
Age-Related Alterations of Hyaluronan and Collagen in Extracellular Matrix of the Muscle Spindles

 ,
,  ,
,  ,
,  ,
,  ,
, 
 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. The Ethylenediaminetetraacetic Acid (EDTA) Decalcification Protocol for the Mice Hindlimbs
2.2. Immunohistochemistry Staining: Analysis of Collagen Type I (COLI) and Collagen Type III (COLIII)
2.3. Immunohistochemistry Staining: Analysis of Hyaluronic Acid Binding Protein (HABP)
2.4. Image Analysis
2.5. Statistical Analysis
3. Results
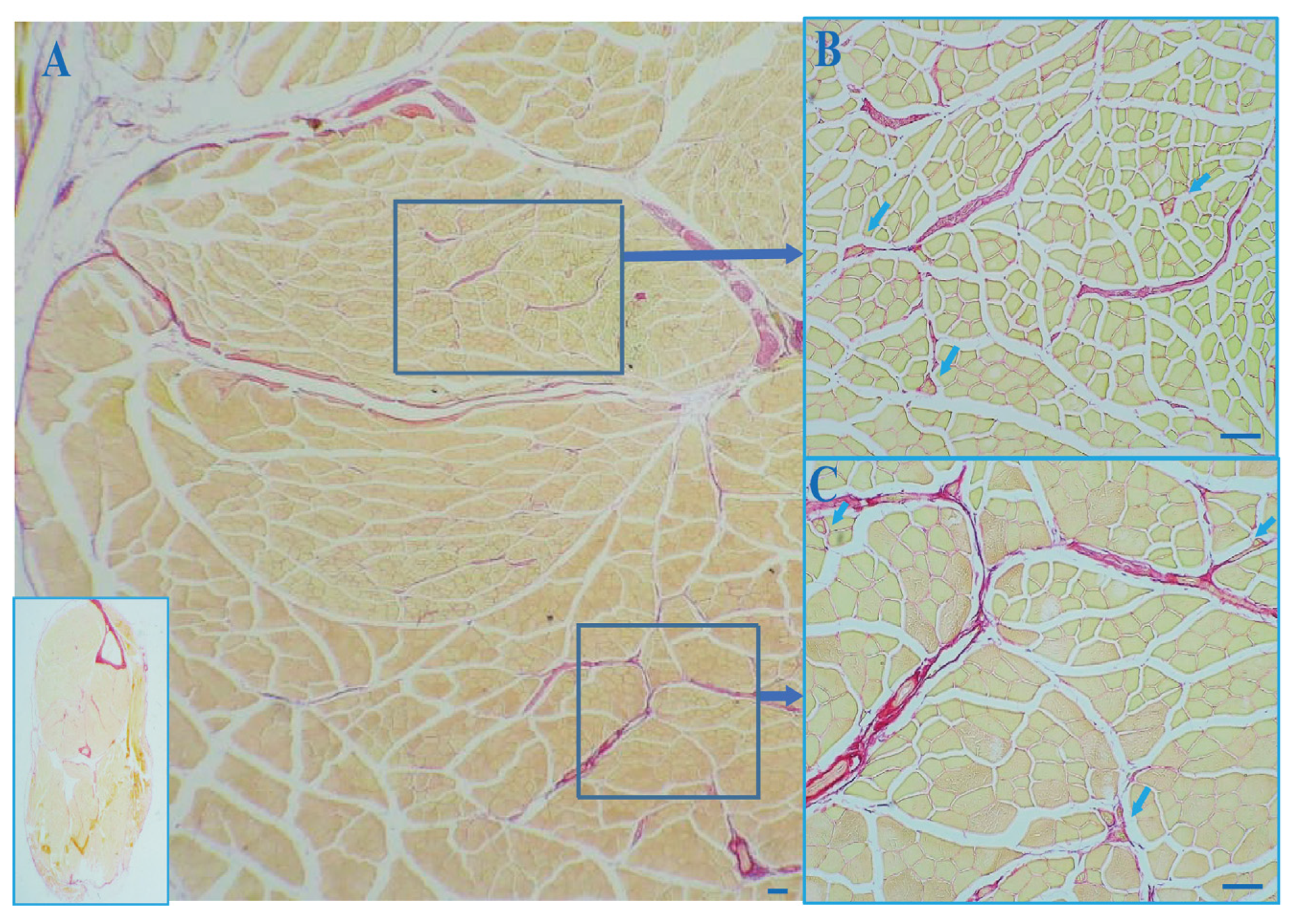
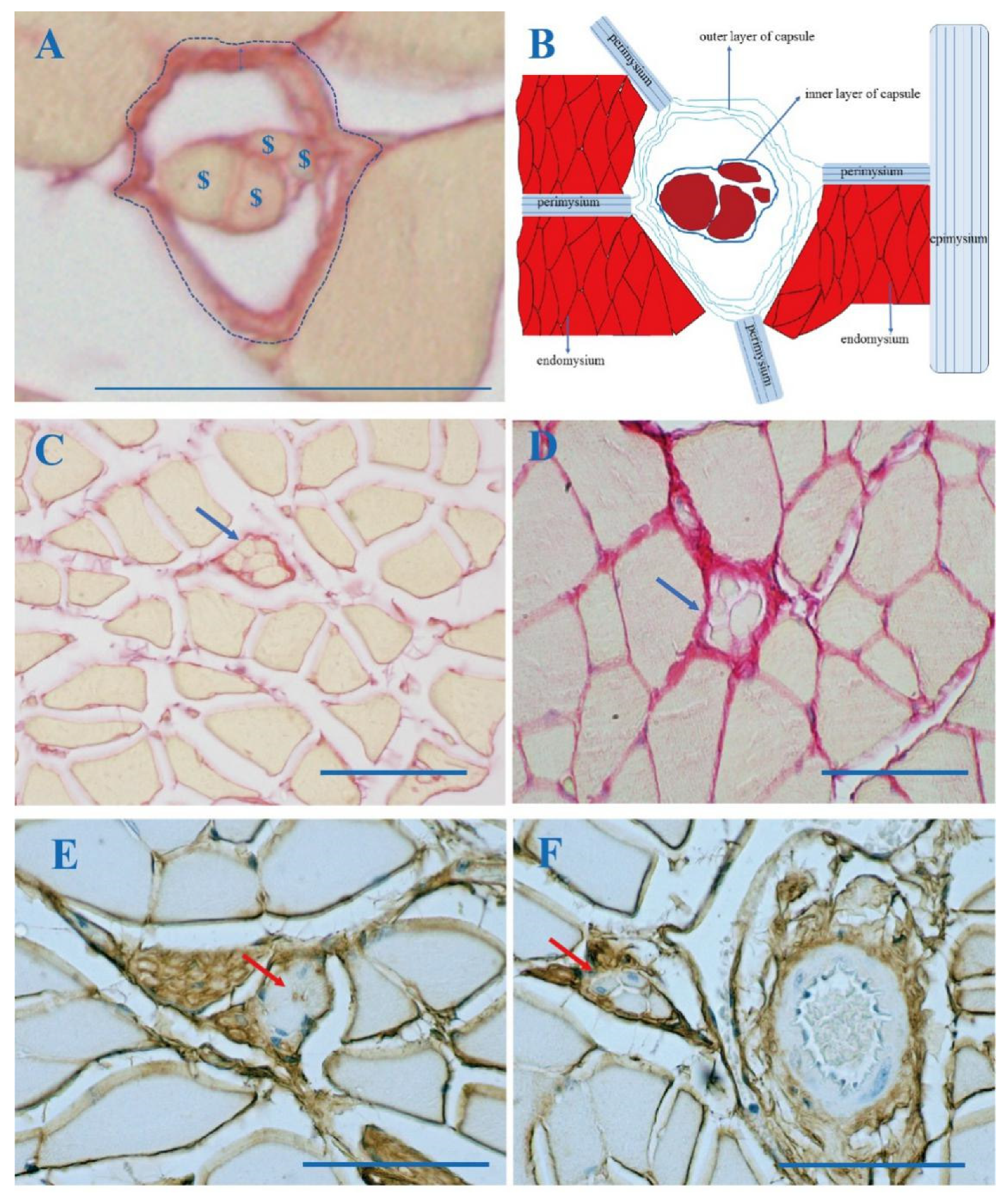
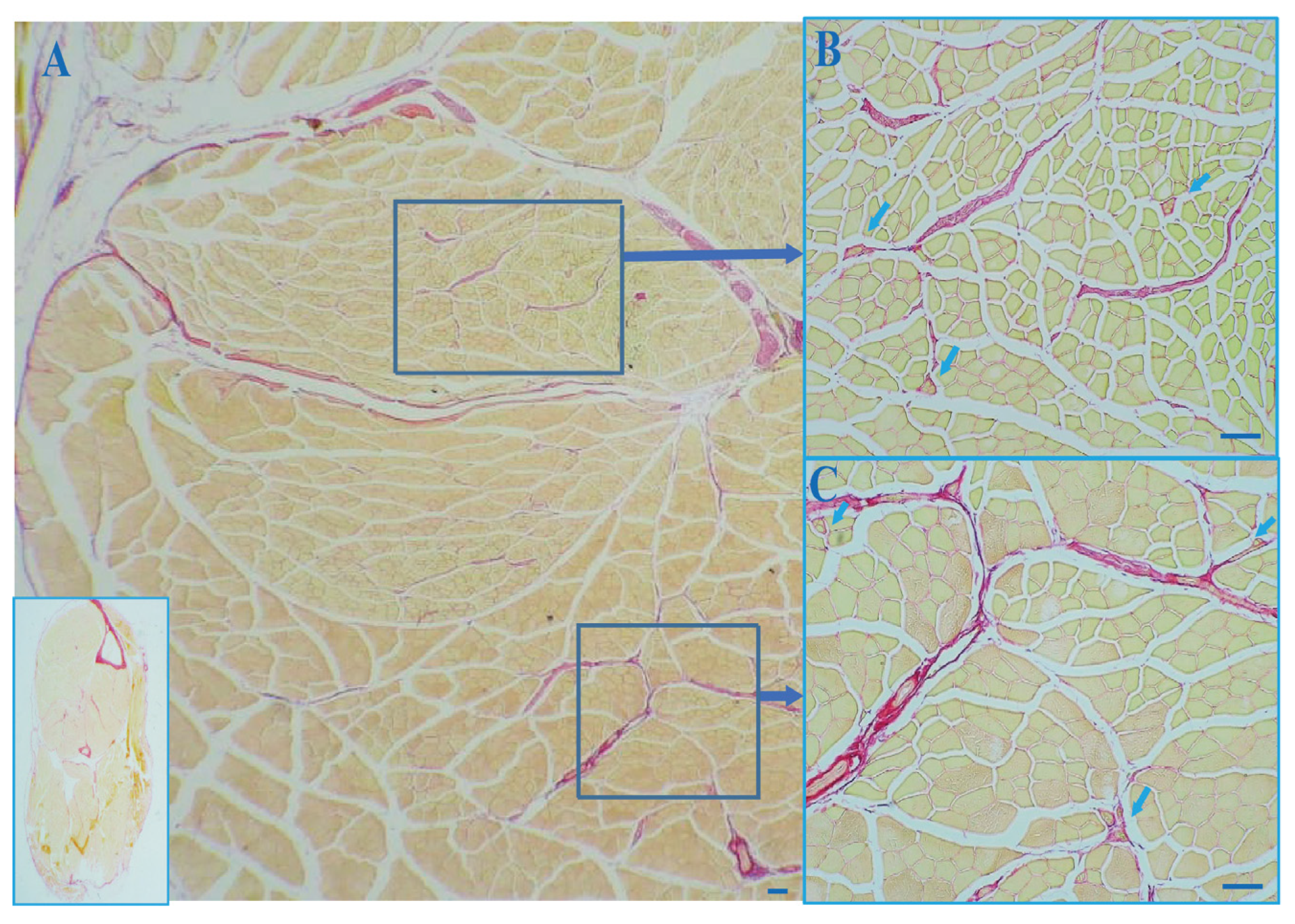
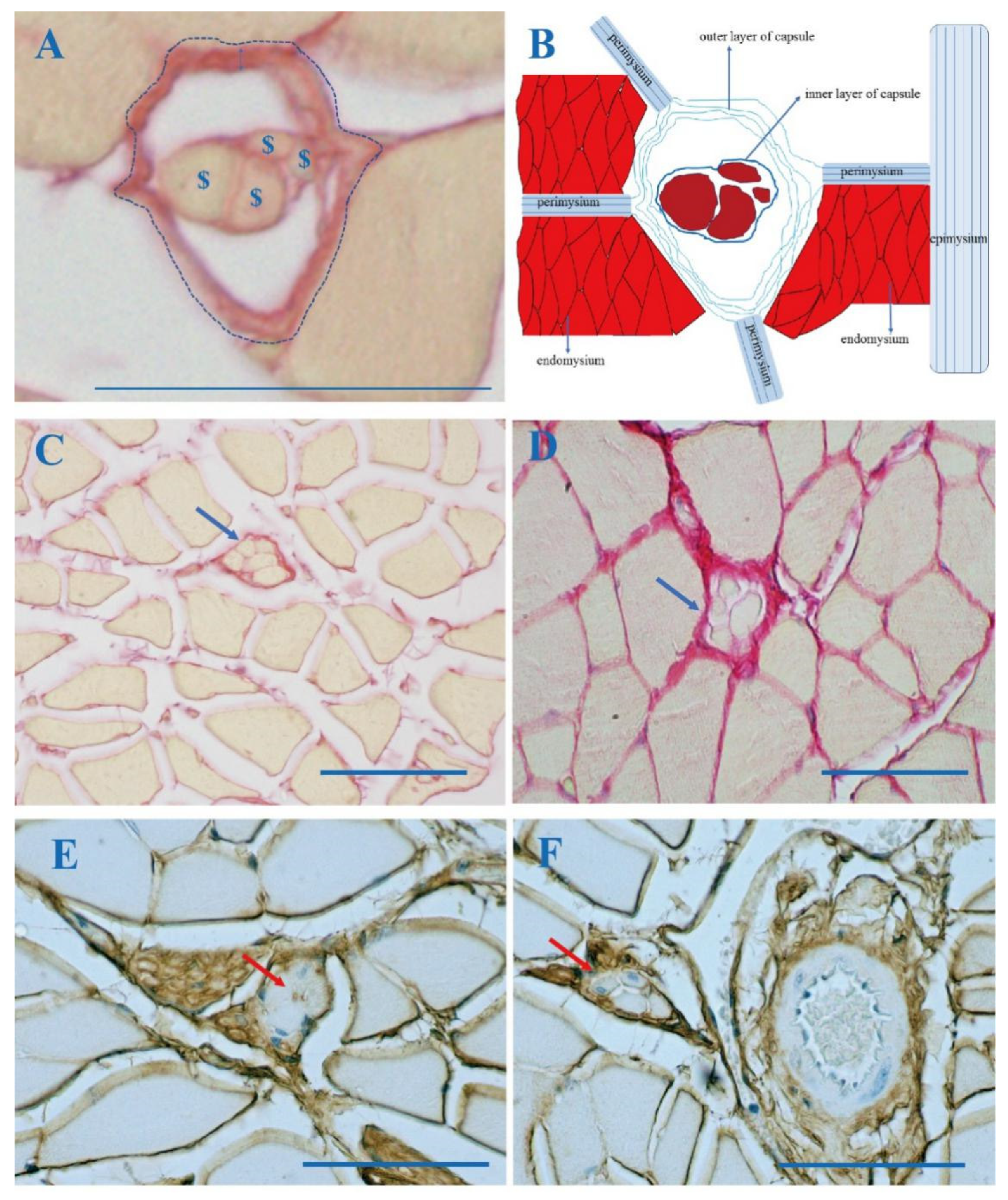
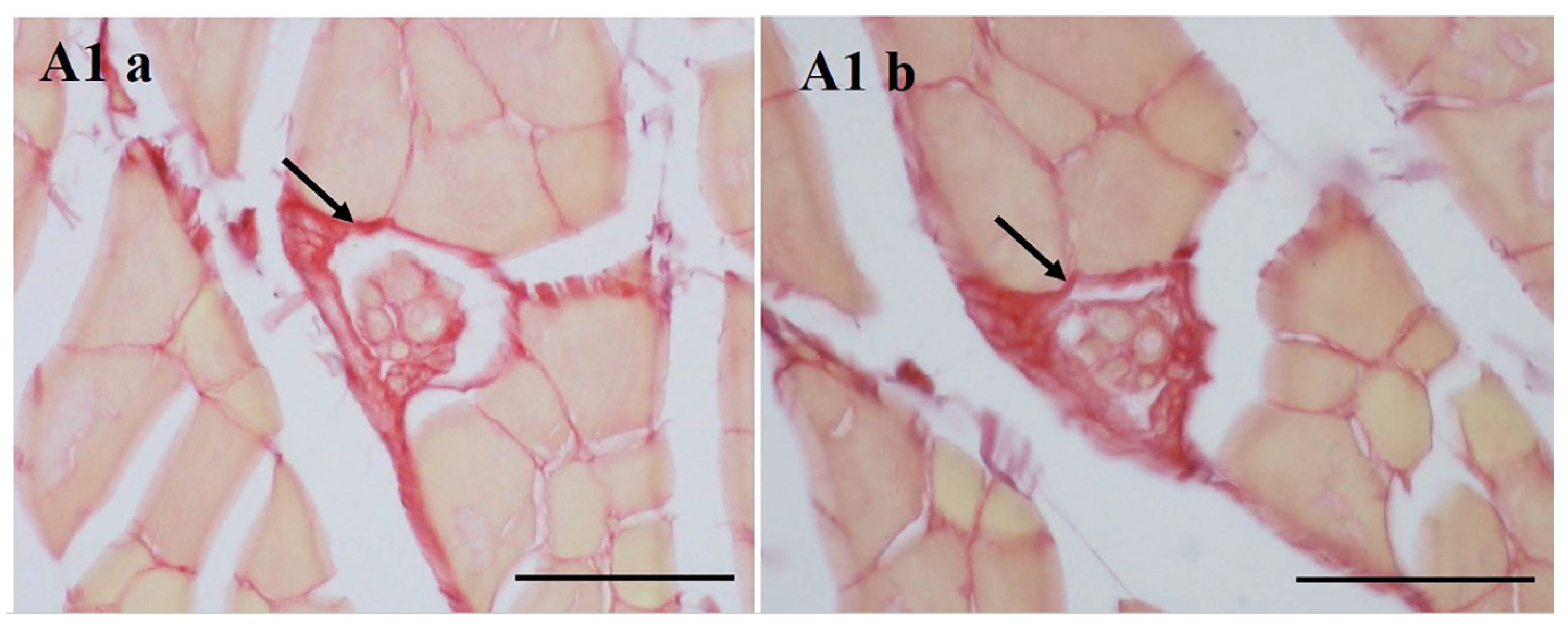
3.1. The Outer Capsule of MS Is Continuous with Intramuscular Connective Tissue (IMCT)
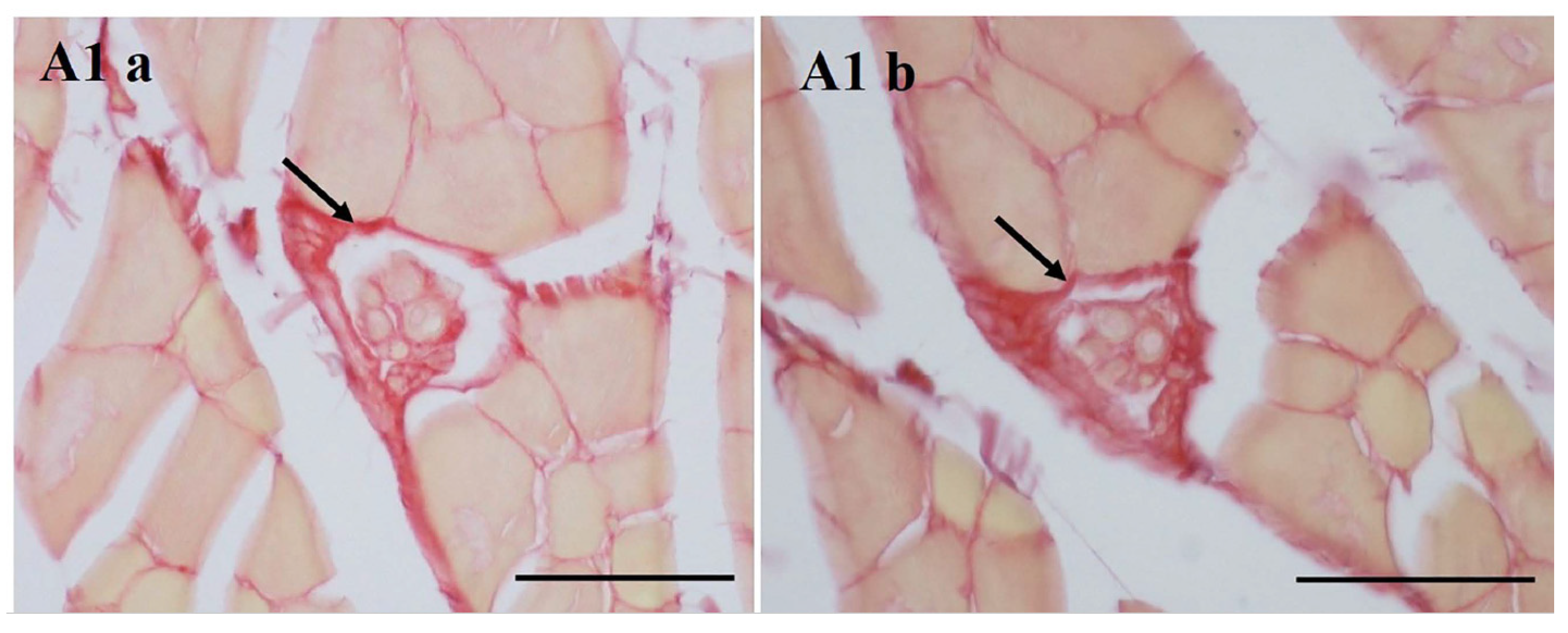
3.2. Collagen in the MS with Aging
3.3. COLI and COLIII in MS
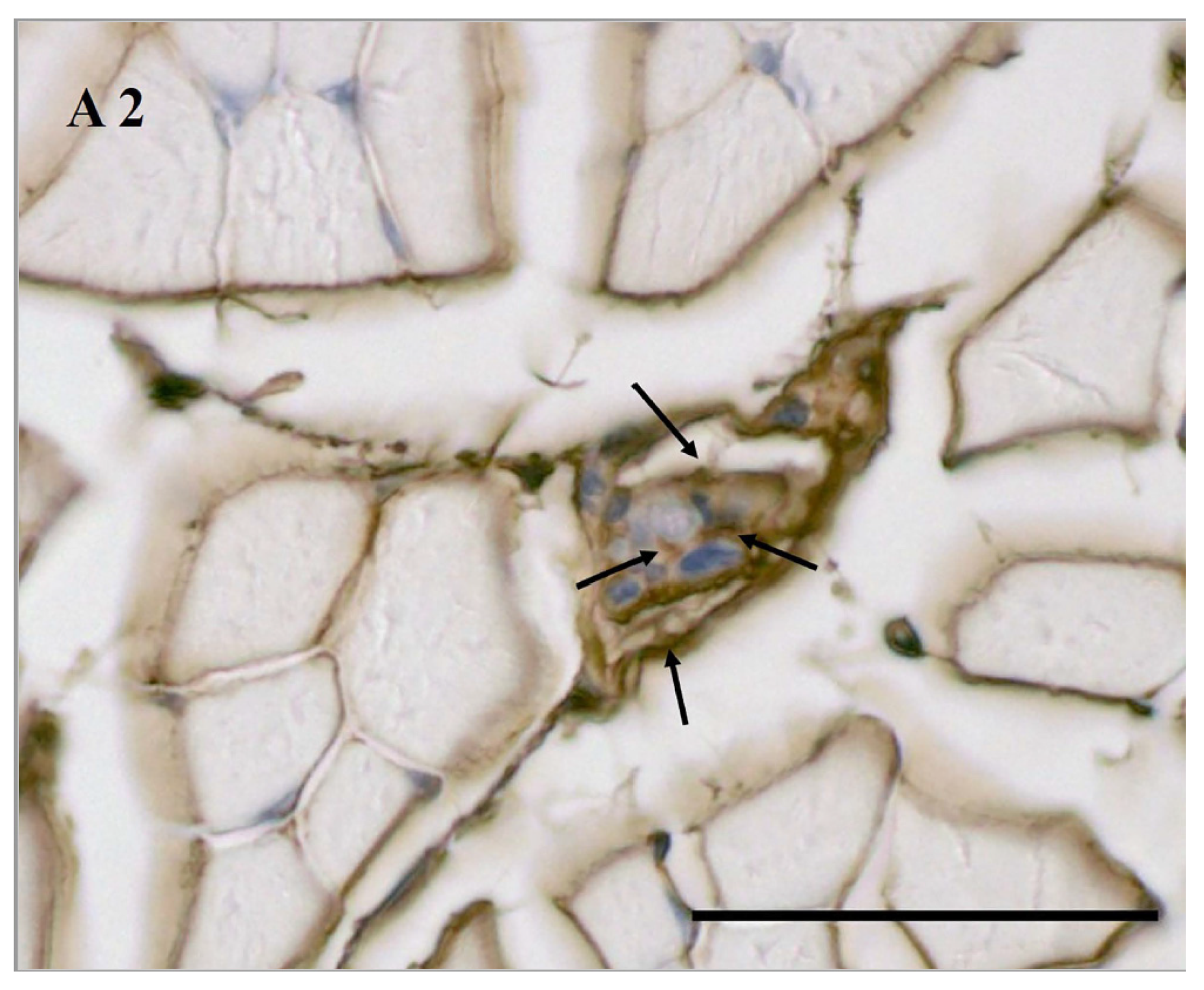
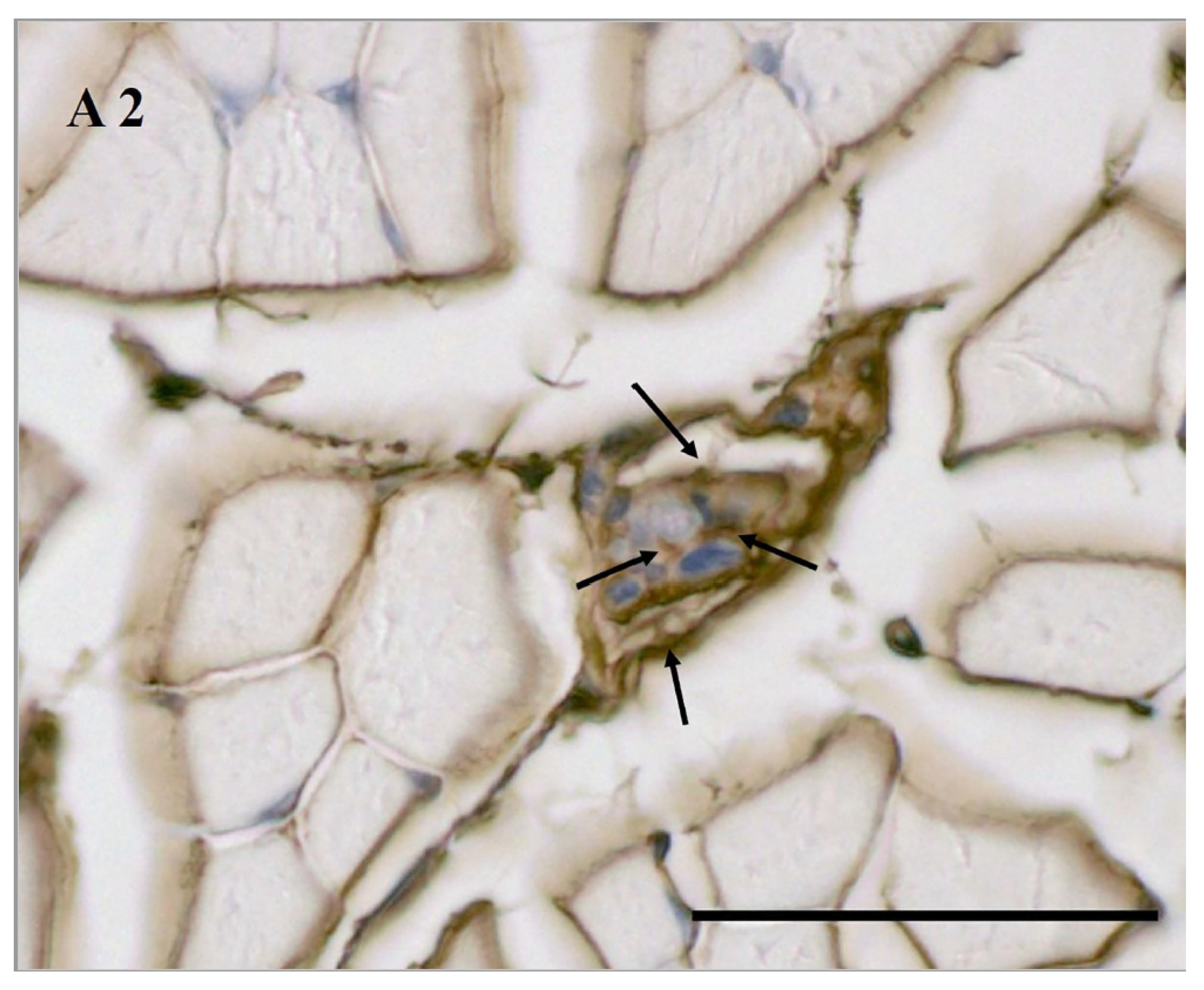
3.4. Age-Related HA in MS According to Biotinylated HABP Immunohistochemical Staining
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
Appendix A
References
- Banks, R.W.; Ellaway, P.H.; Prochazka, A.; Proske, U. Secondary endings of muscle spindles: Structure, reflex action, role in motor control and proprioception. Exp. Physiol. 2021, 106, 2339–2366. [Google Scholar] [CrossRef]
- Windhorst, U. Muscle proprioceptive feedback and spinal networks. Brain Res. Bull. 2007, 73, 155–202. [Google Scholar] [CrossRef] [PubMed]
- Miwa, T.; Miwa, Y.; Kanda, K. Dynamic and static sensitivities of muscle spindle primary endings in aged rats to ramp stretch. Neurosci. Lett. 1995, 201, 179–182. [Google Scholar] [CrossRef]
- De Reuck, J. Biometric analyses of spindles in normal human skeletal muscles. Acta Neurol. Belg. 1973, 73, 339–347. [Google Scholar] [PubMed]
