
Investigating the Relationship between White Matter Connectivity and Motivational Circuits in Subjects with Deficit Schizophrenia: A Diffusion Tensor Imaging (DTI) Study


 ,
,  ,
,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects
- diagnosis of schizophrenia based on the criteria of the DSM-IV, confirmed by the Mini International Neuropsychiatric Interview Plus (MINI-Plus);
- age between 18 and 65 years;
- negative history of intellectual disability, head trauma with unconsciousness, alcohol or substance abuse within the previous six months (except for cigarette smoking);
- no treatment modifications and/or hospitalization due to symptom exacerbation in the last three months;
- treatment with second-generation antipsychotics [82].
- presence of current or lifetime Axis I or II psychiatric diagnosis; history of psychiatric hospitalization;
- history of head trauma with unconsciousness;
- history of substance abuse or dependence (except for cigarette smoking) and use of drugs that affect the central nervous system.
2.2. Assessment Instruments
2.3. MRI Acquisition and Parameters
2.4. Region of Interest
- Left and right DLPFC were defined combining on each side the Brodmann areas 9 and 46 [89], as defined in the WFUPick-Atlas.
- Orbito-frontal cortices were preliminarily obtained by combining the Brodmann areas 10 and 11, as defined in the WFUPick-Atlas, and were then divided on each side of the brain in their medial (mOFC) and lateral (lOFC) parts using the sagittal planes placed 20 mm off-center as separators [90].
- For each side, vaIC, daIC and pIC ROIs were obtained by dividing the entire available ROIs of insular cortex in the Harvard-Oxford Cortical Structural Atlas [91], based on its connectivity [92]. DTI pre-processing and probabilistic tractography were performed using the software modules provided in the FMRIB Software Library (FSL, http://fsl.fmrib.ox.ac.uk/fsl, accessed on 15 July 2017).
2.5. Probabilistic Tractography
2.6. Statistical Analysis
3. Results
3.1. Subject Characteristics
3.2. Group Comparison on the Connectivity Index and Fractional Anisotropy between Couples of ROIs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kraepelin, E. Dementia Praecox and Paraphrenia; Chicago Medical Book: Chicago, IL, USA, 1919. [Google Scholar]
- Bleuler, E. Dementia Praecox or the Group of Schizophrenias; International Universities Press: New York, NY, USA, 1950. [Google Scholar]
- Foussias, G.; Agid, O.; Fervaha, G.; Remington, G. Negative symptoms of schizophrenia: Clinical features, relevance to real world functioning and specificity versus other CNS disorders. Eur. Neuropsychopharmacol. 2014, 24, 693–709. [Google Scholar] [CrossRef]
- Galderisi, S.; Mucci, A.; Buchanan, R.W.; Arango, C. Negative symptoms of schizophrenia: New developments and unanswered research questions. Lancet Psychiatry 2018, 5, 664–677. [Google Scholar] [CrossRef]
- Sahakian, B.; Savulich, G. Innovative methods for improving cognition, motivation and wellbeing in schizophrenia. World Psychiatry 2019, 18, 168–170. [Google Scholar] [CrossRef]
- Falkai, P.; Schmitt, A. The need to develop personalized interventions to improve cognition in schizophrenia. World Psychiatry 2019, 18, 170. [Google Scholar] [CrossRef]
- Harvey, P.D.; Strassnig, M.T. Cognition and disability in schizophrenia: Cognition-related skills deficits and decision-making challenges add to morbidity. World Psychiatry 2019, 18, 165–167. [Google Scholar] [CrossRef]
- Keefe, R.S. Why are there no approved treatments for cognitive impairment in schizophrenia? World Psychiatry 2019, 18, 167–168. [Google Scholar] [CrossRef] [PubMed]
- Reichenberg, A.; Velthorst, E.; Davidson, M. Cognitive impairment and psychosis in schizophrenia: Independent or linked conditions? World Psychiatry 2019, 18, 162–163. [Google Scholar] [CrossRef] [PubMed]
- Correll, C.U.; Schooler, N.R. Negative Symptoms in Schizophrenia: A Review and Clinical Guide for Recognition, Assessment, and Treatment. Neuropsychiatr. Dis. Treat. 2020, 16, 519–534. [Google Scholar] [CrossRef] [PubMed]
- Galderisi, S.; Rucci, P.; Mucci, A.; Rossi, A.; Rocca, P.; Bertolino, A.; Aguglia, E.; Amore, M.; Bellomo, A.; Bozzatello, P.; et al. The interplay among psychopathology, personal resources, context-related factors and real-life functioning in schizophrenia: Stability in relationships after 4 years and differences in network structure between recovered and non-recovered patients. World Psychiatry 2020, 19, 81–91. [Google Scholar] [CrossRef]
- Heckers, S.; Kendler, K.S. The evolution of Kraepelin’s nosological principles. World Psychiatry 2020, 19, 381–388. [Google Scholar] [CrossRef]
