M.; Mizoguchi, E. Animal Models of Colitis-Associated Carcinogenesis. J. Biomed. Biotechnol. 2011, 2011, 342637. [Google Scholar] [CrossRef] [Green Version]
- Danese, S.; Mantovani, A. Inflammatory bowel disease and intestinal cancer: A paradigm of the Yin–Yang interplay between inflammation and cancer. Oncogene 2010, 29, 3313–3323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leystra, A.A.; Clapper, M.L. Gut Microbiota Influences Experimental Outcomes in Mouse Models of Colorectal Cancer. Genes 2019, 10, 900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bürtin, F.; Mullins, C.; Linnebacher, M. Mouse models of colorectal cancer: Past, present and future perspectives. World J. Gastroenterol. 2020, 26, 1394–1426. [Google Scholar] [CrossRef]
- Parang, B.; Barrett, C.W.; Williams, C.S. AOM/DSS Model of Colitis-Associated Cancer. In Gastrointestinal Physiology and Diseases; Humana Press: New York, NY, USA, 2016; Volume 1422, pp. 297–307. [Google Scholar] [CrossRef] [Green Version]
- Silva, I.; Pinto, R.; Mateus, V. Preclinical Study in Vivo for New Pharmacological Approaches in Inflammatory Bowel Disease: A Systematic Review of Chronic Model of TNBS-Induced Colitis. J. Clin. Med. 2019, 8, 1574. [Google Scholar] [CrossRef] [Green Version]
- Randhawa, P.K.; Singh, K.; Singh, N.; Jaggi, A.S. A Review on Chemical-Induced Inflammatory Bowel Disease Models in Rodents. Korean J. Physiol. Pharmacol. 2014, 18, 279–288. [Google Scholar] [CrossRef] [Green Version]
- Wirtz, S.; Neurath, M.F. Mouse models of inflammatory bowel disease. Adv. Drug Deliv. Rev. 2007, 59, 1073–1083. [Google Scholar] [CrossRef]
- Bang, B.; Lichtenberger, L.M. Methods of Inducing Inflammatory Bowel Disease in Mice. Curr. Protoc. Pharmacol. 2016, 72, 1–42. [Google Scholar] [CrossRef] [PubMed]
- Qin, H.-Y.; Wu, J.C.Y.; Tong, X.-D.; Sung, J.J.Y.; Xu, H.-X.; Bian, Z.-X. Systematic review of animal models of post-infectious/post-inflammatory irritable bowel syndrome. J. Gastroenterol. 2010, 46, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.K.; Lawrance, I.C. Animal Models of IBD-Associated CRC and Colorectal Cancer Tumorigenesis. Clin. Med. Insights Ther. 2015, 7, CMT-S18489. [Google Scholar] [CrossRef]
- Abbas, A.K.; Murphy, K.M.; Sher, A. Functional diversity of helper T lymphocytes. Nature 1996, 383, 787–793, Erratum in J. Clin. Med. 2019, 8, 1574. [Google Scholar] [CrossRef]
- Antoniou, E.; Margonis, G.A.; Angelou, A.; Pikouli, A.; Argiri, P.; Karavokyros, I.; Papalois, A.; Pikoulis, E. The TNBS-induced colitis animal model: An overview. Ann. Med. Surg. 2016, 11, 9–15. [Google Scholar] [CrossRef]
- Bramhall, M.; Flórez-Vargas, O.; Stevens, R.; Brass, A.; Cruickshank, S. Quality of Methods Reporting in Animal Models of Colitis. Inflamm. Bowel Dis. 2015, 21, 1248–1259. [Google Scholar] [CrossRef] [Green Version]
- Ohkusa, T. Production of experimental ulcerative colitis in hamsters by dextran sulfate sodium and changes in intestinal microflora. Jpn. J. Gastroenterol. 1985, 82, 1327–1336. [Google Scholar]
- Okayasu, I.; Hatakeyama, S.; Yamada, M.; Ohkusa, T.; Inagaki, Y.; Nakaya, R. A novel method in the induction of reliable experimental acute and chronic ulcerative colitis in mice. Gastroenterology 1990, 98, 694–702. [Google Scholar] [CrossRef]
- Gadaleta, R.M.; Garcia-Irigoyen, O.; Moschetta, A. Exploration of Inflammatory Bowel Disease in Mice: Chemically Induced Murine Models of Inflammatory Bowel Disease (IBD). Curr. Protoc. Mouse Biol. 2017, 7, 13–28. [Google Scholar] [CrossRef]
- Thakur, B.K.; Saha, P.; Banik, G.; Saha, D.R.; Grover, S.; Batish, V.K.; Das, S. Live and heat-killed probiotic Lactobacillus casei Lbs2 protects from experimental colitis through Toll-like receptor 2-dependent induction of T-regulatory response. Int. Immunopharmacol. 2016, 36, 39–50. [Google Scholar] [CrossRef]
- Perše, M.; Cerar, A. Dextran Sodium Sulphate Colitis Mouse Model: Traps and Tricks. J. Biomed. Biotechnol. 2012, 2012, 718617. [Google Scholar] [CrossRef] [Green Version]
- Mizoguchi, E.; Nguyen, D.; Low, D. Animal models of ulcerative colitis and their application in drug research. Drug Des. Dev. Ther. 2013, 7, 1341–1357. [Google Scholar] [CrossRef] [Green Version]
- Scheiffele, F.; Fuss, I.J. Induction of TNBS Colitis in Mice. Curr. Protoc. Immunol. 2002, 49, 1–14. [Google Scholar] [CrossRef]
- Aljahdali, N.; Gadonna-Widehem, P.; Delayre-Orthez, C.; Marier, D.; Garnier, B.; Carbonero, F.; Anton, P.M. Repeated Oral Exposure to N ε-Carboxymethyllysine, a Maillard Reaction Product, Alleviates Gut Microbiota Dysbiosis in Colitic Mice. Am. J. Dig. Dis. 2017, 62, 3370–3384. [Google Scholar] [CrossRef]
- Linden, D.R.; Chen, J.-X.; Gershon, M.D.; Sharkey, K.A.; Mawe, G.M. Serotonin availability is increased in mucosa of guinea pigs with TNBS-induced colitis. Am. J. Physiol. Liver Physiol. 2003, 285, G207–G216. [Google Scholar] [CrossRef] [Green Version]
- Linden, D.R.; Foley, K.F.; McQuoid, C.; Simpson, J.; Sharkey, K.A.; Mawe, G.M. Serotonin transporter function and expression are reduced in mice with TNBS-induced colitis. Neurogastroenterol. Motil. 2005, 17, 565–574. [Google Scholar] [CrossRef]
- Chen, J.; Pitmon, E.; Wang, K. Microbiome, Inflammation and Colorectal Cancer. Semin. Immunol. 2017, 32, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; du Sert, N.P.; Vollert, J.; Rice, A.S.C. General Principles of Preclinical Study Design. Handb. Exp. Pharmacol. 2019, 257, 55–69. [Google Scholar] [CrossRef] [Green Version]