- Boyd-Clark, L.; Briggs, C.; Galea, M. Muscle spindle distribution, morphology, and density in longus colli and multifidus muscles of the cervical spine. Spine 2002, 27, 694–701. [Google Scholar] [CrossRef] [Green Version]
- Boyd-Clark, L.; Galea, M.; Briggs, C.; Opeskin, K. Monitoring age-related changes of collagen content and vascularity in ganglia using unbiased stereological methods. J. Microsc. 2000, 200, 284–290. [Google Scholar] [CrossRef]
- Roberts, N.; Cruz-Orive, L.; Reid, N.; Brodie, D.; Bourne, M.; Edwards, R. Unbiased estimation of human body composition by the Cavalieri method using magnetic resonance imaging. J. Microsc. 1993, 171, 239–253. [Google Scholar] [CrossRef] [PubMed]
- Roberts, N.; Garden, A.; Cruz-Orive, L.; Whitehouse, G.; Edwards, R. Estimation of fetal volume by magnetic resonance imaging and stereology. Br. J. Radiol. 1994, 67, 1067–1077. [Google Scholar] [CrossRef] [PubMed]
- Swash, M.; Fox, K.P. The effect of age on human skeletal muscle studies of the morphology and innervation of muscle spindles. J. Neurol. Sci. 1972, 16, 417–432. [Google Scholar] [CrossRef]
- Kim, G.H.; Suzuki, S.; Kanda, K. Age-related physiological and morphological changes of muscle spindles in rats. J. Physiol. 2007, 582, 525–538. [Google Scholar] [CrossRef]
- Goble, D.J.; Coxon, J.P.; Wenderoth, N.; Van Impe, A.; Swinnen, S.P. Proprioceptive sensibility in the elderly: Degeneration, functional consequences and plastic-adaptive processes. Neurosci. Biobehav. Rev. 2009, 33, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, S.K.; Stanley, O.L.; Valdez, G. Impact of aging on proprioceptive sensory neurons and intrafusal muscle fibers in mice. J. Gerontol. A Biomed. Sci. Med. Sci. 2017, 72, 771–779. [Google Scholar] [CrossRef]
- Rosant, C.; Nagel, M.-D.; Pérot, C. Aging affects passive stiffness and spindle function of the rat soleus muscle. Exp. Gerontol. 2007, 42, 301–308. [Google Scholar] [CrossRef]