- Galderisi, S.; Kaiser, S.; Bitter, I.; Nordentoft, M.; Mucci, A.; Sabé, M.; Giordano, G.M.; Nielsen, M.; Glenthøj, L.B.; Pezzella, P.; et al. EPA guidance on treatment of negative symptoms in schizophrenia. Eur. Psychiatry 2021, 64, e21. [Google Scholar] [CrossRef]
- Galderisi, S.; Mucci, A.; Dollfus, S.; Nordentoft, M.; Falkai, P.; Kaiser, S.; Giordano, G.M.; Vandevelde, A.; Nielsen, M.; Glenthøj, L.B.; et al. EPA guidance on assessment of negative symptoms in schizophrenia. Eur. Psychiatry 2021, 64, 1–91. [Google Scholar] [CrossRef]
- Kirkpatrick, B.; Fenton, W.S.; Carpenter, W.T.; Marder, S.R. The NIMH-MATRICS Consensus Statement on Negative Symptoms. Schizophr. Bull. 2006, 32, 214–219. [Google Scholar] [CrossRef]
- Marder, S.R.; Galderisi, S. The current conceptualization of negative symptoms in schizophrenia. World Psychiatry 2017, 16, 14–24. [Google Scholar] [CrossRef]
- Reed, G.M.; First, M.B.; Kogan, C.S.; Hyman, S.E.; Gureje, O.; Gaebel, W.; Maj, M.; Stein, D.; Maercker, A.; Tyrer, P.; et al. Innovations and changes in the ICD-11 classification of mental, behavioural and neurodevelopmental disorders. World Psychiatry 2019, 18, 3–19. [Google Scholar] [CrossRef] [PubMed]
- Gaebel, W.; Falkai, P.; Hasan, A. The revised German evidence- and consensus-based schizophrenia guideline. World Psychiatry 2020, 19, 117–119. [Google Scholar] [CrossRef]
- Giordano, G.M.; Koenig, T.; Mucci, A.; Vignapiano, A.; Amodio, A.; di Lorenzo, G.; Siracusano, A.; Bellomo, A.; Altamura, M.; Monteleone, P.; et al. Neurophysiological correlates of Avolition-apathy in schizophrenia: A resting-EEG microstates study. NeuroImage Clin. 2018, 20, 627–636. [Google Scholar] [CrossRef] [PubMed]
- Giordano, G.M.; Brando, F.; Perrottelli, A.; Di Lorenzo, G.; Siracusano, A.; Giuliani, L.; Pezzella, P.; Altamura, M.; Bellomo, A.; Cascino, G.; et al. Tracing links between early auditory information processing and negative symptoms in schizophrenia: An ERP study. Front. Psychiatry, 2021; in press. [Google Scholar]
- Galderisi, S.; Maj, M.; Mucci, A.; Cassano, G.B.; Invernizzi, G.; Rossi, A.; Vita, A.; Dell’Osso, L.; Daneluzzo, E.; Pini, S. Historical, Psychopathological, Neurological, and Neuropsychological Aspects of Deficit Schizophrenia: A Multicenter Study. Am. J. Psychiatry 2002, 159, 983–990. [Google Scholar] [CrossRef]
- Buchanan, R.W. Persistent Negative Symptoms in Schizophrenia: An Overview. Schizophr. Bull. 2007, 33, 1013–1022. [Google Scholar] [CrossRef]
- Kirkpatrick, B.; Galderisi, S. Deficit schizophrenia: An update. World Psychiatry 2008, 7, 143–147. [Google Scholar] [CrossRef]
- Galderisi, S.; Maj, M. Deficit schizophrenia: An overview of clinical, biological and treatment aspects. Eur. Psychiatry 2009, 24, 493–500. [Google Scholar] [CrossRef]
- Kirkpatrick, B. Progress in the Study of Negative Symptoms. Schizophr. Bull. 2014, 40, S101–S106. [Google Scholar] [CrossRef] [PubMed]
- Kirkpatrick, B. Recognizing Primary vs. Secondary Negative Symptoms and Apathy vs. Expression Domains. J. Clin. Psychiatry 2014, 75, e09. [Google Scholar] [CrossRef]
- Galderisi, S.; Merlotti, E.; Mucci, A. Neurobiological background of negative symptoms. Eur. Arch. Psychiatry Clin. Neurosci. 2015, 265, 543–558. [Google Scholar] [CrossRef] [PubMed]
- Mucci, A.; Merlotti, E.; Üçok, A.; Aleman, A.; Galderisi, S. Primary and persistent negative symptoms: Concepts, assessments and neurobiological bases. Schizophr. Res. 2016, 186, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Bucci, P.; Galderisi, S. Categorizing and assessing negative symptoms. Curr. Opin. Psychiatry 2017, 30, 201–208. [Google Scholar] [CrossRef]
- Kirkpatrick, B.; Mucci, A.; Galderisi, S. Primary, Enduring Negative Symptoms: An Update on Research. Schizophr. Bull. 2017, 43, 730–736. [Google Scholar] [CrossRef]
- Barch, D.M.; Dowd, E.C. Goal Representations and Motivational Drive in Schizophrenia: The Role of Prefrontal-Striatal Interactions. Schizophr. Bull. 2010, 36, 919–934. [Google Scholar] [CrossRef]
- Foussias, G.; Remington, G. Negative Symptoms in Schizophrenia: Avolition and Occam’s Razor. Schizophr. Bull. 2008, 36, 359–369. [Google Scholar] [CrossRef]
- Bromberg-Martin, E.S.; Matsumoto, M.; Hikosaka, O. Dopamine in Motivational Control: Rewarding, Aversive, and Alerting. Neuron 2010, 68, 815–834. [Google Scholar] [CrossRef]
- Simpson, E.H.; Waltz, J.A.; Kellendonk, C.; Balsam, P.D. Schizophrenia in Translation: Dissecting Motivation in Schizophrenia and Rodents. Schizophr. Bull. 2012, 38, 1111–1117. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Strauss, G.P.; Horan, W.P.; Kirkpatrick, B.; Fischer, B.A.; Keller, W.R.; Miski, P.; Buchanan, R.W.; Green, M.F.; Carpenter, W.T. Deconstructing negative symptoms of schizophrenia: Avolition–apathy and diminished expression clusters predict clinical presentation and functional outcome. J. Psychiatr. Res. 2013, 47, 783–790. [Google Scholar] [CrossRef] [PubMed]