- Walter, L.; Pujada, A.; Bhatnagar, N.; Bialkowska, A.B.; Yang, V.W.; Laroui, H.; Garg, P. Epithelial derived-matrix metalloproteinase (MMP9) exhibits a novel defensive role of tumor suppressor in colitis associated cancer by activating MMP9-Notch1-ARF-p53 axis. Oncotarget 2017, 8, 364–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, Y.; Dai, X.; Li, K.; Gui, G.; Liu, J.; Yang, H. Clostridium butyricum partially regulates the development of colitis-associated cancer through miR-200c. Cell. Mol. Biol. 2017, 63, 59–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, H.; Wang, J.; Phan, C.U.; Chen, Q.; Hu, X.; Shao, G.; Zhou, J.; Lai, L.; Tang, G. Cyclodextrin-based host-guest complexes loaded with regorafenib for colorectal cancer treatment. Nat. Commun. 2021, 12, 759. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Bi, Y.; Zhong, J.; Ren, Z.; Liu, Y.; Jia, J.; Yu, M.; Tan, Y.; Zhang, Q.; Yu, X. Pristimerin suppresses colorectal cancer through inhibiting inflammatory responses and Wnt/β-catenin signaling. Toxicol. Appl. Pharmacol. 2020, 386, 114813. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Wang, K.; Wu, W.; Lv, L.; Bian, X.; Yang, L.; Wang, Q.; Li, Y.; Ye, J.; Fang, D.; et al. Administration of Bifidobacterium bifidum CGMCC 15068 modulates gut microbiota and metabolome in azoxymethane (AOM)/dextran sulphate sodium (DSS)-induced colitis-associated colon cancer (CAC) in mice. Appl. Microbiol. Biotechnol. 2020, 104, 5915–5928. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Su, X.; Dai, L.; Chen, N.; Fang, C.; Dong, Z.; Fu, J.; Yu, Y.; Wang, W.; Zhang, H.; et al. Temporal DNA methylation pattern and targeted therapy in colitis-associated cancer. Carcinogenesis 2020, 41, 235–244. [Google Scholar] [CrossRef]
- Josa, V.; Ferenczi, S.; Szalai, R.; Fuder, E.; Kuti, D.; Horvath, K.; Hegedus, N.; Kovacs, T.; Bagamery, G.; Juhasz, B.; et al. Thrombocytosis and Effects of IL-6 Knock-Out in a Colitis-Associated Cancer Model. Int. J. Mol. Sci. 2020, 21, 6218. [Google Scholar] [CrossRef]
- Levi-Galibov, O.; Lavon, H.; Wassermann-Dozorets, R.; Pevsner-Fischer, M.; Mayer, S.; Wershof, E.; Stein, Y.; Brown, L.E.; Zhang, W.; Friedman, G.; et al. Heat Shock Factor 1-dependent extracellular matrix remodeling mediates the transition from chronic intestinal inflammation to colon cancer. Nat. Commun. 2020, 11, 6245. [Google Scholar] [CrossRef]
- Chartier, L.C.; Hebart, M.L.; Howarth, G.S.; Whittaker, A.L.; Mashtoub, S. Affective state determination in a mouse model of colitis-associated colorectal cancer. PLoS ONE 2020, 15, e0228413. [Google Scholar] [CrossRef] [Green Version]
- Chartier, L.C.; Howarth, G.S.; Mashtoub, S. Chemotherapy-induced mucositis development in a murine model of colitis-associated colorectal cancer. Scand. J. Gastroenterol. 2020, 55, 47–54. [Google Scholar] [CrossRef]
- Chen, L.; Chen, M.-Y.; Shao, L.; Zhang, W.; Rao, T.; Zhou, H.-H.; Huang, W.-H. Panax notoginseng saponins prevent colitis-associated colorectal cancer development: The role of gut microbiota. Chin. J. Nat. Med. 2020, 18, 500–507. [Google Scholar] [CrossRef]
- Hwang, S.; Lee, C.G.; Jo, M.; Park, C.O.; Gwon, S.-Y.; Hwang, S.; Yi, H.C.; Lee, S.-Y.; Eom, Y.-B.; Karim, B.; et al. Enterotoxigenic Bacteroides fragilis infection exacerbates tumorigenesis in AOM/DSS mouse model. Int. J. Med Sci. 2020, 17, 145–152. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Liu, Y.; Wang, Y.; Li, X.; Liu, X.; Guo, M.; Tan, Y.; Qin, X.; Wang, X.; Jiang, M. Sucralose Promotes Colitis-Associated Colorectal Cancer Risk in a Murine Model Along With Changes in Microbiota. Front. Oncol. 2020, 10, 710. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Wang, D.; Qu, S.; Zhao, H.; Lin, Y. miR-370-3p Alleviates Ulcerative Colitis-Related Colorectal Cancer in Mice Through Inhibiting the Inflammatory Response and Epithelial-Mesenchymal Transition. Drug Des. Dev. Ther. 2020, 14, 1127–1141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, F.; Liu, M.; Wang, H.; Shi, G.; Chen, B.; Chen, T.; Yuan, X.; Zhu, P.; Zhou, J.; Wang, Q.; et al. Wu Mei Wan attenuates CAC by regulating gut microbiota and the NF-kB/IL6-STAT3 signaling pathway. Biomed. Pharmacother. 2020, 125, 109982. [Google Scholar] [CrossRef]
- Oh, N.S.; Joung, J.Y.; Lee, J.Y.; Kim, Y.J.; Kim, Y.; Kim, S.H. A synbiotic combination of Lactobacillus gasseri 505 and Cudrania tricuspidata leaf extract prevents hepatic toxicity induced by colorectal cancer in mice. J. Dairy Sci. 2020, 103, 2947–2955. [Google Scholar] [CrossRef] [PubMed]
- Angelou, A.; Papalois, A.E.; Antoniou, E.; Wang, J.; Amini, N.; Pikouli, A.; Andreatos, N.; Buettner, S.; Munir, M.; Theodoropoulos, G.; et al. The Interplay Between Innate Immunity (TLR-4) and sCD40L in the Context of an Animal Model of Colitis-associated Cancer. Anticancer Res. 2020, 40, 5457–5462. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, K.; Deguchi, A.; Motoyoshi, A.; Morita, A.; Maebashi, U.; Nakamoto, T.; Kawanishi, S.; Sueyoshi, M.; Nishimura, K.; Takata, K.; et al. Role of transient receptor potential vanilloid subtype 4 in the regulation of azoymethane/dextran sulphate sodium-induced colitis-associated cancer in mice. Eur. J. Pharmacol. 2020, 867, 172853. [Google Scholar] [CrossRef]
- Yang, M.; Zhang, F.; Yang, C.; Wang, L.; Sung, J.; Garg, P.; Zhang, M.; Merlin, D. Oral Targeted Delivery by Nanoparticles Enhances Efficacy of an Hsp90 Inhibitor by Reducing Systemic Exposure in Murine Models of Colitis and Colitis-Associated Cancer. J. Crohns Colitis 2020, 14, 130–141. [Google Scholar] [CrossRef]
