
Early Reconstitution of Antibody Secreting Cells after Allogeneic Stem Cell Transplantation

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Conditioning Regimen and Haploidentical Hematopoietic Cell Transplantation (HHCT)
2.3. Flow-Cytometry Based Analysis of Immune Reconstitution and Chimerism
2.4. ELISpot Assay to Detect IgM, IgG and IgG Antibody Secreting Cells
2.5. Detection of Immunoglobulin and Cytokine Levels Using Multiplex Immunoassay System
2.6. ELISA
2.7. Statistical Analysis
3. Results
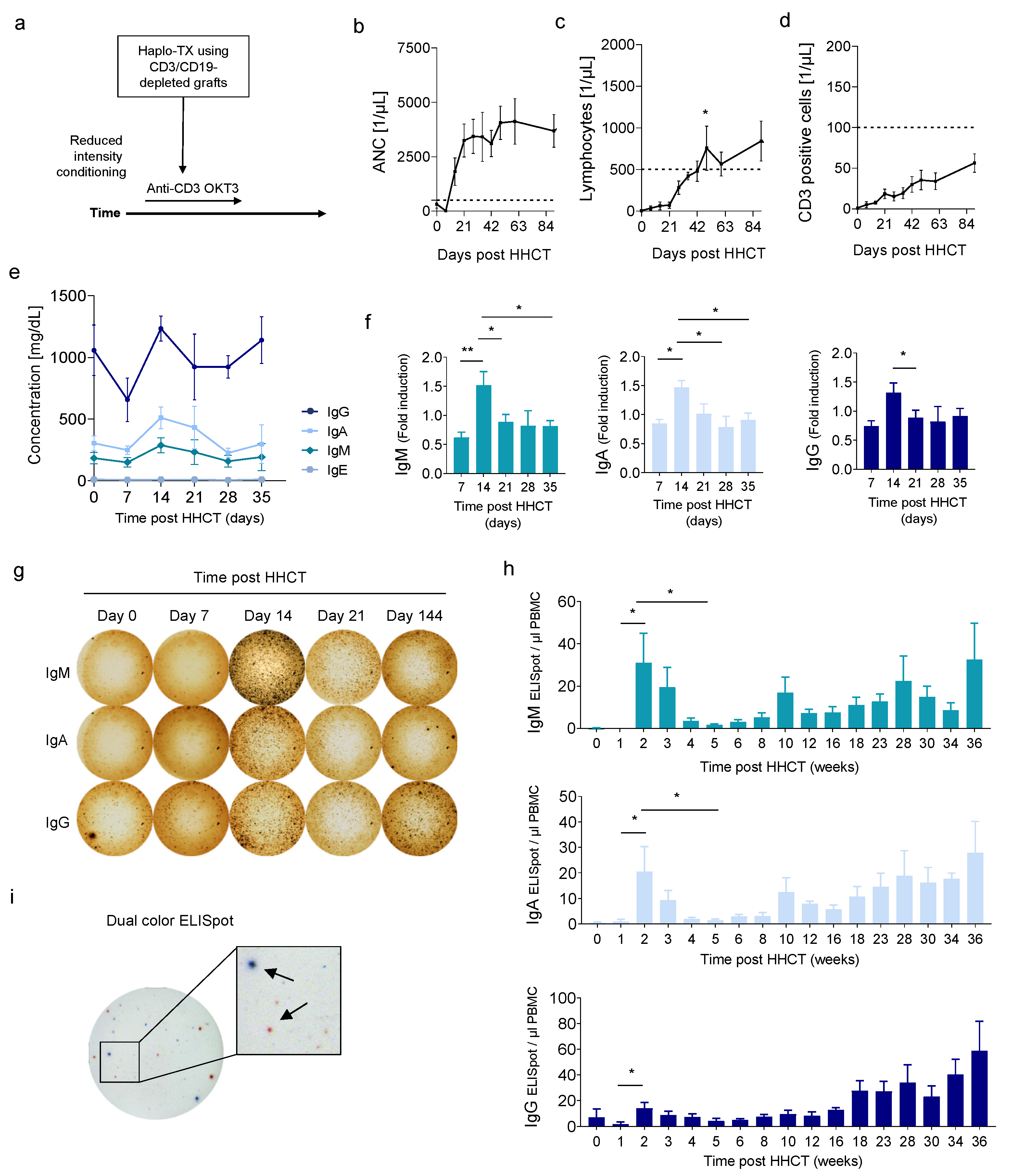
3.1. Early Increase of Serum Immunoglobulins and ASCs after HHCT with T and B Cell-Depleted Grafts
3.2. Correlation of CD19− CD27− CD38low/+ CD138− Cells with Early ASC after HHCT
3.3. Cytokine Levels during Immune Cell Reconstitution
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ogonek, J.; Kralj Juric, M.; Ghimire, S.; Varanasi, P.R.; Holler, E.; Greinix, H.; Weissinger, E. Immune Reconstitution after Allogeneic Hematopoietic Stem Cell Transplantation. Front. Immunol. 2016, 7, 507. [Google Scholar] [CrossRef] [Green Version]
- van den Brink, M.R.; Velardi, E.; Perales, M.A. Immune reconstitution following stem cell transplantation. Hematol. Am. Soc. Hematol. Educ. Program 2015, 2015, 215–219. [Google Scholar] [CrossRef] [Green Version]
- Storek, J.; Geddes, M.; Khan, F.; Huard, B.; Helg, C.; Chalandon, Y.; Passweg, J.; Roosnek, E. Reconstitution of the immune system after hematopoietic stem cell transplantation in humans. Semin. Immunopathol. 2008, 30, 425–437. [Google Scholar] [CrossRef]
- Chaudhry, M.S.; Velardi, E.; Malard, F.; van den Brink, M.R. Immune Reconstitution after Allogeneic Hematopoietic Stem Cell Transplantation: Time to T up the Thymus. J. Immunol. 2017, 198, 40–46. [Google Scholar] [CrossRef]
- Fujimaki, K.; Maruta, A.; Yoshida, M.; Kodama, F.; Matsuzaki, M.; Fujisawa, S.; Kanamori, H.; Ishigatsubo, Y. Immune reconstitution assessed during five years after allogeneic bone marrow transplantation. Bone Marrow Transpl. 2001, 27, 1275–1281. [Google Scholar] [CrossRef] [Green Version]
- Heining, C.; Spyridonidis, A.; Bernhardt, E.; Schulte-Mönting, J.; Behringer, D.; Grüllich, C.; Jakob, A.; Bertz, H.; Finke, J. Lymphocyte reconstitution following allogeneic hematopoietic stem cell transplantation: A retrospective study including 148 patients. Bone Marrow Transpl. 2007, 39, 613–622. [Google Scholar] [CrossRef] [PubMed]
- Le, R.Q.; Melenhorst, J.J.; Battiwalla, M.; Hill, B.; Memon, S.; Savani, B.N.; Shenoy, A.; Hensel, N.F.; Koklanaris, E.K.; Keyvanfar, K.; et al. Evolution of the donor T-cell repertoire in recipients in the second decade after allogeneic stem cell transplantation. Blood 2011, 117, 5250–5256. [Google Scholar] [CrossRef]