- Morris, R.; Quail, S.; Griffiths, K.R.; Green, M.; Balleine, B.W. Corticostriatal Control of Goal-Directed Action Is Impaired in Schizophrenia. Biol. Psychiatry 2015, 77, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Mucci, A.; Dima, D.; Soricelli, A.; Volpe, U.; Bucci, P.; Frangou, S.; Prinster, A.; Salvatore, M.; Galderisi, S.; Maj, M. Is avolition in schizophrenia associated with a deficit of dorsal caudate activity? A functional magnetic resonance imaging study during reward anticipation and feedback. Psychol. Med. 2015, 45, 1765–1778. [Google Scholar] [CrossRef] [PubMed]
- Salamone, J.D.; Yohn, S.E.; Lopez-Cruz, L.; Miguel, N.S.; Correa, M. Activational and effort-related aspects of motivation: Neural mechanisms and implications for psychopathology. Brain 2016, 139, 1325–1347. [Google Scholar] [CrossRef]
- Giordano, G.M.; Stanziano, M.; Papa, M.; Mucci, A.; Prinster, A.; Soricelli, A.; Galderisi, S. Functional connectivity of the ventral tegmental area and avolition in subjects with schizophrenia: A resting state functional MRI study. Eur. Neuropsychopharmacol. 2018, 28, 589–602. [Google Scholar] [CrossRef]
- Amodio, A.; Quarantelli, M.; Mucci, A.; Prinster, A.; Soricelli, A.; Vignapiano, A.; Giordano, G.M.; Merlotti, E.; Nicita, A.; Galderisi, S. Avolition-Apathy and White Matter Connectivity in Schizophrenia: Reduced Fractional Anisotropy between Amygdala and Insular Cortex. Clin. EEG Neurosci. 2017, 49, 55–65. [Google Scholar] [CrossRef]
- Sanislow, C.A. RDoC at 10: Changing the discourse for psychopathology. World Psychiatry 2020, 19, 311–312. [Google Scholar] [CrossRef]
- Menon, V. Brain networks and cognitive impairment in psychiatric disorders. World Psychiatry 2020, 19, 309–310. [Google Scholar] [CrossRef]
- Barch, D.M.; Pagliaccio, D.; Luking, K. Mechanisms Underlying Motivational Deficits in Psychopathology: Similarities and Differences in Depression and Schizophrenia. Curr Top. Behav. Neurosci. 2016, 27, 411–449. [Google Scholar] [CrossRef]
- Striatal dopamine, reward, and decision making in schizophrenia. Dialog. Clin. Neurosci. 2016, 18, 77–89. [CrossRef]
- Green, M.F.; Horan, W.P.; Barch, D.M.; Gold, J.M. Effort-Based Decision Making: A Novel Approach for Assessing Motivation in Schizophrenia. Schizophr. Bull. 2015, 41, 1035–1044. [Google Scholar] [CrossRef]
- Mucci, A.; Galderisi, S.; Green, M.F.; Nuechterlein, K.; Rucci, P.; Gibertoni, D.; Rossi, A.; Rocca, P.; Bertolino, A.; Bucci, P.; et al. Familial aggregation of MATRICS Consensus Cognitive Battery scores in a large sample of outpatients with schizophrenia and their unaffected relatives. Psychol. Med. 2017, 48, 1359–1366. [Google Scholar] [CrossRef] [PubMed]
- Davidson, M. Cognitive impairment as a diagnostic criterion and treatment target in schizophrenia. World Psychiatry 2019, 18, 171–172. [Google Scholar] [CrossRef] [PubMed]
- Grant, P.M.; Best, M.W.; Beck, A.T. The meaning of group differences in cognitive test performance. World Psychiatry 2019, 18, 163–164. [Google Scholar] [CrossRef]
- Green, M.F.; Horan, W.P.; Lee, J. Nonsocial and social cognition in schizophrenia: Current evidence and future directions. World Psychiatry 2019, 18, 146–161. [Google Scholar] [CrossRef]
- Besteher, B.; Brambilla, P.; Nenadić, I. Twin studies of brain structure and cognition in schizophrenia. Neurosci. Biobehav. Rev. 2019, 109, 103–113. [Google Scholar] [CrossRef]
- Moritz, S.; Silverstein, S.M.; Dietrichkeit, M.; Gallinat, J. Neurocognitive deficits in schizophrenia are likely to be less severe and less related to the disorder than previously thought. World Psychiatry 2020, 19, 254–255. [Google Scholar] [CrossRef]
- Caputo, F.; Giordano, G.M.; Mucci, A.; Galderisi, S. Aetiopathological mechanisms of negative symptoms in schizophrenia. Die Psychiatr. 2016, 13, 121–129. [Google Scholar] [CrossRef]
- Melle, I. Cognition in schizophrenia: A marker of underlying neurodevelopmental problems? World Psychiatry 2019, 18, 164–165. [Google Scholar] [CrossRef] [PubMed]
- Miller, E.M.; Shankar, M.U.; Knutson, B.; McClure, S.M. Dissociating Motivation from Reward in Human Striatal Activity. J. Cogn. Neurosci. 2014, 26, 1075–1084. [Google Scholar] [CrossRef] [PubMed]
- Bissonette, G.B.; Roesch, M.R. Development and function of the midbrain dopamine system: What we know and what we need to. Genes Brain Behav. 2015, 15, 62–73. [Google Scholar] [CrossRef]
- Bissonette, G.B.; Roesch, M.R. Neurophysiology of Reward-Guided Behavior: Correlates Related to Predictions, Value, Motivation, Errors, Attention, and Action. Curr. Top. Behav. Neurosci. 2016, 27, 199–230. [Google Scholar] [CrossRef]