- Ganzleben, I.; Hohmann, M.; Grünberg, A.; Gonzales-Menezes, J.; Vieth, M.; Liebing, E.; Günther, C.; Thonn, V.; Beß, D.; Becker, C.; et al. Topical application of Chlorin e6-PVP (Ce6-PVP) for improved endoscopic detection of neoplastic lesions in a murine colitis-associated cancer model. Sci. Rep. 2020, 10, 13129. [Google Scholar] [CrossRef]
- Li, Z.-W.; Sun, B.; Gong, T.; Guo, S.; Zhang, J.; Wang, J.; Sugawara, A.; Jiang, M.; Yan, J.; Gurary, A.; et al. GNAI1 and GNAI3 Reduce Colitis-Associated Tumorigenesis in Mice by Blocking IL6 Signaling and Down-regulating Expression of GNAI2. Gastroenterology 2019, 156, 2297–2312. [Google Scholar] [CrossRef]
- Hua, F.; Shang, S.; Yang, Y.-W.; Zhang, H.-Z.; Xu, T.-L.; Yu, J.-J.; Zhou, D.-D.; Cui, B.; Li, K.; Lv, X.-X.; et al. TRIB3 Interacts With β-Catenin and TCF4 to Increase Stem Cell Features of Colorectal Cancer Stem Cells and Tumorigenesis. Gastroenterology 2019, 156, 708–721.e15. [Google Scholar] [CrossRef] [Green Version]
- Zhu, W.; Miyata, N.; Winter, M.G.; Arenales, A.; Hughes, E.R.; Spiga, L.; Kim, J.; Sifuentes-Dominguez, L.; Starokadomskyy, P.; Gopal, P.; et al. Editing of the gut microbiota reduces carcinogenesis in mouse models of colitis-associated colorectal cancer. J. Exp. Med. 2019, 216, 2378–2393. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.-S.; Zhang, H.-X.; Li, W.-W.; Ran, Y.; Liu, T.-T.; Xiong, M.-G.; Li, Q.-L.; Wang, S.-Y.; Wu, M.; Shu, H.-B.; et al. FAM64A positively regulates STAT3 activity to promote Th17 differentiation and colitis-associated carcinogenesis. Proc. Natl. Acad. Sci. USA 2019, 116, 10447–10452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.-Q.; Cui, S.-X.; Qu, X.-J. Metformin inhibited colitis and colitis-associated cancer (CAC) through protecting mitochondrial structures of colorectal epithelial cells in mice. Cancer Biol. Ther. 2019, 20, 338–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.-L.; Ding, C.-L.; Qian, C.-L.; Qi, Z.-T.; Wang, W. Retinoid acid induced 16 deficiency aggravates colitis and colitis-associated tumorigenesis in mice. Cell Death Dis. 2019, 10, 958. [Google Scholar] [CrossRef]
- Lv, Z.; Wang, Z.; Luo, L.; Chen, Y.; Han, G.; Wang, R.; Xiao, H.; Li, X.; Hou, C.; Feng, J.; et al. Spliceosome protein Eftud2 promotes colitis-associated tumorigenesis by modulating inflammatory response of macrophage. Mucosal Immunol. 2019, 12, 1164–1173. [Google Scholar] [CrossRef]
- Jeyakumar, T.; Fodil, N.; Van Der Kraak, L.; Meunier, C.; Cayrol, R.; McGregor, K.; Langlais, D.; Greenwood, C.M.T.; Beauchemin, N.; Gros, P. Inactivation of Interferon Regulatory Factor 1 Causes Susceptibility to Colitis-Associated Colorectal Cancer. Sci. Rep. 2019, 9, 18897. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.; Lee, Y.-S.; Kang, S.W.; Kim, S.; Kim, T.-Y.; Lee, S.-H.; Hwang, S.W.; Kim, J.; Na Kim, E.; Ju, J.-S.; et al. Loss of PKM2 in Lgr5+ intestinal stem cells promotes colitis-associated colorectal cancer. Sci. Rep. 2019, 9, 6212. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhao, J.; Zhao, Y.; Zong, S.; Tian, Y.; Chen, S.; Li, M.; Liu, H.; Zhang, Q.; Jing, X.; et al. Therapeutic effects of lentinan on inflammatory bowel disease and colitis-associated cancer. J. Cell. Mol. Med. 2019, 23, 750–760. [Google Scholar] [CrossRef]
- Deng, F.; He, S.; Cui, S.; Shi, Y.; Tan, Y.; Li, Z.; Huang, C.; Liu, D.; Zhi, F.; Peng, L. A Molecular Targeted Immunotherapeutic Strategy for Ulcerative Colitis via Dual-targeting Nanoparticles Delivering miR-146b to Intestinal Macrophages. J. Crohns Colitis 2019, 13, 482–494. [Google Scholar] [CrossRef]
- Han, W.; Xie, B.; Li, Y.; Shi, L.; Wan, J.; Chen, X.; Wang, H. Orally Deliverable Nanotherapeutics for the Synergistic Treatment of Colitis-Associated Colorectal Cancer. Theranostics 2019, 9, 7458–7473. [Google Scholar] [CrossRef]
- Kanehara, K.; Ohnuma, S.; Kanazawa, Y.; Sato, K.; Kokubo, S.; Suzuki, H.; Karasawa, H.; Suzuki, T.; Suzuki, C.; Naitoh, T.; et al. The indole compound MA-35 attenuates tumorigenesis in an inflammation-induced colon cancer model. Sci. Rep. 2019, 9, 12739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capuano, A.; Pivetta, E.; Sartori, G.; Bosisio, G.; Favero, A.; Cover, E.; Andreuzzi, E.; Colombatti, A.; Cannizzaro, R.; Scanziani, E.; et al. Abrogation of EMILIN1-β1 integrin interaction promotes experimental colitis and colon carcinogenesis. Matrix Biol. 2019, 83, 97–115. [Google Scholar] [CrossRef]
- Lee, J.G.; Eun, C.S.; Jo, S.V.; Lee, A.-R.; Park, C.H.; Han, D.S. The impact of gut microbiota manipulation with antibiotics on colon tumorigenesis in a murine model. PLoS ONE 2019, 14, e0226907. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Ng, T.T.; Sham, K.W.; Zhang, L.; Chan, M.T.; Wu, W.K.; Cheng, C.H. Bufalin, a Traditional Chinese Medicine Compound, Prevents Tumor Formation in Two Murine Models of Colorectal Cancer. Cancer Prev. Res. 2019, 12, 653–666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gröschel, C.; Prinz-Wohlgenannt, M.; Mesteri, I.; George, S.K.; Trawnicek, L.; Heiden, D.; Aggarwal, A.; Tennakoon, S.; Baumgartner, M.; Gasche, C.; et al. Switching to a Healthy Diet Prevents the Detrimental Effects of Western Diet in a Colitis-Associated Colorectal Cancer Model. Nutrients 2019, 12, 45. [Google Scholar] [CrossRef] [Green Version]