- Desaki, J.; Nishida, N. A further observation of muscle spindles in the extensor digitorum longus muscle of the aged rat. J. Electron Microsc. (Tokyo) 2010, 59, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Henry, M.; Baudry, S.J. Age-related changes in leg proprioception: Implications for postural control. J. Neurophysiol. 2019, 122, 525–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavan, P.; Monti, E.; Bondí, M.; Fan, C.; Stecco, C.; Narici, M.; Reggiani, C.; Marcucci, L. Alterations of Extracellular Matrix Mechanical Properties Contribute to Age-Related Functional Impairment of Human Skeletal Muscles. Int. J. Mol. Sci. 2020, 21, 3992. [Google Scholar] [CrossRef]
- Birch, H.L. Extracellular Matrix and Ageing. Subcell. Biochem. 2018, 90, 169–190. [Google Scholar]
- Kadler, K.E.; Baldock, C.; Bella, J.; Boot-Handford, R.P. Collagens at a glance. J. Cell Sci. 2007, 120, 1955–1958. [Google Scholar] [CrossRef] [Green Version]
- Poole, J.J.A.; Mostaço-Guidolin, L.B. Optical Microscopy and the Extracellular Matrix Structure: A Review. Cells 2021, 10, 1760. [Google Scholar] [CrossRef] [PubMed]
- Soundararajan, A.; Ghag, S.A.; Vuda, S.S.; Wang, T.; Pattabiraman, P.P. Cathepsin K Regulates Intraocular Pressure by Modulating Extracellular Matrix Remodeling and Actin-Bundling in the Trabecular Meshwork Outflow Pathway. Cells 2021, 10, 2864. [Google Scholar] [CrossRef]
- Parkes, W.S.; Amargant, F.; Zhou, L.T.; Villanueva, C.E.; Duncan, F.E.; Pritchard, M.T. Hyaluronan and Collagen Are Prominent Extracellular Matrix Components in Bovine and Porcine Ovaries. Genes 2021, 12, 1186. [Google Scholar] [CrossRef]
- Burr, S.D.; Stewart, J.A., Jr. Rap1a Regulates Cardiac Fibroblast Contraction of 3D Diabetic Collagen Matrices by Increased Activation of the AGE/RAGE Cascade. Cells 2021, 10, 1286. [Google Scholar] [CrossRef]
- Dalewski, B.; Białkowska, K.; Pałka, Ł.; Jakubowska, A.; Kiczmer, P.; Sobolewska, E. COL5A1 RS12722 Is Associated with Temporomandibular Joint Anterior Disc Displacement without Reduction in Polish Caucasians. Cells 2021, 10, 2423. [Google Scholar] [CrossRef]