- O’Doherty, J.P. Multiple Systems for the Motivational Control of Behavior and Associated Neural Substrates in Humans. Curr. Top. Behav. Neurosci. 2016, 27, 291–312. [Google Scholar] [CrossRef] [PubMed]
- Cohen, A.; Schwartz, E.; Le, T.P.; Fedechko, T.; Kirkpatrick, B.; Strauss, G.P. Using biobehavioral technologies to effectively advance research on negative symptoms. World Psychiatry 2019, 18, 103–104. [Google Scholar] [CrossRef] [PubMed]
- Bowie, C.R. Cognitive remediation for severe mental illness: State of the field and future directions. World Psychiatry 2019, 18, 274–275. [Google Scholar] [CrossRef]
- Stefani, M.R. Rule Learning and Reward Contingency Are Associated with Dissociable Patterns of Dopamine Activation in the Rat Prefrontal Cortex, Nucleus Accumbens, and Dorsal Striatum. J. Neurosci. 2006, 26, 8810–8818. [Google Scholar] [CrossRef] [PubMed]
- Hikosaka, O.; Bromberg-Martin, E.; Hong, S.; Matsumoto, M. New insights on the subcortical representation of reward. Curr. Opin. Neurobiol. 2008, 18, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Haber, S.N.; Knutson, B. The Reward Circuit: Linking Primate Anatomy and Human Imaging. Neuropsychopharmacology 2009, 35, 4–26. [Google Scholar] [CrossRef] [PubMed]
- Sesack, S.R.; Grace, A.A. Cortico-Basal Ganglia Reward Network: Microcircuitry. Neuropsychopharmacology 2010, 35, 27–47. [Google Scholar] [CrossRef] [PubMed]
- Cascella, N.G.; Fieldstone, S.C.; Rao, V.A.; Pearlson, G.D.; Sawa, A.; Schretlen, D.J. Gray-matter abnormalities in deficit schizophrenia. Schizophr. Res. 2010, 120, 63–70. [Google Scholar] [CrossRef]
- Friston, K.J.; Frith, C.D. Schizophrenia: A disconnection syndrome? Clin. Neurosci. 1995, 3, 89–97. [Google Scholar] [PubMed]
- Buchsbaum, M.S.; Tang, C.Y.; Peled, S.; Gudbjartsson, H.; Lu, D.; Hazlett, E.A.; Downhill, J.; Haznedar, M.; Fallon, J.H.; Atlas, S.W. MRI white matter diffusion anisotropy and PET metabolic rate in schizophrenia. NeuroReport 1998, 9, 425–430. [Google Scholar] [CrossRef]
- Lim, K.O.; Hedehus, M.; Moseley, M.; de Crespigny, A.; Sullivan, E.V.; Pfefferbaum, A. Compromised White Matter Tract Integrity in Schizophrenia Inferred from Diffusion Tensor Imaging. Arch. Gen. Psychiatry 1999, 56, 367–374. [Google Scholar] [CrossRef] [PubMed]
- Ardekani, B.A.; Nierenberg, J.; Hoptman, M.J.; Javitt, D.C.; Lim, K.O. MRI study of white matter diffusion anisotropy in schizophrenia. NeuroReport 2003, 14, 2025–2029. [Google Scholar] [CrossRef][Green Version]
- Hoptman, M.J.; D’Angelo, D.; Catalano, D.; Mauro, C.J.; Shehzad, Z.E.; Kelly, A.M.C.; Castellanos, F.X.; Javitt, D.C.; Milham, M.P. Amygdalofrontal Functional Disconnectivity and Aggression in Schizophrenia. Schizophr. Bull. 2009, 36, 1020–1028. [Google Scholar] [CrossRef]
- Pettersson-Yeo, W.; Allen, P.; Benetti, S.; McGuire, P.; Mechelli, A. Dysconnectivity in schizophrenia: Where are we now? Neurosci. Biobehav. Rev. 2011, 35, 1110–1124. [Google Scholar] [CrossRef]
- Kelly, S.; Jahanshad, N.; Zalesky, A.; Kochunov, P.; Agartz, I.; Alloza, C.; Andreassen, O.A.; Arango, C.; Banaj, N.; Bouix, S.; et al. Widespread white matter microstructural differences in schizophrenia across 4322 individuals: Results from the ENIGMA Schizophrenia DTI Working Group. Mol. Psychiatry 2017, 23, 1261–1269. [Google Scholar] [CrossRef]
- Reininghaus, U.; Böhnke, J.R.; Chavez-Baldini, U.; Gibbons, R.; Ivleva, E.; Clementz, B.A.; Pearlson, G.D.; Keshavan, M.S.; Sweeney, J.A.; Tamminga, C.A. Transdiagnostic dimensions of psychosis in the Bipolar-Schizophrenia Network on Intermediate Phenotypes (B-SNIP). World Psychiatry 2019, 18, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Nath, M.; Wong, T.P.; Srivastava, L.K. Neurodevelopmental insights into circuit dysconnectivity in schizophrenia. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2020, 104, 110047. [Google Scholar] [CrossRef]
- Bullmore, E.; Frangou, S.; Murray, R. The dysplastic net hypothesis: An integration of developmental and dysconnectivity theories of schizophrenia. Schizophr. Res. 1997, 28, 143–156. [Google Scholar] [CrossRef]
- Rowland, L.M.; Spieker, E.A.; Francis, A.; Barker, P.B.; Carpenter, W.T.; Buchanan, R.W. White Matter Alterations in Deficit Schizophrenia. Neuropsychopharmacology 2008, 34, 1514–1522. [Google Scholar] [CrossRef] [PubMed]
- Kitis, O.; Ozalay, O.; Zengin, E.B.; Haznedaroglu, D.; Eker, M.C.; Yalvac, D.; Oguz, K.; Coburn, K.; Gonul, A.S. Reduced left uncinate fasciculus fractional anisotropy in deficit schizophrenia but not in non-deficit schizophrenia. Psychiatry Clin. Neurosci. 2012, 66, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Voineskos, A.N.; Foussias, G.; Lerch, J.; Felsky, D.; Remington, G.; Rajji, T.K.; Lobaugh, N.; Pollock, B.G.; Mulsant, B.H. Neuroimaging Evidence for the Deficit Subtype of Schizophrenia. JAMA Psychiatry 2013, 70, 472–480. [Google Scholar] [CrossRef] [PubMed]