- Fujita, M.; Ito-Fujita, Y.; Iyoda, T.; Sasada, M.; Okada, Y.; Ishibashi, K.; Osawa, T.; Kodama, H.; Fukai, F.; Suzuki, H. Peptide TNIIIA2 Derived from Tenascin-C Contributes to Malignant Progression in Colitis-Associated Colorectal Cancer via β1-Integrin Activation in Fibroblasts. Int. J. Mol. Sci. 2019, 20, 2752. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Ouyang, M.; Guo, Q.; Jia, J.; Liu, R.; Jiang, Y.; Wu, M.; Shen, S. Changes in the intestinal microecology induced by bacillus subtilis inhibit the occurrence of ulcerative colitis and associated cancers: A study on the mechanisms. Am. J. Cancer Res. 2019, 9, 872–886. [Google Scholar]
- Wunderlich, C.M.; Ackermann, P.J.; Ostermann, A.L.; Adams-Quack, P.; Vogt, M.C.; Tran, M.L.; Nikolajev, A.; Waisman, A.; Garbers, C.; Theurich, S.; et al. Obesity exacerbates colitis-associated cancer via IL-6-regulated macrophage polarisation and CCL-20/CCR-6-mediated lymphocyte recruitment. Nat. Commun. 2018, 9, 1646. [Google Scholar] [CrossRef]
- Lee, Y.K.; Mehrabian, P.; Boyajian, S.; Wu, W.-L.; Selicha, J.; Vonderfecht, S.; Mazmanian, S.K. The Protective Role of Bacteroides fragilis in a Murine Model of Colitis-Associated Colorectal Cancer. mSphere 2018, 3, e00587-18. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Gao, X.; Yu, T.; Yuan, L.; Dai, J.; Wang, W.; Chen, G.; Jiao, C.; Zhou, W.; Huang, Q.; et al. REGγ Controls Hippo Signaling and Reciprocal NF-κB–YAP Regulation to Promote Colon Cancer. Clin. Cancer Res. 2018, 24, 2015–2025. [Google Scholar] [CrossRef] [Green Version]
- Deng, F.; Peng, L.; Li, Z.; Tan, G.; Liang, E.; Chen, S.; Zhao, X.; Zhi, F. YAP triggers the Wnt/β-catenin signalling pathway and promotes enterocyte self-renewal, regeneration and tumorigenesis after DSS-induced injury. Cell Death Dis. 2018, 9, 153. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.Q.; Nie, S.P.; Shen, M.Y.; Hu, J.L.; Yu, Q.; Gong, D.; Xie, M.Y. Tea Polysaccharides Inhibit Colitis-Associated Colorectal Cancer via Interleukin-6/STAT3 Pathway. J. Agric. Food Chem. 2018, 66, 4384–4393. [Google Scholar] [CrossRef] [PubMed]
- Servais, L.; Wéra, O.; Epoh, J.D.; Delierneux, C.; Bouznad, N.; Rahmouni, S.; Mazzucchelli, G.; Baiwir, D.; Delvenne, P.; Lancellotti, P.; et al. Platelets contribute to the initiation of colitis-associated cancer by promoting immunosuppression. J. Thromb. Haemost. 2018, 16, 762–777. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Wang, W.; Shen, B.; Jia, H.; Hou, Z.; Chen, P.; Sun, Y. Pretreatment with probiotic Bifico ameliorates colitis-associated cancer in mice: Transcriptome and gut flora profiling. Cancer Sci. 2018, 109, 666–677. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.-Q.; Li, H.-S.; Nie, S.-P.; Shen, M.-Y.; Hu, J.-L.; Xie, M.-Y. Tea Polysaccharide Prevents Colitis-Associated Carcinogenesis in Mice by Inhibiting the Proliferation and Invasion of Tumor Cells. Int. J. Mol. Sci. 2018, 19, 506. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.-Y.; Yu, L.C.-H.; Yu, I.-S.; Jhuang, Y.-L.; Huang, W.-J.; Yang, C.-Y.; Jeng, Y.-M. Deletion of cadherin-17 enhances intestinal permeability and susceptibility to intestinal tumour formation. J. Pathol. 2018, 246, 289–299. [Google Scholar] [CrossRef]
- Ma, J.; Yang, Y.; Fu, Y.; Guo, F.; Zhang, X.; Xiao, S.; Zhu, W.; Huang, Z.; Zhang, J.; Chen, J. PIAS3-mediated feedback loops promote chronic colitis-associated malignant transformation. Theranostics 2018, 8, 3022–3037. [Google Scholar] [CrossRef]
- Niu, W.; Wu, Z.; Wang, J.; Zhang, H.; Jia, W.; Yang, M.; Luo, Y.; Zhang, X. Tumor Necrosis Factor Ligand-Related Molecule 1A Regulates the Occurrence of Colitis-Associated Colorectal Cancer. Am. J. Dig. Dis. 2018, 63, 2341–2350. [Google Scholar] [CrossRef]
- Amerizadeh, F.; Rezaei, N.; Rahmani, F.; Hassanian, S.M.; Moradi-Marjaneh, R.; Fiuji, H.; Boroumand, N.; Nosrati-Tirkani, A.; Ghayour-Mobarhan, M.; Ferns, G.A.; et al. Crocin synergistically enhances the antiproliferative activity of 5-flurouracil through Wnt/PI3K pathway in a mouse model of colitis-associated colorectal cancer. J. Cell Biochem. 2018, 119, 10250–10261. [Google Scholar] [CrossRef]
- Zhu, Y.; Zhou, J.; Feng, Y.; Chen, L.; Zhang, L.; Yang, F.; Zha, H.; Wang, X.; Han, X.; Shu, C.; et al. Control of Intestinal Inflammation, Colitis-Associated Tumorigenesis, and Macrophage Polarization by Fibrinogen-Like Protein 2. Front. Immunol. 2018, 9, 87. [Google Scholar] [CrossRef] [Green Version]
- Dubé, P.E.; Liu, C.Y.; Girish, N.; Washington, M.K.; Polk, D.B. Pharmacological activation of epidermal growth factor receptor signaling inhibits colitis-associated cancer in mice. Sci. Rep. 2018, 8, 9119. [Google Scholar] [CrossRef] [PubMed]
- Sharp, S.; Malizia, R.A.; Walrath, T.; D’Souza, S.S.; Booth, C.J.; Kartchner, B.J.; Lee, E.C.; Stain, S.C.; O’Connor, W. DNA damage response genes mark the early transition from colitis to neoplasia in colitis-associated colon cancer. Gene 2018, 677, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Song, Z.-Y.; Qu, X.-J.; Li, F.; Zhang, L.; Li, W.-B.; Cui, S.-X. M10, a novel derivative of Myricetin, prevents ulcerative colitis and colorectal tumor through attenuating robust endoplasmic reticulum stress. Carcinogenesis 2018, 39, 889–899. [Google Scholar] [CrossRef] [PubMed]
- Yuan, G.; Chen, T.; Zhang, H.; Cao, Q.; Qiu, Y.; Que, B.; Peng, S.; Chen, M.; Ji, W. Comprehensive analysis of differential circular RNA expression in a mouse model of colitis-induced colon carcinoma. Mol. Carcinog. 2018, 57, 1825–1834. [Google Scholar] [CrossRef] [PubMed]
- Masuda, J.; Umemura, C.; Yokozawa, M.; Yamauchi, K.; Seko, T.; Yamashita, M.; Yamashita, Y. Dietary Supplementation of Selenoneine-Containing Tuna Dark Muscle Extract Effectively Reduces Pathology of Experimental Colorectal Cancers in Mice. Nutrients 2018, 10, 1380. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Wei, W.; Li, X.; Shen, P.; Ju, D.; Wang, Z.; Zhang, R.; Yang, F.; Chen, C.; Cao, K.; et al. BMI1 and MEL18 Promote Colitis-Associated Cancer in Mice via REG3B and STAT3. Gastroenterology 2017, 153, 1607–1620. [Google Scholar] [CrossRef] [Green Version]