- Pawlina, W.; Ross, M.H. Connective tissue. In Histology: A Text and Atlas: With Correlated Cell and Molecular Biology; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2018; pp. 156–192. [Google Scholar]
- Järvinen, T.A.; Józsa, L.; Kannus, P.; Järvinen, T.L.; Järvinen, M. Organization and distribution of intramuscular connective tissue in normal and immobilized skeletal muscles. J. Muscle Res. Cell Motil. 2002, 23, 245–254. [Google Scholar] [CrossRef]
- Stecco, A.; Stecco, C.; Raghavan, P. Peripheral mechanisms contributing to spasticity and implications for treatment. Curr. Phys. Med. Rehabil. Rep. 2014, 2, 121–127. [Google Scholar] [CrossRef] [Green Version]
- Dieler, R.; Schröder, J.M. Increase of elastic fibres in muscle spindles of rats following single or repeated denervation with or without reinnervation. Virchows Arch. A Pathol. Anat. Histopathol. 1990, 417, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Purslow, P.P. The structure and role of intramuscular connective tissue in muscle function. Front. Physiol. 2020, 11, 495. [Google Scholar] [CrossRef]
- Koike, S.; Mukudai, S.; Hisa, Y. Muscle Spindles and Intramuscular Ganglia. In Neuroanatomy and Neurophysiology of the Larynx; Springer: Berlin/Heidelberg, Germany, 2016; pp. 11–20. [Google Scholar]
- Ovalle, W.; Dow, P. Morphological Aspects of the Muscle Spindle Capsule and Its Functional Significance. In The Muscle Spindle; Springer: Berlin/Heidelberg, Germany, 1985; pp. 23–28. [Google Scholar]
- Dutta, S.; Sengupta, P. Men and mice: Relating their ages. Life Sci. 2016, 152, 244–248. [Google Scholar] [CrossRef]
- Banks, R. The motor innervation of mammalian muscle spindles. Prog. Neurobiol. 1994, 43, 323–362. [Google Scholar] [CrossRef]
- Bewick, G.S.; Banks, R.W. Mechanotransduction in the muscle spindle. Pflügers Arch. Eur. J. Physiol. 2015, 467, 175–190. [Google Scholar] [CrossRef] [Green Version]