- Spalletta, G.; de Rossi, P.; Piras, F.; Iorio, M.; Dacquino, C.; Scanu, F.; Girardi, P.; Caltagirone, C.; Kirkpatrick, B.; Chiapponi, C. Brain white matter microstructure in deficit and non-deficit subtypes of schizophrenia. Psychiatry Res. Neuroimaging 2015, 231, 252–261. [Google Scholar] [CrossRef]
- Tan, A.S.; Chew, Q.H.; Sim, K. Cerebral white matter changes in deficit and non-deficit subtypes of schizophrenia. J. Neural Transm. 2020, 127, 1073–1079. [Google Scholar] [CrossRef]
- Podwalski, P.; Tyburski, E.; Szczygieł, K.; Waszczuk, K.; Rek-Owodziń, K.; Mak, M.; Plichta, P.; Bielecki, M.; Rudkowski, K.; Kucharska-Mazur, J.; et al. White Matter Integrity of the Corpus Callosum and Psychopathological Dimensions in Deficit and Non-Deficit Schizophrenia Patients. J. Clin. Med. 2021, 10, 2225. [Google Scholar] [CrossRef]
- Kay, S.R.; Fiszbein, A.; Opler, L.A. The Positive and Negative Syndrome Scale (PANSS) for Schizophrenia. Schizophr. Bull. 1987, 13, 261–276. [Google Scholar] [CrossRef]
- Fabrazzo, M.; Prisco, V.; Sampogna, G.; Perris, F.; Catapano, F.; Monteleone, A.M.; Maj, M. Clozapine versus other antipsychotics during the first 18 weeks of treatment: A retrospective study on risk factor increase of blood dyscrasias. Psychiatry Res. 2017, 256, 275–282. [Google Scholar] [CrossRef]
- Kirkpatrick, B.; Buchanan, R.W.; McKenny, P.D.; Alphs, L.D.; Carpenter, W.T. The schedule for the deficit syndrome: An instrument for research in schizophrenia. Psychiatry Res. 1989, 30, 119–123. [Google Scholar] [CrossRef]
- Gardner, D.M.; Murphy, A.L.; O’Donnell, H.; Centorrino, F.; Baldessarini, R.J. International Consensus Study of Antipsychotic Dosing. Am. J. Psychiatry 2010, 167, 686–693. [Google Scholar] [CrossRef] [PubMed]
- Bracht, T.; Horn, H.; Strik, W.; Federspiel, A.; Razavi, N.; Stegmayer, K.; Wiest, R.; Dierks, T.; Müller, T.J.; Walther, S. White matter pathway organization of the reward system is related to positive and negative symptoms in schizophrenia. Schizophr. Res. 2014, 153, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Pizzagalli, D.A.; Holmes, A.; Dillon, D.G.; Goetz, E.L.; Birk, J.; Bogdan, R.; Dougherty, D.D.; Iosifescu, D.V.; Rauch, S.L.; Fava, M. Reduced Caudate and Nucleus Accumbens Response to Rewards in Unmedicated Individuals with Major Depressive Disorder. Am. J. Psychiatry 2009, 166, 702–710. [Google Scholar] [CrossRef] [PubMed]
- Maldjian, J.A.; Laurienti, P.J.; Kraft, R.A.; Burdette, J.H. An automated method for neuroanatomic and cytoarchitectonic atlas-based interrogation of fMRI data sets. NeuroImage 2003, 19, 1233–1239. [Google Scholar] [CrossRef]
- Blood, A.J.; Iosifescu, D.V.; Makris, N.; Perlis, R.H.; Kennedy, D.N.; Dougherty, D.D.; Kim, B.W.; Lee, M.J.; Wu, S.; Lee, S.H.; et al. Microstructural Abnormalities in Subcortical Reward Circuitry of Subjects with Major Depressive Disorder. PLoS ONE 2010, 5, e13945. [Google Scholar] [CrossRef]
- Nieuwenhuys, R.; Voogd, J.; van Huizen, C. The Human Central Nervous System, 4th ed.; Springer: Berlin, Germany, 2008. [Google Scholar]
- Kringelbach, M.L. The human orbitofrontal cortex: Linking reward to hedonic experience. Nat. Rev. Neurosci. 2005, 6, 691–702. [Google Scholar] [CrossRef]
- Makris, N.; Goldstein, J.M.; Kennedy, D.; Hodge, S.M.; Caviness, V.S.; Faraone, S.; Tsuang, M.T.; Seidman, L.J. Decreased volume of left and total anterior insular lobule in schizophrenia. Schizophr. Res. 2006, 83, 155–171. [Google Scholar] [CrossRef]
- Deen, B.; Pitskel, N.B.; Pelphrey, K.A. Three Systems of Insular Functional Connectivity Identified with Cluster Analysis. Cereb. Cortex 2010, 21, 1498–1506. [Google Scholar] [CrossRef]
- Smith, S.M.; Jenkinson, M.; Woolrich, M.W.; Beckmann, C.F.; Behrens, T.E.; Johansen-Berg, H.; Bannister, P.R.; de Luca, M.; Drobnjak, I.; Flitney, D.E.; et al. Advances in functional and structural MR image analysis and implementation as FSL. NeuroImage 2004, 23, S208–S219. [Google Scholar] [CrossRef]
- Leemans, A.; Jones, D.K. TheB-matrix must be rotated when correcting for subject motion in DTI data. Magn. Reson. Med. 2009, 61, 1336–1349. [Google Scholar] [CrossRef]
- Smith, S.M. Fast robust automated brain extraction. Hum. Brain Mapp. 2002, 17, 143–155. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.; Zhanga, Y.; Jenkinson, M.; Chenab, J.; Matthews, P.M.; Federico, A.; de Stefano, N. Accurate, Robust, and Automated Longitudinal and Cross-Sectional Brain Change Analysis. NeuroImage 2002, 17, 479–489. [Google Scholar] [CrossRef]
- Behrens, T.; Johansen-Berg, H.; Jbabdi, S.; Rushworth, M.; Woolrich, M. Probabilistic diffusion tractography with multiple fibre orientations: What can we gain? NeuroImage 2007, 34, 144–155. [Google Scholar] [CrossRef] [PubMed]