- Marie-Cardine, A.; Divay, F.; Dutot, I.; Green, A.; Perdrix, A.; Boyer, O.; Contentin, N.; Tilly, H.; Tron, F.; Vannier, J.P.; et al. Transitional B cells in humans: Characterization and insight from B lymphocyte reconstitution after hematopoietic stem cell transplantation. Clin. Immunol. 2008, 127, 14–25. [Google Scholar] [CrossRef] [PubMed]
- Holodick, N.E.; Rodríguez-Zhurbenko, N.; Hernández, A.M. Defining Natural Antibodies. Front. Immunol. 2017, 8, 872. [Google Scholar] [CrossRef] [Green Version]
- Aziz, M.; Holodick, N.E.; Rothstein, T.L.; Wang, P. The role of B-1 cells in inflammation. Immunol. Res. 2015, 63, 153–166. [Google Scholar] [CrossRef] [PubMed]
- Tsay, G.J.; Zouali, M. The Interplay between Innate-Like B Cells and Other Cell Types in Autoimmunity. Front. Immunol. 2018, 9, 1064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffin, D.O.; Holodick, N.E.; Rothstein, T.L. Human B1 cells in umbilical cord and adult peripheral blood express the novel phenotype CD20+CD27+CD43+CD70−. J. Exp. Med. 2011, 208, 67–80. [Google Scholar] [CrossRef]
- Bethge, W.A.; Haegele, M.; Faul, C.; Lang, P.; Schumm, M.; Bornhauser, M.; Handgretinger, R.; Kanz, L. Haploidentical allogeneic hematopoietic cell transplantation in adults with reduced-intensity conditioning and CD3/CD19 depletion: Fast engraftment and low toxicity. Exp. Hematol. 2006, 34, 1746–1752. [Google Scholar] [CrossRef]
- Federmann, B.; Bornhauser, M.; Meisner, C.; Kordelas, L.; Beelen, D.W.; Stuhler, G.; Stelljes, M.; Schwerdtfeger, R.; Christopeit, M.; Behre, G.; et al. Haploidentical allogeneic hematopoietic cell transplantation in adults using CD3/CD19 depletion and reduced intensity conditioning: A phase II study. Haematologica 2012, 97, 1523–1531. [Google Scholar] [CrossRef] [Green Version]
- Bader, P.; Beck, J.; Frey, A.; Schlegel, P.G.; Hebarth, H.; Handgretinger, R.; Einsele, H.; Niemeyer, C.; Benda, N.; Faul, C.; et al. Serial and quantitative analysis of mixed hematopoietic chimerism by PCR in patients with acute leukemias allows the prediction of relapse after allogeneic BMT. Bone Marrow Transpl. 1998, 21, 487–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crotty, S.; Aubert, R.D.; Glidewell, J.; Ahmed, R. Tracking human antigen-specific memory B cells: A sensitive and generalized ELISPOT system. J. Immunol. Methods 2004, 286, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Bonsignori, M.; Moody, M.A.; Parks, R.J.; Holl, T.M.; Kelsoe, G.; Hicks, C.B.; Vandergrift, N.; Tomaras, G.D.; Haynes, B.F. HIV-1 envelope induces memory B cell responses that correlate with plasma antibody levels after envelope gp120 protein vaccination or HIV-1 infection. J. Immunol. 2009, 183, 2708–2717. [Google Scholar] [CrossRef]