- Crowe, A.R.; Yue, W. Semi-quantitative determination of protein expression using immunohistochemistry staining and analysis: An integrated protocol. Bio-Protocol 2019, 9, e3465. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Maier, A. Extracellular matrix and transmembrane linkages at the termination of intrafusal fibers and the outer capsule in chicken muscle spindles. J. Morphol. 1996, 228, 335–346. [Google Scholar] [CrossRef]
- Ovalle, W.K.; Dow, P.R.; Nahirney, P.C. Structure, distribution and innervation of muscle spindles in avian fast and slow skeletal muscle. J. Anat. 1999, 194, 381–394. [Google Scholar] [CrossRef] [PubMed]
- Eldred, E.; Maier, A.; Bridgman, C.F. Differences in intrafusal fiber content of spindles in several muscles of the cat. Exp. Neurol. 1974, 45, 8–18. [Google Scholar] [CrossRef]
- Desaki, J.; Uehara, Y. A fine-structural study of the termination of intrafusal muscle fibres in the Chinese hamster. Cell Tissue Res. 1983, 234, 723–733. [Google Scholar] [CrossRef] [PubMed]
- Sahgal, V.; Subramani, V.; Sahgal, S. Ultrastructure of Attachments of Human Intrafusal Fibers. In Mechanoreceptors; Springer: Berlin/Heidelberg, Germany, 1988; pp. 247–253. [Google Scholar]
- Mokone, G.G.; Schwellnus, M.P.; Noakes, T.D.; Collins, M. The COL5A1 gene and Achilles tendon pathology. Scand. J. Med. Sci. Sports 2006, 16, 19–26. [Google Scholar] [CrossRef]
- September, A.V.; Cook, J.; Handley, C.J.; Van Der Merwe, L.; Schwellnus, M.P.; Collins, M. Variants within the COL5A1 gene are associated with Achilles tendinopathy in two populations. Br. J. Sports Med. 2008, 43, 357–365. [Google Scholar] [CrossRef]
- Posthumus, M.; September, A.V.; O’Cuinneagain, D.; Van Der Merwe, W.; Schwellnus, M.P.; Collins, M. The COL5A1 gene is associated with increased risk of anterior cruciate ligament ruptures in female participants. Am. J. Sports Med. 2009, 37, 2234–2240. [Google Scholar] [CrossRef]