- Morris, D.M.; Embleton, K.V.; Parker, G.J. Probabilistic fibre tracking: Differentiation of connections from chance events. NeuroImage 2008, 42, 1329–1339. [Google Scholar] [CrossRef] [PubMed]
- Keshavan, M.; Lizano, P.; Prasad, K. The synaptic pruning hypothesis of schizophrenia: Promises and challenges. World Psychiatry 2020, 19, 110–111. [Google Scholar] [CrossRef]
- Parkes, S.; Balleine, B.W. Incentive Memory: Evidence the Basolateral Amygdala Encodes and the Insular Cortex Retrieves Outcome Values to Guide Choice between Goal-Directed Actions. J. Neurosci. 2013, 33, 8753–8763. [Google Scholar] [CrossRef]
- Parkes, S.; Bradfield, L.; Balleine, B.W. Interaction of Insular Cortex and Ventral Striatum Mediates the Effect of Incentive Memory on Choice between Goal-Directed Actions. J. Neurosci. 2015, 35, 6464–6471. [Google Scholar] [CrossRef] [PubMed]
- Pickens, C.; Saddoris, M.P.; Setlow, B.; Gallagher, M.; Holland, P.C.; Schoenbaum, G. Different Roles for Orbitofrontal Cortex and Basolateral Amygdala in a Reinforcer Devaluation Task. J. Neurosci. 2003, 23, 11078–11084. [Google Scholar] [CrossRef]
- Izquierdo, A.; Suda, R.K.; Murray, E. Bilateral Orbital Prefrontal Cortex Lesions in Rhesus Monkeys Disrupt Choices Guided by Both Reward Value and Reward Contingency. J. Neurosci. 2004, 24, 7540–7548. [Google Scholar] [CrossRef]
- Ostlund, S.B.; Balleine, B. Orbitofrontal Cortex Mediates Outcome Encoding in Pavlovian but Not Instrumental Conditioning. J. Neurosci. 2007, 27, 4819–4825. [Google Scholar] [CrossRef] [PubMed]
- Burke, K.A.; Franz, T.M.; Miller, D.N.; Schoenbaum, G. The role of the orbitofrontal cortex in the pursuit of happiness and more specific rewards. Nature 2008, 454, 340–344. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, Y.K.; Roesch, M.R.; Stalnaker, T.A.; Haney, R.Z.; Calu, D.; Taylor, A.R.; Burke, K.A.; Schoenbaum, G. The Orbitofrontal Cortex and Ventral Tegmental Area Are Necessary for Learning from Unexpected Outcomes. Neuron 2009, 62, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Kahnt, T.; Heinzle, J.; Park, S.; Haynes, J.-D. The neural code of reward anticipation in human orbitofrontal cortex. Proc. Natl. Acad. Sci. USA 2010, 107, 6010–6015. [Google Scholar] [CrossRef]
- Everitt, B.; Cador, M.; Robbins, T. Interactions between the amygdala and ventral striatum in stimulus-reward associations: Studies using a second-order schedule of sexual reinforcement. Neuroscience 1989, 30, 63–75. [Google Scholar] [CrossRef]
- Everitt, B.J.; Parkinson, J.A.; Olmstead, M.C.; Arroyo, M.; Robledo, P.; Robbins, T. Associative Processes in Addiction and Reward the Role of Amygdala-Ventral Striatal Subsystems. Ann. N. Y. Acad. Sci. 1999, 877, 412–438. [Google Scholar] [CrossRef]
- Baxter, M.G.; Murray, E. The amygdala and reward. Nat. Rev. Neurosci. 2002, 3, 563–573. [Google Scholar] [CrossRef]
- Murray, E.A. The amygdala, reward and emotion. Trends Cogn. Sci. 2007, 11, 489–497. [Google Scholar] [CrossRef]
- Ramirez, D.R.; Savage, L.M. Differential involvement of the basolateral amygdala, orbitofrontal cortex, and nucleus accumbens core in the acquisition and use of reward expectancies. Behav. Neurosci. 2007, 121, 896–906. [Google Scholar] [CrossRef]
- Harsay, H.A.; Spaan, M.; Wijnen, J.G.; Ridderinkhof, K.R. Error Awareness and Salience Processing in the Oddball Task: Shared Neural Mechanisms. Front. Hum. Neurosci. 2012, 6, 246. [Google Scholar] [CrossRef]
- Liang, X.; He, Y.; Salmeron, B.J.; Gu, H.; Stein, E.A.; Yang, Y. Interactions between the Salience and Default-Mode Networks Are Disrupted in Cocaine Addiction. J. Neurosci. 2015, 35, 8081–8090. [Google Scholar] [CrossRef] [PubMed]
- Rogers-Carter, M.; Christianson, J.P. An insular view of the social decision-making network. Neurosci. Biobehav. Rev. 2019, 103, 119–132. [Google Scholar] [CrossRef] [PubMed]
- Rogers-Carter, M.M.; Djerdjaj, A.; Gribbons, K.B.; Varela, J.A.; Christianson, J.P. Insular Cortex Projections to Nucleus Accumbens Core Mediate Social Approach to Stressed Juvenile Rats. J. Neurosci. 2019, 39, 8717–8729. [Google Scholar] [CrossRef] [PubMed]
- Barch, D.M. Nonsocial and social cognitive function in psychosis: Interrelationships, specificity and innovative approaches. World Psychiatry 2019, 18, 117–118. [Google Scholar] [CrossRef]
- Kotov, R.; Jonas, K.G.; Carpenter, W.T.; Dretsch, M.N.; Eaton, N.R.; Forbes, M.K.; Forbush, K.T.; Hobbs, K.; Reininghaus, U.; Slade, T.; et al. Validity and utility of Hierarchical Taxonomy of Psychopathology (HiTOP): I. Psychosis superspectrum. World Psychiatry 2020, 19, 151–172. [Google Scholar] [CrossRef]
- Rodgers, K.M.; Benison, A.M.; Klein, A.; Barth, D.S. Auditory, Somatosensory, and Multisensory Insular Cortex in the Rat. Cereb. Cortex 2008, 18, 2941–2951. [Google Scholar] [CrossRef]
