
Identifying Common Pathogenic Features in Deep Endometriotic Nodules and Uterine Adenomyosis

 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Participants and Tissue Collection
2.2. Immunohistochemistry
2.3. Double Immunohistochemistry
2.4. Picrosirius Red Staining
3. Results
3.1. Clinicopathological Data
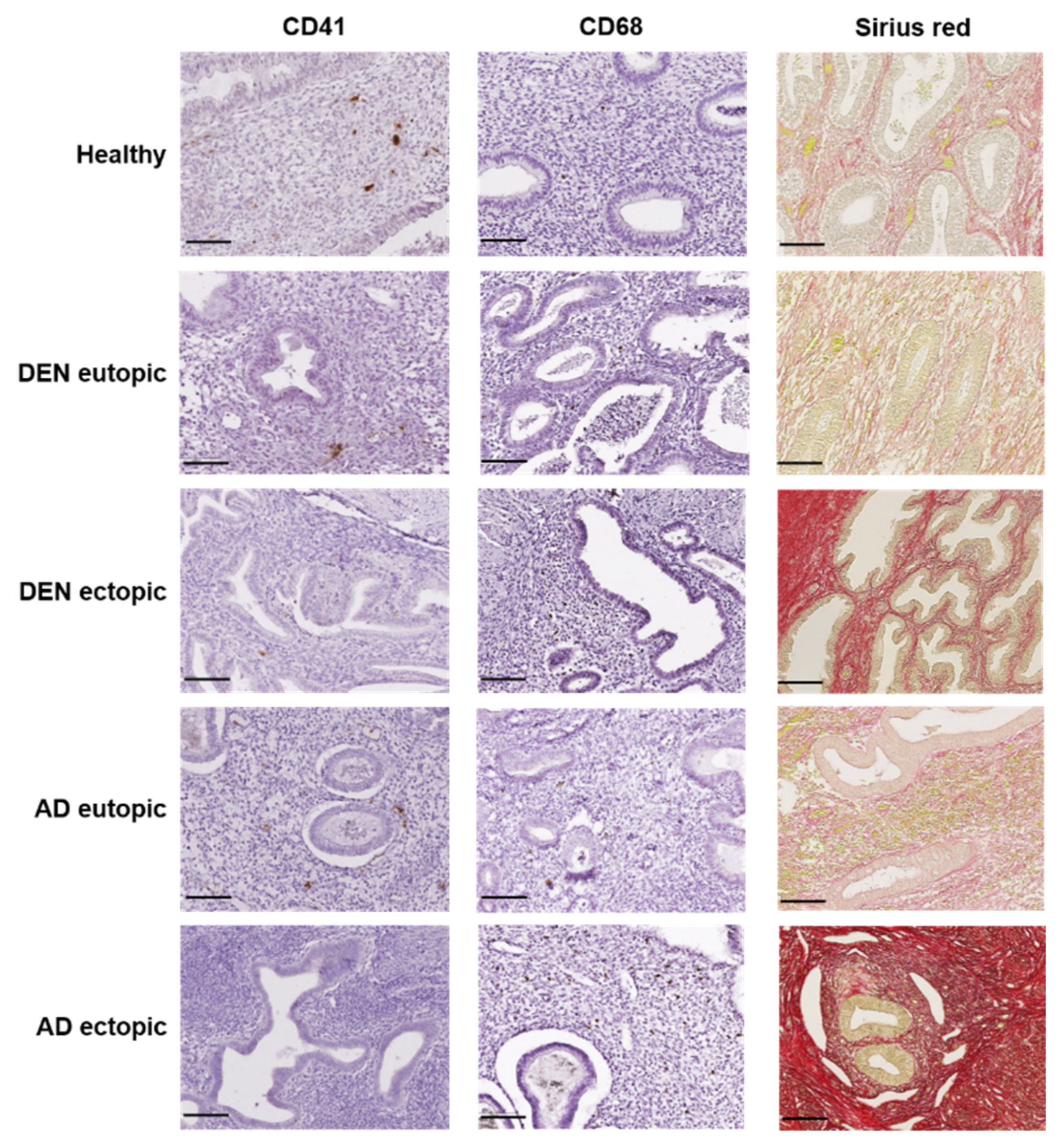
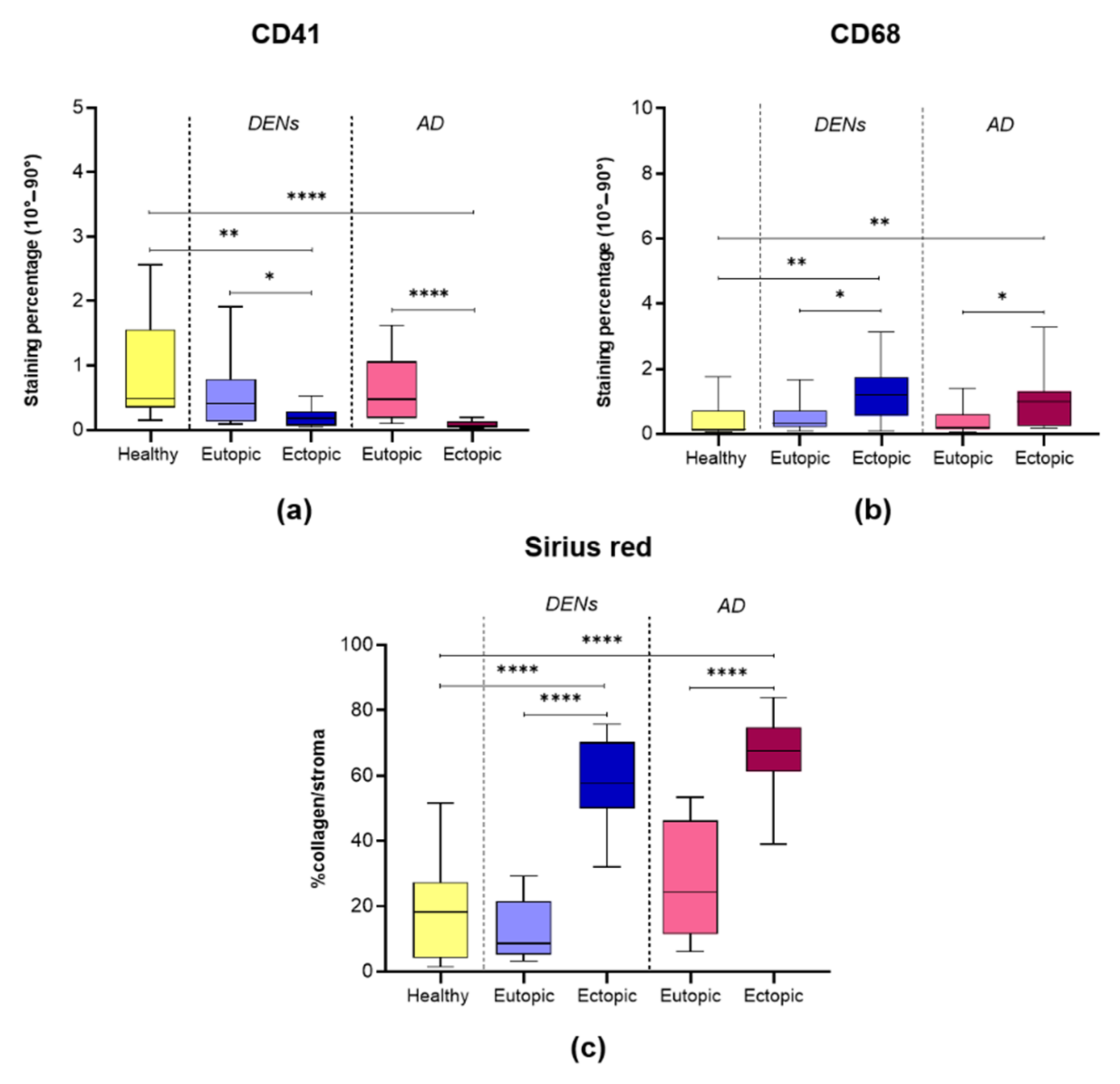
3.2. Platelet Aggregation
3.3. Macrophage Accumulation
3.4. Collagen Fiber Content
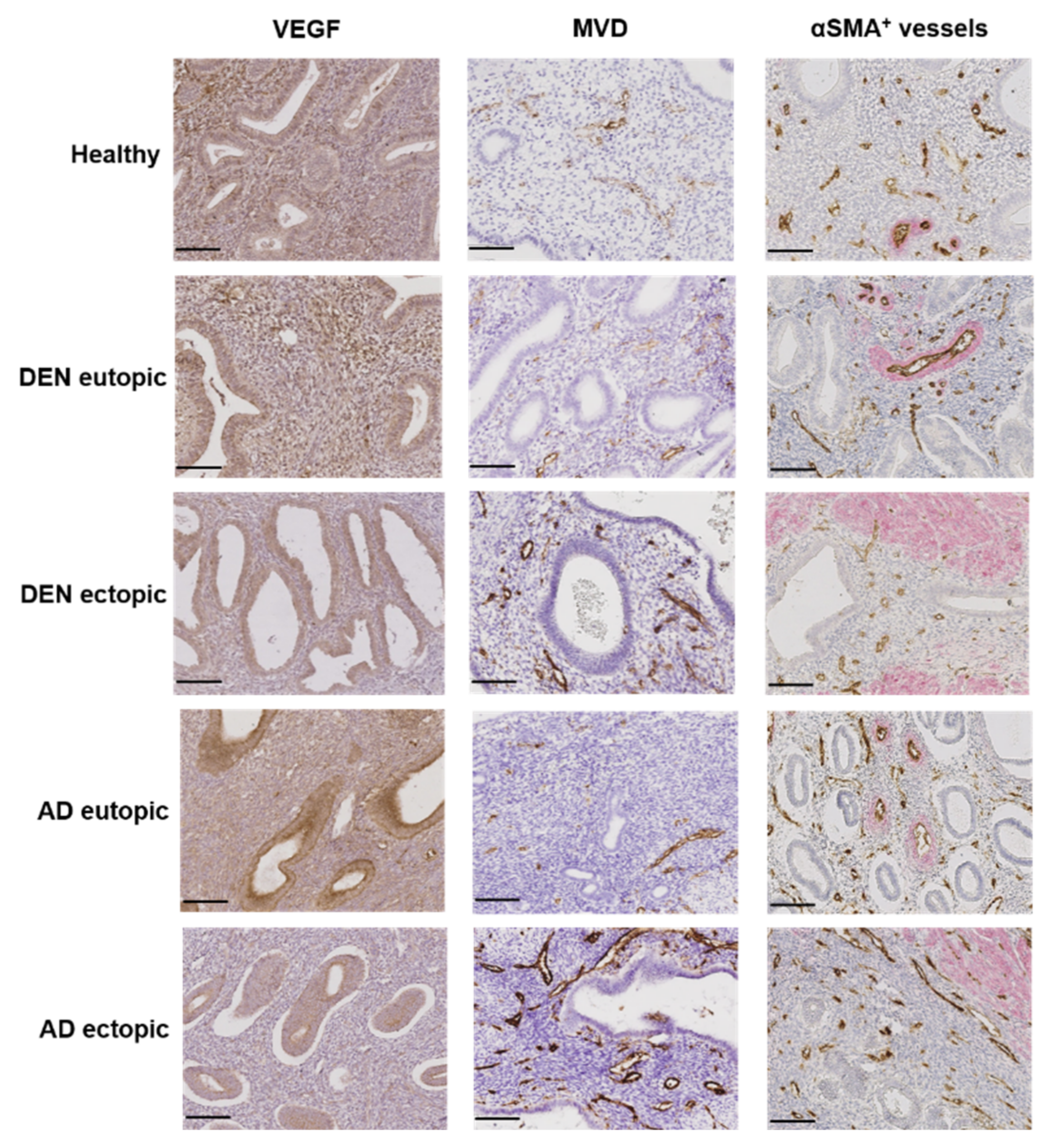
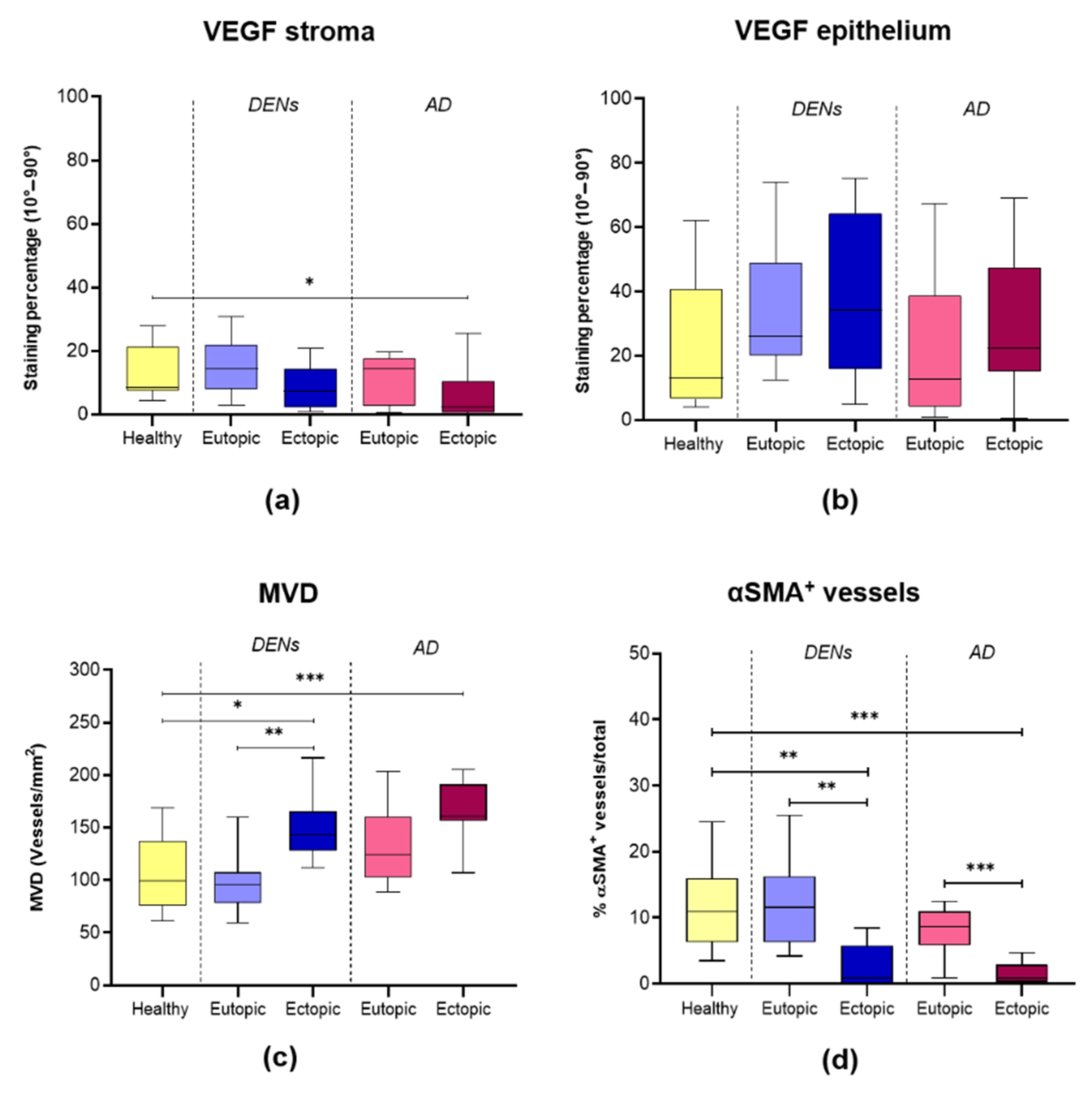
3.5. Angiogenesis and Vessel Stability
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nisolle, M.; Donnez, J. Peritoneal endometriosis, ovarian endometriosis, and adenomyotic nodules of the rectovaginal septum are three different entities. Fertil. Steril. 1997, 68, 585–596. [Google Scholar] [CrossRef]
- Giudice, L.C.; Kao, L.C. Endometriosis. Lancet 2004, 364, 1789–1799. [Google Scholar] [CrossRef]
- Taylor, H.S.; Kotlyar, A.M.; Flores, V.A. Endometriosis is a chronic systemic disease: Clinical challenges and novel innovations. Lancet 2021, 397, 839–852. [Google Scholar] [CrossRef]
- Leyendecker, G.; Bilgicyildirim, A.; Inacker, M.; Stalf, T.; Huppert, P.; Mall, G.; Böttcher, B.; Wildt, L. Adenomyosis and endometriosis. Re-visiting their association and further insights into the mechanisms of auto-traumatisation. An MRI study. Arch. Gynecol. Obstet. 