- O’Connell, K.; Knight, H.; Ficek, K.; Leonska-Duniec, A.; Maciejewska-Karlowska, A.; Sawczuk, M.; Stepien-Slodkowska, M.; O’Cuinneagain, D.; van der Merwe, W.; Posthumus, M.; et al. Interactions between collagen gene variants and risk of anterior cruciate ligament rupture. Eur. J. Sport Sci. 2015, 15, 341–350. [Google Scholar] [CrossRef]
- Altinisik, J.; Meric, G.; Erduran, M.; Ates, O.; Ulusal, A.E.; Akseki, D. The BstUI and DpnII variants of the COL5A1 gene are associated with tennis elbow. Am. J. Sports Med. 2015, 43, 1784–1789. [Google Scholar] [CrossRef]
- Pirri, C.; Fede, C.; Pirri, N.; Petrelli, L.; Fan, C.; De Caro, R.; Stecco, C. Diabetic Foot: The Role of Fasciae, a Narrative Review. Biology 2021, 10, 759. [Google Scholar] [CrossRef]
- Correa-Gallegos, D.; Jiang, D.; Christ, S.; Ramesh, P.; Ye, H.; Wannemacher, J.; Gopal, S.K.; Yu, Q.; Aichler, M.; Walch, A.; et al. Patch repair of deep wounds by mobilized fascia. Nat. Cell Biol. 2019, 576, 287–292. [Google Scholar] [CrossRef]
- Kang, L.; Ayala, J.E.; Lee-Young, R.S.; Zhang, Z.; James, F.D.; Neufer, P.D.; Pozzi, A.; Zutter, M.M.; Wasserman, D.H. Di-etinduced muscle insulin resistance is associated with extracellular matrix remodeling and interaction with integrin al-pha2beta1 in mice. Diabetes 2011, 60, 416–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stecco, C. Functional Atlas of the Human Fascial System E-Book; Elsevier Health Sciences: Amsterdam, The Netherlands, 2014. [Google Scholar]
- Fukami, Y. Studies of capsule and capsular space of cat muscle spindles. J. Physiol. 1986, 376, 281–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pratt, R.L. Hyaluronan and the Fascial Frontier. Int. J. Mol. Sci. 2021, 22, 6845. [Google Scholar] [CrossRef]
- De Luca, C.; Kline, J. Influence of proprioceptive feedback on the firing rate and recruitment of motoneurons. J. Neural Eng. 2012, 9, 016007. [Google Scholar] [CrossRef]