- Gogolla, N.; Takesian, A.E.; Feng, G.; Fagiolini, M.; Hensch, T.K. Sensory Integration in Mouse Insular Cortex Reflects GABA Circuit Maturation. Neuron 2014, 83, 894–905. [Google Scholar] [CrossRef]
- Gogolla, N. The insular cortex. Curr. Biol. 2017, 27, R580–R586. [Google Scholar] [CrossRef]
- Wright, C.; Groenewegen, H. Patterns of overlap and segregation between insular cortical, intermediodorsal thalamic and basal amygdaloid afferents in the nucleus accumbens of the rat. Neuroscience 1996, 73, 359–373. [Google Scholar] [CrossRef]
- Wise, R.A. Brain Reward Circuitry: Insights from Unsensed Incentives. Neuron 2002, 36, 229–240. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| HCs (n = 35) | NDS (n = 37) | DS (n = 9) | F | p | |
|---|---|---|---|---|---|
| Age (years) | 32.94 ± 8.80 | 36.57 ± 7.50 | 33.00 ± 8.53 | 1.952 | 0.149 |
| Gender (M/F) | 17/18 | 25/12 | 5/4 | 1.340 | 0.268 |
| Paternal education (years) | 11.31 ± 5.85 | 8.41 ± 4.64 | 9.00 ± 4.09 | 2.965 | 0.057 |
| Maternal education (years) | 10.34 ± 5.67 | 8.49 ± 4.69 | 8.33 ± 4.47 | 1.352 | 0.265 |
| RMS | 0.34 ± 0.10 | 0.41 ± 0.10 | 0.40 ± 0.11 | 3.131 | 0.049 * |
| Total SDS | - | 7.82 ± 5.60 | 11.00 ± 6.70 | 1.740 | 0.195 |
| SDS Experiential domain | - | 4.76 ± 3.45 | 6.29 ± 3.20 | 1.161 | 0.288 |
| SDS Expressive Deficit domain | - | 3.06 ± 2.47 | 4.71 ± 3.59 | 2.196 | 0.147 |
| PANSS Positive | - | 8.09 ± 4.28 | 6.00 ± 2.45 | 1.541 | 0.222 |
| PANSS Disorganization | - | 7.33 ± 3.68 | 7.43 ± 4.28 | 0.004 | 0.952 |
| PANSS Depression | - | 2.49 ± 0.85 | 1.43 ± 0.50 | 10.224 | 0.003 |
| Chlorpromazine equivalent doses | - | 402.01 ± 190.05 | 263.37 ± 92.34 | 3.003 | 0.092 |
| Brain Pathways | NDS (n = 37) | DS (n = 9) | HCs (n = 35) | F | p |
|---|---|---|---|---|---|
| CI | |||||
| lNAcc to daIC | 10.52 ± 14.31 | 6.02 ± 6.64 | 9.36 ± 10.88 | 0.831 | 0.440 |
| lNAcc to DLPFC | 41.35 ± 54.84 | 15.23 ± 19.29 | 49.31 ± 101.35 | 0.654 | 0.523 |
| lNAcc to lOFC | 119.56 ± 209.92 | 105.95 ± 116.48 | 120.02 ± 156.09 | 0.114 | 0.892 |
| lNAcc to mOFC | 2192.74 ± 1283.04 | 1975.77 ± 859.34 | 2903.13 ± 1783.21 | 2.023 | 0.139 |
| lNAcc to pIC | 552.43 ±591.62 | 1132.7 ± 850.63 | 463.43 ± 408.79 | 4.823 | 0.011 |
| lNAcc to vaIC | 646.07 ± 509.67 | 1246.7 ± 1052.93 | 920.0 ± 881.79 | 2.453 | 0.093 |
| lAmy to daIC | 66.28 ± 66.38 | 29.66 ± 24.57 | 69.33 ± 69.13 | 1.532 | 0.223 |
| lAmy to DLPFC | 40.26 ± 35.93 | 23.7 ± 17.01 | 53.3 ± 46.16 | 1.795 | 0.173 |
| lAmy to lOFC | 160.01 ± 175.11 | 74.5 ±31.49 | 14,328 ± 160.88 | 1.922 | 0.153 |
| lAmy to mOFC | 832.51 ±547.32 | 598.92 ±337.07 | 1012.18 ± 647.54 | 1.662 | 0.197 |
| lAmy to pIC | 2231.79 ± 1861.32 | 3076.22 ± 1806.24 | 1748.80 ± 1274.76 | 3.323 | 0.041 |
| lAmy to vaIC | 2418.53 ± 1169.48 | 2452.89 ± 916.74 | 2908.10 ± 1094.67 | 1.424 | 0.247 |
| lVTA to daIC | 57.8 ± 89.75 | 12.48 ± 13.78 | 33.06 ± 41.98 | 1.428 | 0.246 |
| lVTA to DLPFC | 125.04 ± 157.90 | 76.05 ± 100.19 | 129.57 ± 85.92 | 1.101 | 0.338 |
| lVTA to lOFC | 90.15 ± 79.93 | 36.76 ± 32.64 | 126.96 ± 140.15 | 2.560 | 0.084 |
| lVTA to mOFC | 66.57 ± 67.51 | 80.52 ± 153.03 | 102.19 ± 131.15 | 1.027 | 0.363 |
| lVTA to pIC | 76.27 ± 91.95 | 59.45 ± 68.97 | 46.88 ± 58.16 | 1.251 | 0.292 |
| lVTA to vaIC | 17.21 ± 27.71 | 8.95 ± 3.49 | 16.54 ± 17.90 | 0.763 | 0.470 |
| rNAcc to daIC | 11.09 ± 44.34 | 12.36 ± 14.58 | 7.99 ± 16.06 | 0.088 | 0.916 |
| rNAcc to DLPFC | 18.54 ± 26.02 | 17.21 ± 23.43 | 31.69 ± 59.31 | 0.874 | 0.421 |
| rNAcc to lOFC | 449.74 ± 512.68 | 552.32 ± 553.16 | 583.00 ± 575.02 | 0.099 | 0.906 |
| rNAcc to mOFC | 1352 ± 943.98 | 1660.94 ± 1159.38 | 2216.77 ± 1315.54 | 3.717 | 0.029 |
| rNAcc to pIC | 129.18 ± 244.89 | 89.27 ± 110.05 | 80.98 ± 92.41 | 0.119 | 0.888 |
| rNAcc to vaIC | 732.42 ± 961.42 | 1369.91 ± 1410.50 | 718.38 ± 700.50 | 2.082 | 0.132 |
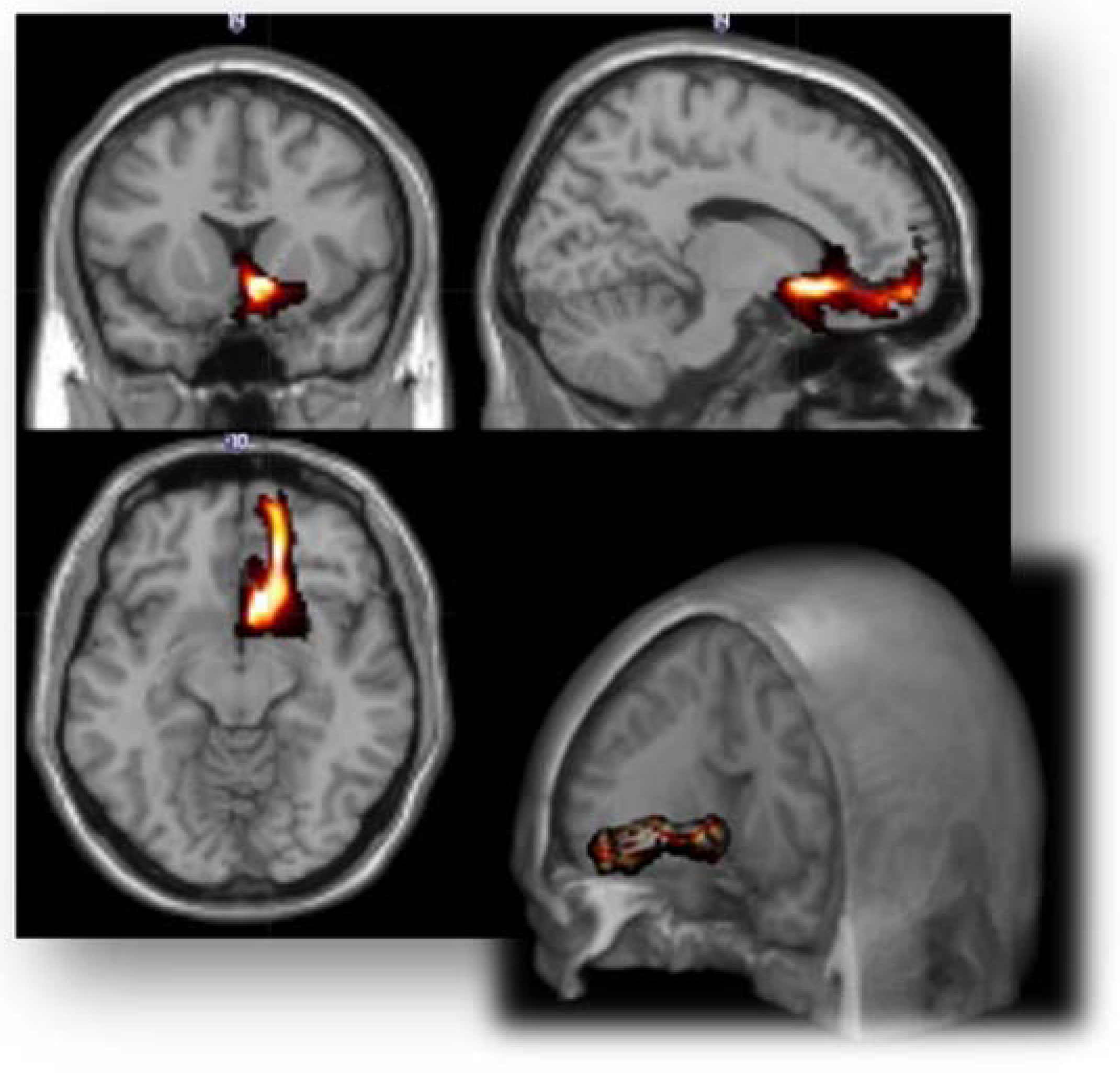
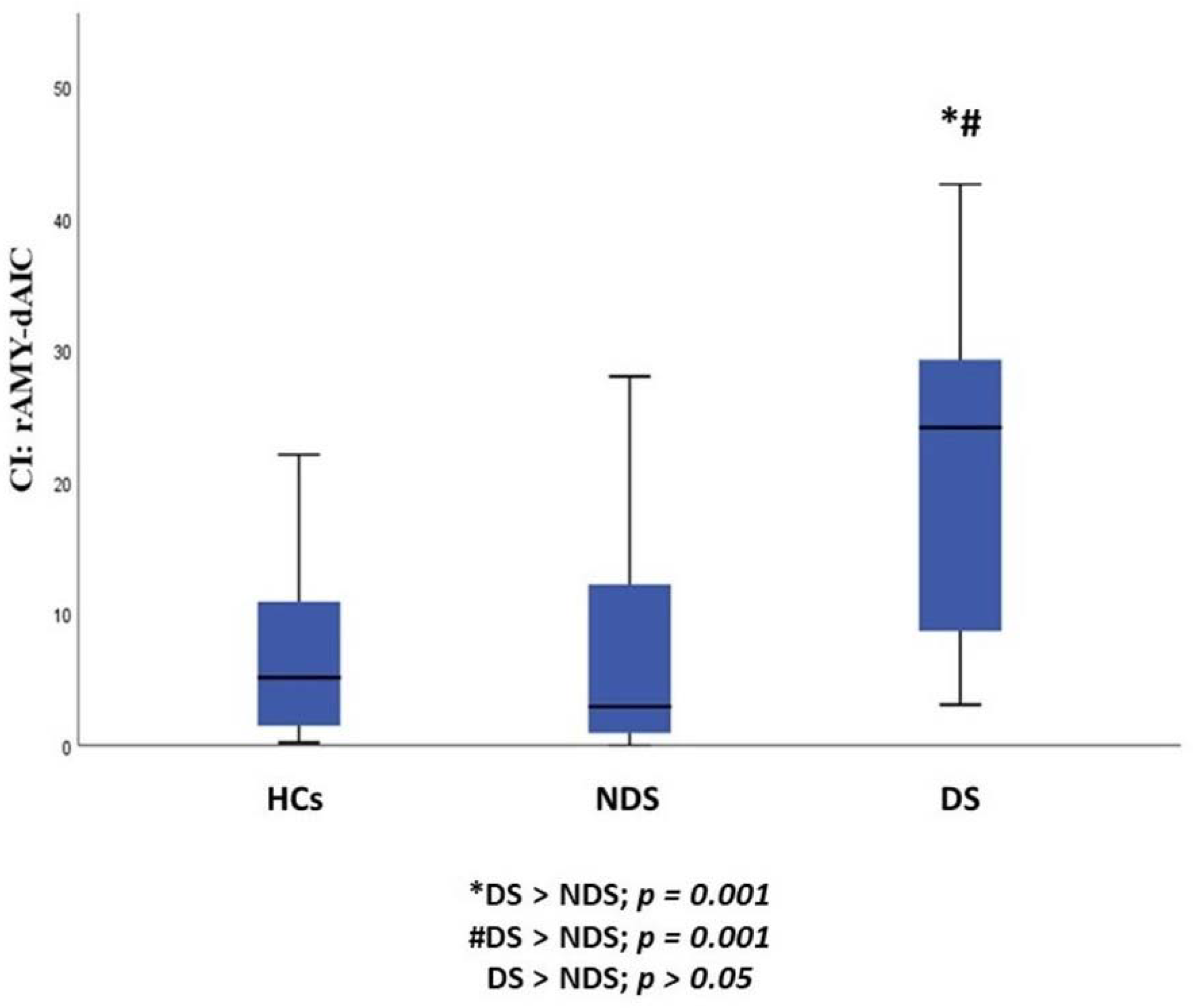
| rAmy to daIC | 8.39 ± 11.46 | 25.53 ± 21.64 | 7.82 ± 9.24 | 8.190 | 0.001 |
| rAmy to DLPFC | 20.1 ±24.27 | 18.14 ± 9.89 | 38.16 ± 32.92 | 4.356 | 0.016 |
| rAmy to lOFC | 95.41 ± 101.87 | 59.48 ± 39.84 | 129.18 ± 107.97 | 1.436 | 0.244 |
| rAmy to mOFC | 1246.66 ± 1069.13 | 937.87 ± 852.42 | 1075.12 ± 992.10 | 0.389 | 0.679 |
| rAmy to pIC | 77.06 ± 144.08 | 118.83 ± 138.32 | 70.62 ± 80.30 | 0.985 | 0.378 |
| rAmy to vaIC | 736.59 ± 824.11 | 818.15 ± 756.58 | 631.73 ± 508.82 | 0.502 | 0.607 |
| rVTA to daIC | 23.3 ± 40.78 | 17.45 ± 28.28 | 35.64 ± 59.95 | 0.683 | 0.508 |
| rVTA to DLPFC | 122.45 ± 94.52 | 77.53 ± 68.19 | 148.46 ± 124.59 | 2.100 | 0.130 |
| rVTA to lOFC | 149.33 ± 169.72 | 64.63 ± 82.54 | 134.87 ± 151.53 | 1.262 | 0.289 |
| rVTA to mOFC | 67.24 ± 93.39 | 26.85 ± 29.73 | 52.22 ± 91.28 | 0.656 | 0.522 |