- Viennois, E.; Merlin, D.; Gewirtz, A.T.; Chassaing, B. Dietary Emulsifier–Induced Low-Grade Inflammation Promotes Colon Carcinogenesis. Cancer Res. 2017, 77, 27–40. [Google Scholar] [CrossRef] [Green Version]
- Menghini, P.; Di Martino, L.; Lopetuso, L.R.; Corridoni, D.; Webster, J.C.; Xin, W.; Arseneau, K.O.; Lam, M.; Pizarro, T.T.; Cominelli, F. A novel model of colitis-associated cancer in SAMP1/YitFc mice with Crohn’s disease-like ileitis. PLoS ONE 2017, 12, e0174121. [Google Scholar] [CrossRef]
- Lai, C.S.; Yang, G.; Li, S.; Lee, P.S.; Wang, B.N.; Chung, M.C.; Nagabhushanam, K.; Ho, C.T.; Pan, M.H. 3′-Hydroxypterostilbene Suppresses Colitis-Associated Tumorigenesis by Inhibition of IL-6/STAT3 Signaling in Mice. J. Agric. Food Chem. 2017, 65, 9655–9664. [Google Scholar] [CrossRef]
- Zhu, H.; Xu, W.-Y.; Hu, Z.; Zhang, H.; Shen, Y.; Lu, S.; Wei, C.; Wang, Z.-G. RNA virus receptor Rig-I monitors gut microbiota and inhibits colitis-associated colorectal cancer. J. Exp. Clin. Cancer Res. 2017, 36, 2. [Google Scholar] [CrossRef] [Green Version]
- Hennenberg, E.M.; Eyking, A.; Reis, H.; Cario, E. MDR1A deficiency restrains tumor growth in murine colitis-associated carcinogenesis. PLoS ONE 2017, 12, e0180834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Guo, W.; Shen, L.; Chen, Z.; Luo, Q.; Luo, X.; Feng, G.; Shu, Y.; Gu, Y.; Xu, Q.; et al. T lymphocyte SHP2-deficiency triggers anti-tumor immunity to inhibit colitis-associated cancer in mice. Oncotarget 2017, 8, 7586–7597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, T.; Dang, S.; Zhu, R.; Wang, Y.; Nie, Z.; Hong, T.; Zhang, W. Adamts18 deficiency promotes colon carcinogenesis by enhancing β-catenin and p38MAPK/ERK1/2 signaling in the mouse model of AOM/DSS-induced colitis-associated colorectal cancer. Oncotarget 2017, 8, 18979–18990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rothschild, D.E.; Zhang, Y.; Diao, N.; Lee, C.K.; Chen, K.; Caswell, C.C.; Slade, D.J.; Helm, R.F.; LeRoith, T.; Li, L.; et al. Enhanced Mucosal Defense and Reduced Tumor Burden in Mice with the Compromised Negative Regulator IRAK-M. eBioMedicine 2017, 15, 36–47. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Bai, J.; Zhang, L.; Lou, F.; Ke, F.; Cai, W.; Wang, H. Conditional knockout of microRNA-31 promotes the development of colitis associated cancer. Biochem. Biophys. Res. Commun. 2017, 490, 62–68. [Google Scholar] [CrossRef]
- Marelli, G.; Erreni, M.; Anselmo, A.; Taverniti, V.; Guglielmetti, S.; Mantovani, A.; Allavena, P. Heme-oxygenase-1 Production by Intestinal CX3CR1+ Macrophages Helps to Resolve Inflammation and Prevents Carcinogenesis. Cancer Res. 2017, 77, 4472–4485. [Google Scholar] [CrossRef] [Green Version]
- Wei, T.-T.; Lin, Y.-T.; Tseng, R.-Y.; Shun, C.-T.; Lin, Y.-C.; Wu, M.-S.; Fang, J.-M.; Chen, C.-C. Prevention of Colitis and Colitis-Associated Colorectal Cancer by a Novel Polypharmacological Histone Deacetylase Inhibitor. Clin. Cancer Res. 2016, 22, 4158–4169. [Google Scholar] [CrossRef] [Green Version]
- Giner, E.; Recio, M.C.; Ríos, J.L.; Cerdá-Nicolás, J.M.; Giner, R.M. Chemopreventive effect of oleuropein in colitis-associated colorectal cancer in c57bl/6 mice. Mol. Nutr. Food Res. 2016, 60, 242–255. [Google Scholar] [CrossRef]
- Das, S.; Rachagani, S.; Sheinin, Y.; Smith, L.M.; Gurumurthy, C.B.; Roy, H.K.; Batra, S.K. Mice deficient in Muc4 are resistant to experimental colitis and colitis-associated colorectal cancer. Oncogene 2016, 35, 2645–2654. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Xu, R.; Snider, A.J.; Schrandt, J.; Li, Y.; Bialkowska, A.B.; Li, M.; Zhou, J.; Hannun, Y.A.; Obeid, L.M.; et al. Alkaline ceramidase 3 deficiency aggravates colitis and colitis-associated tumorigenesis in mice by hyperactivating the innate immune system. Cell Death Dis. 2016, 7, e2124. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.; Wu, Y.; Deng, B.; Li, J.; Cao, H.; Qu, Y.; Qian, X.; Zhong, G. Isoliquiritigenin decreases the incidence of colitis-associated colorectal cancer by modulating the intestinal microbiota. Oncotarget 2016, 7, 85318–85331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Martino, L.; Dave, M.; Menghini, P.; Xin, W.; Arseneau, K.O.; Pizarro, T.T.; Cominelli, F. Protective Role for TWEAK/Fn14 in Regulating Acute Intestinal Inflammation and Colitis-Associated Tumorigenesis. Cancer Res. 2016, 76, 6533–6542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.; Yang, Y.; Hu, C.; Cheng, X.; Xu, Y.; Cai, X.; Wang, M.; Yang, C.S.; Cao, P. Protective effects of Huangqin Decoction against ulcerative colitis and associated cancer in mice. Oncotarget 2016, 7, 61643–61655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattaveewong, T.; Wongkrasant, P.; Chanchai, S.; Pichyangkura, R.; Chatsudthipong, V.; Muanprasat, C. Chitosan oligosaccharide suppresses tumor progression in a mouse model of colitis-associated colorectal cancer through AMPK activation and suppression of NF-κB and mTOR signaling. Carbohydr. Polym. 2016, 145, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Urrutia-Ortega, I.M.; Garduño-Balderas, L.G.; Delgado-Buenrostro, N.L.; Freyre-Fonseca, V.; Flores-Flores, J.O.; González-Robles, A.; Pedraza-Chaverri, J.; Hernández-Pando, R.; Rodríguez-Sosa, M.; León-Cabrera, S.; et al. Food-grade titanium dioxide exposure exacerbates tumor formation in colitis associated cancer model. Food Chem. Toxicol. 2016, 93, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.M.; Kim, N.; Son, H.J.; Park, J.H.; Nam, R.H.; Ham, M.H.; Choi, D.; Sohn, S.H.; Shin, E.; Hwang, Y.-J.; et al. The Effect of Sex on the Azoxymethane/Dextran Sulfate Sodium-treated Mice Model of Colon Cancer. J. Cancer Prev. 2016, 21, 271–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, L.; Jiang, F.C.; Zhang, L.M.; He, W.T.; Liu, J.H.; Li, M.Q.; Zhang, X.; Xing, S.; Guo, H.; Zhou, P. Targeting of MyD88 Homodimerization by Novel Synthetic Inhibitor TJ-M2010-5 in Preventing Colitis-Associated Colorectal Cancer. J. Natl. Cancer Inst. 2016, 108, djv364. [Google Scholar] [CrossRef] [Green Version]