2015, 291, 917–932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Solares, J.; Donnez, J.; Donnez, O.; Dolmans, M.M. Pathogenesis of uterine adenomyosis: Invagination or metaplasia? Fertil. Steril. 2018, 109, 371–379. [Google Scholar] [CrossRef]
- Donnez, J.; Dolmans, M.M.; Fellah, L. What if deep endometriotic nodules and uterine adenomyosis were actually two forms of the same disease? Fertil. Steril. 2019, 111, 454–456. [Google Scholar] [CrossRef] [Green Version]
- Stratopoulou, C.A.; Donnez, J.; Dolmans, M.M. Origin and pathogenic mechanisms of uterine adenomyosis: What is known so far. Reprod. Sci. 2021, 28, 2087–2097. [Google Scholar] [CrossRef]
- Naftalin, J.; Hoo, W.; Pateman, K.; Mavrelos, D.; Holland, T.; Jurkovic, D. How common is adenomyosis? A prospective study of prevalence using transvaginal ultrasound in a gynaecology clinic. Hum. Reprod. 2012, 27, 3432–3439. [Google Scholar] [CrossRef] [Green Version]
- Chapron, C.; Tosti, C.; Marcellin, L.; Bourdon, M.; Lafay-Pillet, M.C.; Millischer, A.E.; Streuli, I.; Borghese, B.; Petraglia, F.; Santulli, P. Relationship between the magnetic resonance imaging appearance of adenomyosis and endometriosis phenotypes. Hum. Reprod. 2017, 32, 1393–1401. [Google Scholar] [CrossRef] [Green Version]