- Stecco, L. Atlas of Physiology of the Muscular Fascia; Piccin: Padova, Italy, 2016. [Google Scholar]
- Blecher, R.; Krief, S.; Galili, T.; Biton, I.E.; Stern, T.; Assaraf, E.; Levanon, D.; Appel, E.; Anekstein, Y.; Agar, G. The propri-oceptive system masterminds spinal alignment: Insight into the mechanism of scoliosis. Dev. Cell 2017, 42, 388–399. [Google Scholar] [CrossRef]
- Abedi Khoozani, P.; Blohm, G. Neck muscle spindle noise biases reaches in a multisensory integration task. J. Neurophysiol. 2018, 120, 893–909. [Google Scholar] [CrossRef]
- Langevin, H.M.; Rizzo, D.M.; Fox, J.R.; Badger, G.J.; Junru, W.; Konofagou, E.E.; Stevens-Tuttle, D.; Bouffard, N.A.; Krag, M.H. Dynamic morphometric characterization of local connective tissue network structure in humans using ultrasound. BMC Syst. Biol. 2007, 5, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pirri, C.; Fede, C.; Petrelli, L.; Guidolin, D.; Fan, C.; De Caro, R.; Stecco, C. An anatomical comparison of the fasciae of the thigh: A macroscopic, microscopic and ultrasound imaging study. J. Anat. 2021, 238, 999–1009. [Google Scholar] [CrossRef] [PubMed]
- Kerns, M.J.J.; Darst, M.A.; Olsen, T.G.; Fenster, M.; Hall, P.; Grevey, S. Shrinkage of cutaneous specimens: Formalin or other factors involved? J. Cutan. Pathol. 2008, 35, 1093–1096. [Google Scholar] [CrossRef] [PubMed]
- Leonard, C.T. The Neuroscience of Human Movement; Mosby Incorporated: St. Louis, MO, USA, 1998. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Group A | Group B | Group C | A vs. B p-Value | A vs. C p-Value | B vs. C p-Value |
|---|---|---|---|---|---|---|
| $ Thickness of the outer capsule layer | 3.02 ± 0.26 | 3.64 ± 0.31 | 5.81 ± 0.85 | 0.022 * | 0.003 ** | 0.007 ** |
| $ Total collagen 40× (% area) | 2.95 ± 0.46 | 4.42 ± 1.23 | 9.29 ± 0.81 | 0.116 | <0.001 *** | <0.001 *** |