- Kangwan, N.; Kim, Y.-J.; Han, Y.-M.; Jeong, M.; Park, J.-M.; Hahm, K.-B. Concerted actions of ameliorated colitis, aberrant crypt foci inhibition and 15-hydroxyprostaglandin dehydrogenase induction by sonic hedgehog inhibitor led to prevention of colitis-associated cancer. Int. J. Cancer 2016, 138, 1482–1493. [Google Scholar] [CrossRef] [Green Version]
- Koblansky, A.A.; Truax, A.D.; Liu, R.; Montgomery, S.A.; Ding, S.; Wilson, J.E.; Brickey, W.J.; Mühlbauer, M.; McFadden, R.-M.T.; Hu, P.; et al. The Innate Immune Receptor NLRX1 Functions as a Tumor Suppressor by Reducing Colon Tumorigenesis and Key Tumor-Promoting Signals. Cell Rep. 2016, 14, 2562–2575. [Google Scholar] [CrossRef] [Green Version]
- Sharma, D.; Malik, A.; Guy, C.S.; Karki, R.; Vogel, P.; Kanneganti, T.-D. Pyrin Inflammasome Regulates Tight Junction Integrity to Restrict Colitis and Tumorigenesis. Gastroenterology 2018, 154, 948–964.e8. [Google Scholar] [CrossRef]
- Fung, K.Y.; Putoczki, T. In Vivo Models of Inflammatory Bowel Disease and Colitis-Associated Cancer. In Inflammation and Cancer. Methods in Molecular Biology; Humana Press: New York, NY, USA, 2018; Volume 1725, pp. 3–13. [Google Scholar] [CrossRef]
- Olesch, C.; Sirait-Fischer, E.; Berkefeld, M.; Fink, A.F.; Susen, R.M.; Ritter, B.; Michels, B.E.; Steinhilber, D.; Greten, F.R.; Savai, R.; et al. S1PR4 ablation reduces tumor growth and improves chemotherapy via CD8+ T cell expansion. J. Clin. Investig. 2020, 130, 5461–5476. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Li, B.; Yang, X.; Liu, T.; Shi, T.; Deng, B.; Zhang, Y.; Jia, L.; Jiang, Z.; He, R. Non-hematopoietic STAT6 induces epithelial tight junction dysfunction and promotes intestinal inflammation and tumorigenesis. Mucosal Immunol. 2019, 12, 1304–1315. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Pan, W.M.; Zhang, H.H.; Song, Y.; Chen, J.; Xiang, Y.; Gu, B.; Li, S.Z.; Du, R.L.; Zhang, X.D. Cancer testis antigen 55 deficiency attenuates colitis-associated colorectal cancer by inhibiting NF-κB signaling. Cell Death Dis. 2019, 10, 304. [Google Scholar] [CrossRef] [Green Version]
- Flood, B.; Manils, J.; Nulty, C.; Flis, E.; Kenealy, S.; Barber, G.; Fay, J.; Mills, K.; Kay, E.W.; Creagh, E.M. Caspase-11 regulates the tumour suppressor function of STAT1 in a murine model of colitis-associated carcinogenesis. Oncogene 2019, 38, 2658–2674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amini-Khoei, H.; Momeny, M.; Abdollahi, A.; Dehpour, A.R.; Amiri, S.; Haj-Mirzaian, A.; Tavangar, S.M.; Ghaffari, S.H.; Rahimian, R.; Mehr, S.E. Tropisetron suppresses colitis-associated cancer in a mouse model in the remission stage. Int. Immunopharmacol. 2016, 36, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Hammad, A.; Zheng, Z.-H.; Gao, Y.; Namani, A.; Shi, H.-F.; Tang, X. Identification of novel Nrf2 target genes as prognostic biomarkers in colitis-associated colorectal cancer in Nrf2-deficient mice. Life Sci. 2019, 238, 116968. [Google Scholar] [CrossRef] [PubMed]
- Patil, M.V.K.; Kandhare, A.D.; Bhise, S.D. Anti-inflammatory effect of daucus carota root on experimental colitis in rats. Int. J. Pharm. Pharm. Sci. 2012, 4, 337–343. [Google Scholar]
- Jurjus, A.R.; Khoury, N.N.; Reimund, J.M. Animal models of inflammatory bowel disease. J. Pharmacol. Toxicol. Methods 2004, 50, 81–92. [Google Scholar] [CrossRef]
- Elson, C.O.; Beagley, K.W.; Sharmanov, A.T.; Fujihashi, K.; Kiyono, H.; Tennyson, G.S.; Cong, Y.; Black, C.A.; Ridwan, B.W.; McGhee, J.R. Hapten-induced model of murine inflammatory bowel disease: Mucosa immune responses and protection by tolerance. J. Immunol. 1996, 157, 2174–2185. [Google Scholar]
- Radi, Z.A.; Heuvelman, D.M.; Masferrer, J.L.; Benson, E.L. Pharmacologic evaluation of sulfasalazine, FTY720, and anti-IL-12/23p40 in a TNBS-induced crohn’s disease model. Dig. Dis. Sci. 2011, 56, 2283–2291. [Google Scholar] [CrossRef]
- Lamb, K.; Zhong, F.; Gebhart, G.F.; Bielefeldt, K. Experimental colitis in mice and sensitization of converging visceral and somatic afferent pathways. Am. J. Physiol. Gastrointest. Liver Physiol. 2006, 290, G451–G457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, G.P.; Beck, P.L.; Herridge, M.S.; Depew, W.T.; Szewczuk, M.R.; Wallace, J.L. Hapten-induced model of chronic inflammation and ulceration in the rat colon. Gastroenterology 1989, 96, 795–803. [Google Scholar] [CrossRef]
- Wirtz, S.; Popp, V.; Kindermann, M.; Gerlach, K.; Weigmann, B.; Fichtner-Feigl, S.; Neurath, M.F. Chemically induced mouse models of acute and chronic intestinal inflammation. Nat. Protoc. 2017, 12, 1295–1309. [Google Scholar] [CrossRef]
- Institute for Laboratory Animal Research. Guide for the Care and Use of Laboratory Animals, 8th ed.; National Academies Press: Washington, DC, USA, 2011; pp. 105–133. [Google Scholar]