- Proestling, K.; Birner, P.; Gamperl, S.; Nirtl, N.; Marton, E.; Yerlikaya, G.; Wenzl, R.; Streubel, B.; Husslein, H. Enhanced epithelial to mesenchymal transition (EMT) and upregulated MYC in ectopic lesions contribute independently to endometriosis. Reprod. Biol. Endocrinol. 2015, 13, 75. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.J.; Li, H.Y.; Huang, C.H.; Twu, N.F.; Yen, M.S.; Wang, P.H.; Chou, T.Y.; Liu, Y.N.; Chao, K.C.; Yang, M.H. Oestrogen-induced epithelial-mesenchymal transition of endometrial epithelial cells contributes to the development of adenomyosis. J. Pathol. 2010, 222, 261–270. [Google Scholar] [CrossRef]
- Liu, X.; Shen, M.; Qi, Q.; Zhang, H.; Guo, S.W. Corroborating evidence for platelet-induced epithelial-mesenchymal transition and fibroblast-to-myofibroblast transdifferentiation in the development of adenomyosis. Hum. Reprod. 2016, 31, 734–749. [Google Scholar] [CrossRef] [PubMed]
- An, M.; Li, D.; Yuan, M.; Li, Q.; Zhang, L.; Wang, G. Interaction of macrophages and endometrial cells induces epithelial—Mesenchymal transition-like processes in adenomyosis. Biol. Reprod. 2016, 96, 46–57. [Google Scholar]
- Maybin, J.A.; Hirani, N.; Brown, P.; Jabbour, H.N.; Critchley, H.O.D. The regulation of vascular endothelial growth factor by hypoxia and prostaglandin F2α during human endometrial repair. J. Clin. Endocrinol. Metab. 2011, 96, 2475–2483. [Google Scholar] [CrossRef] [Green Version]