| $ Collagen of the MS alone 40× (% area) | 22.97 ± 6.55 | 25.94 ± 2.36 | 40.80 ± 3.46 | 0.633 | 0.004 ** | <0.001 *** |
| # AOD of COLI in the whole muscle cross section | 0.20 ± 0.02 | 0.27 ± 0.02 | 0.30 ± 0.01 | <0.001 *** | <0.001 *** | 0.032 * |
| # AOD of COLIII in the whole muscle cross section | 0.25 ± 0.02 | 0.28 ± 0.01 | 0.26 ± 0.02 | 0.255 | 0.747 | 0.629 |
| # AOD of COLI in the MS alone | 0.16 ± 0.01 | 0.22 ± 0.01 | 0.28 ± 0.06 | 0.038 * | <0.001 *** | 0.047 * |
| # AOD of COLIII in the MS alone | 0.25 ± 0.07 | 0.27 ± 0.07 | 0.26 ± 0.05 | 0.849 | 0.968 | 0.950 |
| & AOD of HABP in the MS alone | 0.50 (0.45–0.67) | 0.43 (0.40–0.45) | 0.40 (0.33–0.47) | 0.85 | 0.022 * | 1.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, C.; Pirri, C.; Fede, C.; Guidolin, D.; Biz, C.; Petrelli, L.; Porzionato, A.; Macchi, V.; De Caro, R.; Stecco, C. Age-Related Alterations of Hyaluronan and Collagen in Extracellular Matrix of the Muscle Spindles. J. Clin. Med. 2022, 11, 86. https://doi.org/10.3390/jcm11010086
Fan C, Pirri C, Fede C, Guidolin D, Biz C, Petrelli L, Porzionato A, Macchi V, De Caro R, Stecco C. Age-Related Alterations of Hyaluronan and Collagen in Extracellular Matrix of the Muscle Spindles. Journal of Clinical Medicine. 2022; 11(1):86. https://doi.org/10.3390/jcm11010086
Chicago/Turabian StyleFan, Chenglei, Carmelo Pirri, Caterina Fede, Diego Guidolin, Carlo Biz, Lucia Petrelli, Andrea Porzionato, Veronica Macchi, Raffaele De Caro, and Carla Stecco. 2022. "Age-Related Alterations of Hyaluronan and Collagen in Extracellular Matrix of the Muscle Spindles" Journal of Clinical Medicine 11, no. 1: 86. https://doi.org/10.3390/jcm11010086
APA StyleFan, C., Pirri, C., Fede, C., Guidolin, D., Biz, C., Petrelli, L., Porzionato, A., Macchi, V., De Caro, R., & Stecco, C. (2022). Age-Related Alterations of Hyaluronan and Collagen in Extracellular Matrix of the Muscle Spindles. Journal of Clinical Medicine, 11(1), 86. https://doi.org/10.3390/jcm11010086