- Mateus, V.; Rocha, J.; Alves, P.; Mota-Filipe, H.; Sepodes, B.; Pinto, R.M.A. Anti-Inflammatory Effect of Erythropoietin in the TNBS-induced Colitis. Basic Clin. Pharmacol. Toxicol. 2016, 120, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Mateus, V.; Rocha, J.; Alves, P.; Mota-Filipe, H.; Sepodes, B.; Pinto, R. Thiadiazolidinone-8 Ameliorates Inflammation Associated with Experimental Colitis in Mice. Pharmacology 2017, 101, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Mateus, V.; Rocha, J.; Mota-Filipe, H.; Sepodes, B.; Pinto, R. Hemin reduces inflammation associated with TNBS-induced colitis. Clin. Exp. Gastroenterol. 2018, 11, 325–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocha, J.; Eduardo-Figueira, M.; Barateiro, A.; Fernandes, A.; Brites, D.; Bronze, R.; Duarte, C.M.; Serra, A.T.; Pinto, R.; Freitas, M.; et al. Anti-Inflammatory effect of rosmarinic acid and an extract of Rosmarinus officinalis in rat models of local and systemic inflammation. Basic Clin. Pharmacol. Toxicol. 2015, 116, 398–413. [Google Scholar] [CrossRef] [PubMed]
- Direito, R.; Rocha, J.; Lima, A.; Gonçalves, M.M.; Duarte, M.P.; Mateus, V.; Sousa, C.; Fernandes, A.; Pinto, R.; Ferreira, R.B.; et al. Reduction of Inflammation and Colon Injury by a Spearmint Phenolic Extract in Experimental Bowel Disease in Mice. Medicines 2019, 6, 65. [Google Scholar] [CrossRef] [Green Version]
- Direito, R.; Lima, A.; Rocha, J.; Ferreira, R.B.; Mota, J.; Rebelo, P.; Fernandes, A.; Pinto, R.; Alves, P.; Bronze, R.; et al. Dyospiros kaki phenolics inhibit colitis and colon cancer cell proliferation, but not gelatinase activities. J. Nutr. Biochem. 2017, 46, 100–108. [Google Scholar] [CrossRef]
- Hua, H.; Kong, Q.; Zhang, H.; Wang, J.; Luo, T.; Jiang, Y. Targeting mTOR for cancer therapy. J. Hematol. Oncol. 2019, 12, 71. [Google Scholar] [CrossRef]
- Narayanankutty, A. PI3K/Akt/mTOR Pathway as a Therapeutic Target for Colorectal Cancer: A Review of Preclinical and Clinical Evidence. Curr. Drug Targets 2019, 20, 1217–1226. [Google Scholar] [CrossRef] [PubMed]
- Holst, I.; Kleinschmidt, S.; Nolte, I.; Hewicker-Trautwein, M. Expression of Inducible Nitric Oxide, Nitrotyrosine and Manganese Superoxide Dismutase in Dogs with Inflammatory Bowel Disease. J. Comp. Pathol. 2012, 146, 76. [Google Scholar] [CrossRef]
- Mansfield, K.G.; Lin, K.C.; Xia, D.; Newman, J.V.; Schauer, D.B.; MacKey, J.; Lackner, A.A.; Carville, A. Enteropathogenic Escherichia coli and ulcerative colitis in cotton-top tamarins (Saguinus oedipus). J. Infect. Dis. 2001, 184, 803–807. [Google Scholar] [CrossRef] [Green Version]
- Padua, D.; Vu, J.P.; Germano, P.M.; Pisegna, J.R. The Role of Neuropeptides in Mouse Models of Colitis. J. Mol. Neurosci. 2015, 59, 203–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corica, D.; Romano, C. Renal Involvement in Inflammatory Bowel Diseases. J. Crohns Colitis 2015, 10, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Oikonomou, K.; Kapsoritakis, A.; Eleftheriadis, T.; Stefanidis, I.; Potamianos, S. Renal manifestations and complications of inflammatory bowel disease. Inflamm. Bowel Dis. 2011, 17, 1034–1045. [Google Scholar] [CrossRef] [PubMed]
- Rojas-Feria, M.; Castro, M.; Suárez, E.; Ampuero, J.; Romero-Gómez, M. Hepatobiliary manifestations in inflammatory bowel disease: The gut, the drugs and the liver. World J. Gastroenterol. 2013, 19, 7327–7340. [Google Scholar] [CrossRef] [PubMed]
- Hirata, I.; Hoshimoto, M.; Saito, O.; Kayazawa, M.; Nishikawa, T.; Murano, M.; Toshina, K.; Wang, F.-Y.; Matsuse, R. Usefulness of fecal lactoferrin and hemoglobin in diagnosis of colorectal diseases. World J. Gastroenterol. 2007, 13, 1569–1574. [Google Scholar] [CrossRef] [Green Version]
- Jagtap, A.G.; Niphadkar, P.V.; Phadke, A.S. Protective effect of aqueous extract of Bombax malabaricum DC on experimental models of inflammatory bowel disease in rats and mice. Indian J. Exp. Biol. 2011, 49, 343–351. [Google Scholar]
- Mooiweer, E.; Fidder, H.H.; Siersema, P.D.; Laheij, R.J.F.; Oldenburg, B. Fecal Hemoglobin and Calprotectin Are Equally Effective in Identifying Patients with Inflammatory Bowel Disease with Active Endoscopic Inflammation. Inflamm. Bowel Dis. 2014, 20, 307–314. [Google Scholar] [CrossRef]
- Yarur, A.J.; Czul, F.; Levy, C. Hepatobiliary Manifestations of Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2014, 20, 1655–1667. [Google Scholar] [CrossRef] [PubMed]
- Carty, E.; De Brabander, M.; Feakins, R.M.; Rampton, D.S. Measurement of in vivo rectal mucosal cytokine and eicosanoid production in ulcerative colitis using filter paper. Gut 2000, 46, 487–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nairz, M.; Schroll, A.; Moschen, A.; Sonnweber, T.; Theurl, M.; Theurl, I.; Taub, N.; Jamnig, C.; Neurather, D.; Huber, L.A.; et al. Erythropoietin contrastingly affects bacterial infection and experimental colitis by inhibiting nuclear factor-kB-inducible immune pathways. Immunity 2011, 34, 61–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Műzes, G.; Molnár, B.; Tulassay, Z.; Sipos, F. Changes of the cytokine profile in inflammatory bowel diseases. World J. Gastroenterol. 2012, 18, 5848–5861. [Google Scholar] [CrossRef] [Green Version]