- Bazot, M.; Daraï, E. Role of transvaginal sonography and magnetic resonance imaging in the diagnosis of uterine adenomyosis. Fertil. Steril. 2018, 109, 389–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levgur, M.; Abadi, M.A.; Tucker, A. Adenomyosis: Symptoms, histology, and pregnancy terminations. Obstet. Gynecol. 2000, 95, 688–691. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Marien, K.M.; Croons, V.; Waumans, Y.; Sluydts, E.; De Schepper, S.; Andries, L.; Waelput, W.; Fransen, E.; Vermeulen, P.B.; Kockx, M.M.; et al. Development and validation of a histological method to measure microvessel density in whole-slide images of cancer tissue. PLoS ONE 2016, 11, e0161496. [Google Scholar] [CrossRef]
- Martire, F.G.; Zupi, E.; Lazzeri, L.; Morosetti, G.; Conway, F.; Centini, G.; Solima, E.; Pietropolli, A.; Piccione, E.; Exacoustos, C. Transvaginal ultrasound findings after laparoscopic rectosigmoid segmental resection for deep infiltrating endometriosis. J. Ultrasound Med. 2020, 40, 1219–1228. [Google Scholar] [CrossRef] [PubMed]
- Orellana, R.; García-Solares, J.; Donnez, J.; van Kerk, O.; Dolmans, M.M.; Donnez, O. Important role of collective cell migration and nerve fiber density in the development of deep nodular endometriosis. Fertil. Steril. 2017, 107, 987–995.e5. [Google Scholar] [CrossRef] [Green Version]