| rVTA to pIC | 14.77 ± 17.26 | 30.91 ± 69.09 | 14.17 ± 18.00 | 1.283 | 0.283 |
| rVTA to vaIC | 10.68 ± 11.90 | 13.23 ± 17.42 | 17.54 ± 18.96 | 0.543 | 0.583 |
| Brain Pathways | NDS (n = 37) | DS (n = 9) | HCs (n = 35) | F | p |
|---|---|---|---|---|---|
| FA | |||||
| lNAcc to daIC | 1.54 ± 1.27 | 1.26 ± 0.81 | 1.60 ± 1.20 | 0.273 | 0.762 |
| lNAcc to DLPFC | 3.35 ± 3.76 | 1.69 ± 1.25 | 3.27 ± 3.55 | 0.967 | 0.385 |
| lNAcc to lOFC | 7.14 ± 10.8 | 6.45 ± 4.54 | 7.32 ± 7.66 | 0.081 | 0.923 |
| lNAcc to mOFC | 49.93 ± 29.7 | 61.72 ± 41.81 | 63.01 ± 46.42 | 0.857 | 0.429 |
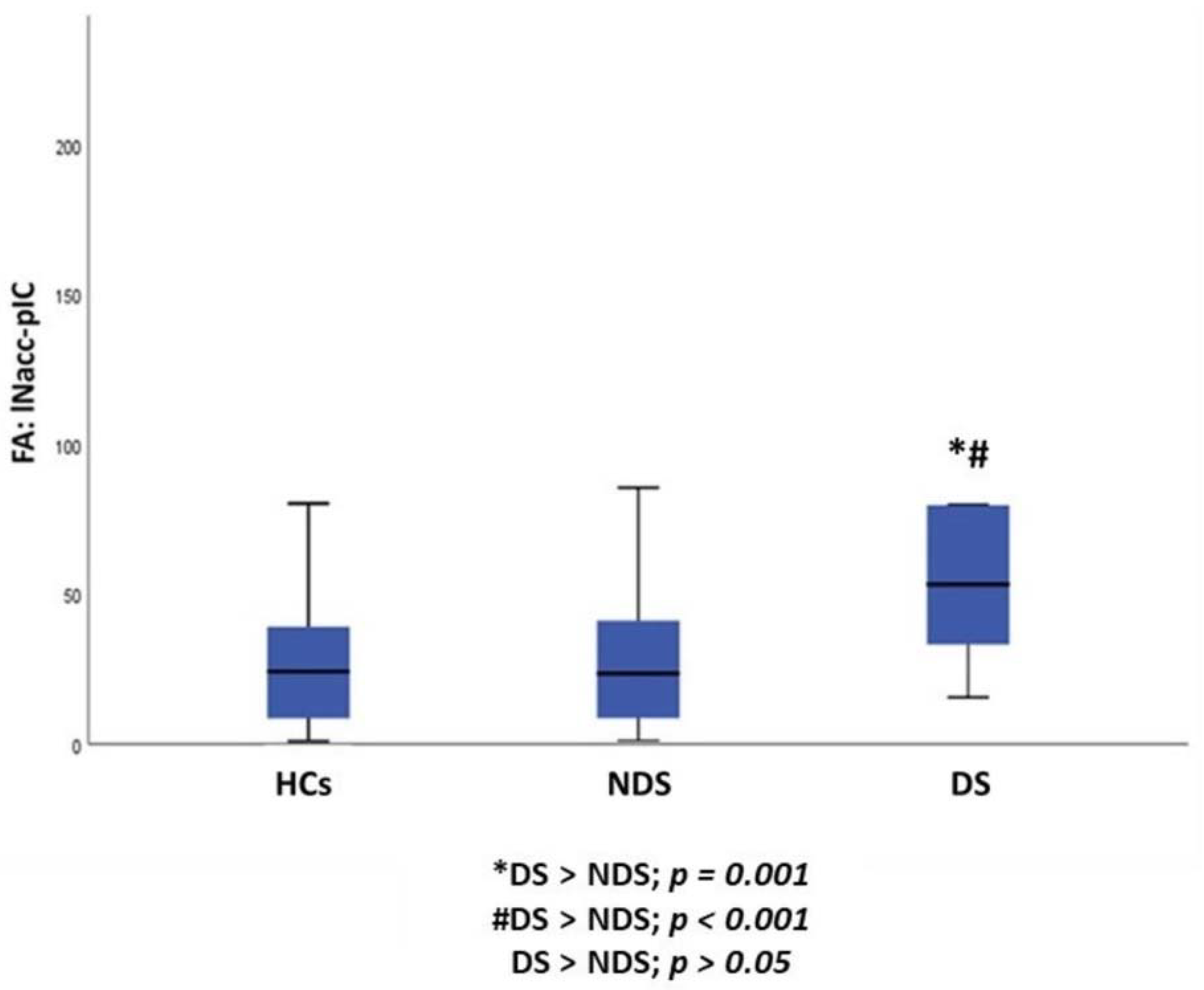
| lNAcc to pIC | 31.77 ± 27.32 | 76.48 ± 68.31 | 28.20 ± 22.57 | 7.760 | 0.001 |
| lNAcc to vaIC | 27.45 ± 22.09 | 52.3 ± 39.84 | 29.48 ± 20.19 | 4.202 | 0.019 |
| lAmy to daIC | 6.85 ± 5.22 | 4.01 ± 1.93 | 6.78 ± 5.80 | 1.645 | 0.200 |
| lAmy to DLPFC | 1047.97 ± 167.3 | 1099.55 ± 207 | 1030.49 ± 232.41 | 0.347 | 0.708 |
| lAmy to lOFC | 0.99 ± 1.36 | 0.57 ± 0.32 | 0.86 ± 0.91 | 0.792 | 0.457 |
| lAmy to mOFC | 12.80 ± 17.30 | 5.14 ± 5.73 | 6.74 ± 9.25 | 0.935 | 0.397 |
| lAmy to pIC | 51.63 ± 36.29 | 74.7 ± 31.73 | 45.06 ± 27.15 | 3.351 | 0.040 |
| lAmy to vaIC | 43.46 ± 17.84 | 43.71 ± 14.56 | 51.67 ± 22.79 | 2.098 | 0.130 |
| lVTA to daIC | 5.00 ± 5.80 | 2.11 ± 0.71 | 4.12 ± 3.99 | 1.079 | 0.345 |
| lVTA to DLPFC | 8.29 ± 8.86 | 5.84 ± 5.69 | 8.36 ± 4.55 | 0.848 | 0.432 |
| lVTA to lOFC | 8.53 ± 6.07 | 5.16 ± 4.09 | 10.04 ± 8.15 | 1.982 | 0.145 |
| lVTA to mOFC | 6.11 ± 4.86 | 5.24 ± 5.8 | 8.04 ± 6.98 | 1.967 | 0.147 |
| lVTA to pIC | 5.99 ± 5.42 | 4.66 ± 3.48 | 3.76 ± 3.35 | 2.324 | 0.105 |
| lVTA to vaIC | 1.96 ± 1.45 | 1.62 ± 0.5 | 2.01 ± 1.12 | 0.389 | 0.679 |
| rNAcc to daIC | 1.58 ± 3.42 | 2.13 ± 1.72 | 1.14 ± 0.94 | 0.540 | 0.585 |
| rNAcc to DLPFC | 2.20 ± 2.93 | 1.67 ± 1.47 | 2.82 ± 4.03 | 0.454 | 0.637 |
| rNAcc to lOFC | 20.76 ± 21.26 | 24.71 ± 25.40 | 24.17 ± 20.67 | 0.019 | 0.981 |
| rNAcc to mOFC | 55.36 ± 42.34 | 65.68 ± 44.25 | 81.19 ± 54.45 | 1.465 | 0.238 |
| rNAcc to pIC | 9.65 ± 14.76 | 7.84 ± 8.17 | 7.32 ± 6.61 | 0.065 | 0.937 |
| rNAcc to vaIC | 33.99 ± 34.61 | 50.40 ± 39.62 | 26.54 ± 19.49 | 2.392 | 0.098 |
| rAmy to daIC | 2.00 ± 1.75 | 3.99 ± 2.16 | 1.85 ± 1.24 | 6.792 | 0.002 |
| rAmy to DLPFC | 17.86 ± 10.45 | 13.17 ± 10.71 | 15.21 ± 11.37 | 0.788 | 0.459 |
| rAmy to lOFC | 1.43 ± 1.53 | 0.65 ± 0.48 | 1.72 ± 2.99 | 1.692 | 0.191 |
| rAmy to mOFC | 8.49 ± 8.26 | 7.91 ± 7.39 | 9.39 ± 9.86 | 0.114 | 0.892 |
| rAmy to pIC | 8.07 ± 11.83 | 9.18 ± 6.32 | 7.21 ± 5.35 | 0.366 | 0.695 |
| rAmy to vaIC | 17.07 ± 11.95 | 20.13 ± 14.73 | 13.61 ± 7.93 | 1.740 | 0.183 |
| rVTA to daIC | 3.75 ± 7.44 | 2.73 ± 2.9 | 3.97 ± 4.86 | 0.214 | 0.808 |
| rVTA to DLPFC | 8.49 ± 5.63 | 5.57 ± 4.14 | 8.44 ± 5.63 | 1.222 | 0.300 |
| rVTA to lOFC | 12.23 ± 11.34 | 6.27 ± 7.3 | 9.24 ± 8.24 | 1.782 | 0.175 |
| rVTA to mOFC | 6.81 ± 5.86 | 3.36 ± 3.49 | 5.17 ± 8.93 | 0.750 | 0.476 |
| rVTA to pIC | 1.88 ± 1.54 | 2.99 ± 4.41 | 1.76 ± 1.26 | 1.437 | 0.244 |
| rVTA to vaIC | 1.66 ± 1.13 | 1.95 ± 1.64 | 2.10 ± 1.38 | 0.357 | 0.701 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giordano, G.M.; Pezzella, P.; Quarantelli, M.; Bucci, P.; Prinster, A.; Soricelli, A.; Perrottelli, A.; Giuliani, L.; Fabrazzo, M.; Galderisi, S. Investigating the Relationship between White Matter Connectivity and Motivational Circuits in Subjects with Deficit Schizophrenia: A Diffusion Tensor Imaging (DTI) Study. J. Clin. Med. 2022, 11, 61. https://doi.org/10.3390/jcm11010061
Giordano GM, Pezzella P, Quarantelli M, Bucci P, Prinster A, Soricelli A, Perrottelli A, Giuliani L, Fabrazzo M, Galderisi S. Investigating the Relationship between White Matter Connectivity and Motivational Circuits in Subjects with Deficit Schizophrenia: A Diffusion Tensor Imaging (DTI) Study. Journal of Clinical Medicine. 2022; 11(1):61. https://doi.org/10.3390/jcm11010061
Chicago/Turabian StyleGiordano, Giulia M., Pasquale Pezzella, Mario Quarantelli, Paola Bucci, Anna Prinster, Andrea Soricelli, Andrea Perrottelli, Luigi Giuliani, Michele Fabrazzo, and Silvana Galderisi. 2022. "Investigating the Relationship between White Matter Connectivity and Motivational Circuits in Subjects with Deficit Schizophrenia: A Diffusion Tensor Imaging (DTI) Study" Journal of Clinical Medicine 11, no. 1: 61. https://doi.org/10.3390/jcm11010061
APA StyleGiordano, G. M., Pezzella, P., Quarantelli, M., Bucci, P., Prinster, A., Soricelli, A., Perrottelli, A., Giuliani, L., Fabrazzo, M., & Galderisi, S. (2022). Investigating the Relationship between White Matter Connectivity and Motivational Circuits in Subjects with Deficit Schizophrenia: A Diffusion Tensor Imaging (DTI) Study. Journal of Clinical Medicine, 11(1), 61. https://doi.org/10.3390/jcm11010061