- Coquerelle, C.; Oldenhove, G.; Acolty, V.; Denoeud, J.; Vansanten, G.; Verdebout, J.-M.; Mellor, A.; Bluestone, J.A.; Moser, M. Anti-CTLA-4 treatment induces IL-10-producing ICOS+ regulatory T cells displaying IDO-dependent anti-inflammatory properties in a mouse model of colitis. Gut 2009, 58, 1363–1373. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Meng, S.; Jiang, H.; Chen, T.; Wu, W. Exosomes derived from interleukin-10-treated dendritic cells can inhibit trinitrobenzene sulfonic acid-induced rat colitis. Scand. J. Gastroenterol. 2010, 45, 1168–1177. [Google Scholar] [CrossRef]
- Xiong, J.; Lin, Y.-H.; Bi, L.-H.; Wang, J.-D.; Bai, Y.; Liu, S.-D. Effects of Interleukin-4 or Interleukin-10 gene therapy on trinitrobenzenesulfonic acid-induced murine colitis. BMC Gastroenterol. 2013, 13, 165. [Google Scholar] [CrossRef] [Green Version]
- Brenner, H.; Kloor, M.; Pox, C.P. Colorectal Cancer. Lancet 2014, 383, 1490–1502. [Google Scholar] [CrossRef]
- Cooper, H.S.; Murthy, S.; Kido, K.; Yoshitake, H.; Flanigan, A. Dysplasia and cancer in the dextran sulfate sodium mouse colitis model. Relevance to colitis-associated neoplasia in the human: A study of histopathology, B-catenin and p53 expression and the role of inflammation. Carcinogenesis 2000, 21, 757–768. [Google Scholar] [CrossRef] [Green Version]
- Nayak, S.B.; George, B.; Mishra, S. Abnormal Length and Position of the Sigmoid Colon and Its Clinical Significance. Kathmandu Univ. Med. J. 2014, 10, 95–97. [Google Scholar] [CrossRef] [Green Version]
- Marion-Letellier, R.; Bohn, P.; Modzelewski, R.; Vera, P.; Aziz, M.; Guerin, C.; Savoye, G.; Savoye-Collet, C. SPECT-computed tomography in rats with TNBS-induced colitis: A first step toward functional imaging. World J. Gastroenterol. 2017, 23, 216–223. [Google Scholar] [CrossRef] [PubMed]
- Melchior, C.; Loeuillard, E.; Marion-Letellier, R.; Nicol, L.; Mulder, P.; Guerin, C.; Bôle-Feysot, C.; Aziz, M.; Déchelotte, P.; Vera, P.; et al. Magnetic Resonance Colonography for Fibrosis Assessment in Rats with Chronic Colitis. PLoS ONE 2014, 9, e100921. [Google Scholar] [CrossRef] [PubMed]
- Alex, P.; Zachos, N.C.; Nguyen, T.; Gonzales, L.; Chen, T.E.; Conklin, L.S.; Centola, M.; Li, X. Distinct Cytokine Patterns Identified from Multiplex Profiles of Murine DSS and TNBS-Induced Colitis. Inflamm. Bowel Dis. 2009, 15, 341–352. [Google Scholar] [CrossRef]
- Bennink, R.J.; van Montfrans, C.; de Jonge, W.J.; Bruin, K.; van Deventer, S.J.; te Velde, A.A. Imaging of intestinal lymphocyte homing by means of pinhole SPECT in a TNBS colitis mouse model. Nucl. Med. Biol. 2004, 31, 93–101. [Google Scholar] [CrossRef]
- He, Q.; Li, X.; Liu, C.; Su, L.; Xia, Z.; Li, X.; Li, Y.; Li, L.; Yan, T.; Feng, Q.; et al. Dysbiosis of the fecal microbiota in the TNBS-induced Crohn’s disease mouse model. Appl. Microbiol. Biotechnol. 2016, 100, 4485–4494. [Google Scholar] [CrossRef] [PubMed]
- Neurath, M.F. Animal Models of Inflammatory Bowel Diseases: Illuminating the Pathogenesis of Colitis, Ileitis and Cancer. Dig. Dis. 2012, 30, 91–94. [Google Scholar] [CrossRef] [Green Version]
- Pawar, P.; Gilda, S.; Sharma, S.; Jagtap, S.; Paradkar, A.; Mahadik, K.; Ranjekar, P.; Harsulkar, A. Rectal gel application of Withania somnifera root extract expounds anti-inflammatory and muco-restorative activity in TNBS-induced Inflammatory Bowel Disease. BMC Complement. Altern. Med. 2011, 11, 34–39. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
| Procarcinogen Reagent-Related Parameters—AOM | Pro-Inflammatory Reagent-Related Parameters | Rodent-Related Parameters | Model Characterization | Reference | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Number of Administrations | Dose (mg/Kg) | Number of TNBS Administrations | TNBS Dose (mg) | TNBS Volume (µL) | TNBS Vehicle | Number of DSS Administrations (Cycles) | DSS Dose (%) | Strain | Gender | Age (Weeks) | ||
| - | - | - | - | - | - | 3 | 3 | C57BL/6 | M; F | 10 | CSS; M; H | [51] |
| 1 | 10 | 1 | 2.5 | 150 | EtOH | - | - | C57BL/6 | - | 8 | CSS; BM; M | [52] |
| 3–20 | - | - | - | - | 1–4 | 1-3 | C57BL/6; FVB/Ant and IL-6; BALB/c; A/J; FVB/NJ; B6:129; ICR; SAMP; AKR | M; F | 4–12 | CSS; BM; M; H | [53,54,55,56,57,58,59,60,61,62,63,64,65,66,67,68,69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85,86,87,88,89,90,91,92,93,94,95,96,97,98,99,100,101,102,103,104,105,106,107,108,109,110,111,112,113,114,115,116,117,118,119,120,121,122,123,124,125,126,127,128,129,130,131,132,133,134] | |
| 2 | 5–12.5 | - | - | - | - | 2–3 | 0.5–2 | C57BL/6; STAT6; Balb/c | M; F | 5–14 | CSS; BM; M; H | [135,136,137,138,139] |
| 5 | 10 | - | - | - | - | 3 | 1.70 | C57BL/6 | F | 6 | CSS; BM; M | [140] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Modesto, R.; Estarreja, J.; Silva, I.; Rocha, J.; Pinto, R.; Mateus, V. Chemically Induced Colitis-Associated Cancer Models in Rodents for Pharmacological Modulation: A Systematic Review. J. Clin. Med. 2022, 11, 2739. https://doi.org/10.3390/jcm11102739
Modesto R, Estarreja J, Silva I, Rocha J, Pinto R, Mateus V. Chemically Induced Colitis-Associated Cancer Models in Rodents for Pharmacological Modulation: A Systematic Review. Journal of Clinical Medicine. 2022; 11(10):2739. https://doi.org/10.3390/jcm11102739
Chicago/Turabian StyleModesto, Rita, João Estarreja, Inês Silva, João Rocha, Rui Pinto, and Vanessa Mateus. 2022. "Chemically Induced Colitis-Associated Cancer Models in Rodents for Pharmacological Modulation: A Systematic Review" Journal of Clinical Medicine 11, no. 10: 2739. https://doi.org/10.3390/jcm11102739
APA StyleModesto, R., Estarreja, J., Silva, I., Rocha, J., Pinto, R., & Mateus, V. (2022). Chemically Induced Colitis-Associated Cancer Models in Rodents for Pharmacological Modulation: A Systematic Review. Journal of Clinical Medicine, 11(10), 2739. https://doi.org/10.3390/jcm11102739