- García-Solares, J.; Dolmans, M.M.; Squifflet, J.L.; Donnez, J.; Donnez, O. Invasion of human deep nodular endometriotic lesions is associated with collective cell migration and nerve development. Fertil. Steril. 2018, 110, 1318–1327. [Google Scholar] [CrossRef] [PubMed]
- Lamouille, S.; Xu, J.; Derynck, R. Molecular mechanisms of epithelial—Mesenchymal transition. Nat. Rev. Mol. Cell Biol. 2014, 15, 178–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosele, S.; Stratopoulou, C.A.; Camboni, A.; Donnez, J.; Dolmans, M.M. Investigation of the role of platelets in the aetiopathogenesis of adenomyosis. Reprod. Biomed. Online 2021, 42, 826–834. [Google Scholar] [CrossRef]
- Wynn, T.A.; Vannella, K.M. Macrophages in tissue repair, regeneration, and fibrosis. Immunity 2016, 44, 450–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capobianco, A.; Rovere-Querini, P. Endometriosis, a disease of the macrophage. Front. Immunol. 2013, 4, 9. [Google Scholar] [CrossRef] [Green Version]
- Forster, R.; Sarginson, A.; Velichkova, A.; Hogg, C.; Dorning, A.; Horne, A.W.; Saunders, P.T.K.; Greaves, E. Macrophage-derived insulin-like growth factor-1 is a key neurotrophic and nerve-sensitizing factor in pain associated with endometriosis. FASEB J. 2019, 33, 11210–11222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anaf, V.; Simon, P.; El