- Vazquez, M.I.; Catalan-Dibene, J.; Zlotnik, A. B cells responses and cytokine production are regulated by their immune microenvironment. Cytokine 2015, 74, 318–326. [Google Scholar] [CrossRef] [Green Version]
- van der Maas, N.G.; Berghuis, D.; van der Burg, M.; Lankester, A.C. B Cell Reconstitution and Influencing Factors after Hematopoietic Stem Cell Transplantation in Children. Front. Immunol. 2019, 10, 782. [Google Scholar] [CrossRef]
- Avery, D.T.; Kalled, S.L.; Ellyard, J.I.; Ambrose, C.; Bixler, S.A.; Thien, M.; Brink, R.; Mackay, F.; Hodgkin, P.D.; Tangye, S.G. BAFF selectively enhances the survival of plasmablasts generated from human memory B cells. J. Clin. Investig. 2003, 112, 286–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antin, J.H.; Emerson, S.G.; Martin, P.; Gadol, N.; Ault, K.A. Leu-1+ (CD5+) B cells. A major lymphoid subpopulation in human fetal spleen: Phenotypic and functional studies. J. Immunol. 1986, 136, 505–510. [Google Scholar] [PubMed]
- Moon, B.G.; Takaki, S.; Miyake, K.; Takatsu, K. The role of IL-5 for mature B-1 cells in homeostatic proliferation, cell survival, and Ig production. J. Immunol. 2004, 172, 6020–6029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baumgarth, N. B-1 Cell Heterogeneity and the Regulation of Natural and Antigen-Induced IgM Production. Front. Immunol. 2016, 7, 324. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hinterleitner, M.; Hinterleitner, C.; Malenke, E.; Federmann, B.; Holzer, U.; Müller, M.; Bethge, W.A.; Wirths, S. Early Reconstitution of Antibody Secreting Cells after Allogeneic Stem Cell Transplantation. J. Clin. Med. 2022, 11, 270. https://doi.org/10.3390/jcm11010270
Hinterleitner M, Hinterleitner C, Malenke E, Federmann B, Holzer U, Müller M, Bethge WA, Wirths S. Early Reconstitution of Antibody Secreting Cells after Allogeneic Stem Cell Transplantation. Journal of Clinical Medicine. 2022; 11(1):270. https://doi.org/10.3390/jcm11010270
Chicago/Turabian StyleHinterleitner, Martina, Clemens Hinterleitner, Elke Malenke, Birgit Federmann, Ursula Holzer, Martin Müller, Wolfgang A. Bethge, and Stefan Wirths. 2022. "Early Reconstitution of Antibody Secreting Cells after Allogeneic Stem Cell Transplantation" Journal of Clinical Medicine 11, no. 1: 270. https://doi.org/10.3390/jcm11010270
APA StyleHinterleitner, M., Hinterleitner, C., Malenke, E., Federmann, B., Holzer, U., Müller, M., Bethge, W. A., & Wirths, S. (2022). Early Reconstitution of Antibody Secreting Cells after Allogeneic Stem Cell Transplantation. Journal of Clinical Medicine, 11(1), 270. https://doi.org/10.3390/jcm11010270