Nakadi, I.; Fayt, I.; Simonart, T.; Buxant, F.; Noel, J.C. Hyperalgesia, nerve infiltration and nerve growth factor expression in deep adenomyotic nodules, peritoneal and ovarian endometriosis. Hum. Reprod. 2002, 17, 1895–1900. [Google Scholar] [CrossRef] [Green Version]
- Brosens, I.A. Endometriosis—A disease because it is characterized by bleeding. Am. J. Obstet. Gynecol. 1997, 176, 263–267. [Google Scholar] [CrossRef]
- Sharkey, A.M.; Day, K.; McPherson, A.; Malik, S.; Licence, D.; Smith, S.K.; Charnock-Jones, D.S. Vascular endothelial growth factor expression in human endometrium is regulated by hypoxia. J. Clin. Endocrinol. Metab. 2000, 85, 402–409. [Google Scholar] [CrossRef]
- Skalli, O.; Pelte, M.F.; Peclet, M.C.; Gabbiani, G.; Gugliotta, P.; Bussolati, G.; Ravazzola, M.; Orci, L. Alpha-smooth muscle actin, a differentiation marker of smooth muscle cells, is present in microfilamentous bundles of pericytes. J. Histochem. Cytochem. 1989, 37, 315–321. [Google Scholar] [CrossRef] [Green Version]
- Rzucidlo, E.M.; Martin, K.A.; Powell, R.J. Regulation of vascular smooth muscle cell differentiation. J. Vasc. Surg. 2007, 45, A25–A32. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, H.; Tsuruchi, N.; Sugihara, K.; Kaku, T.; Saito, T.; Kamura, T.; Tsukamoto, N.; Nakano, H.; Taniguchi, S. Expression of alpha-smooth muscle actin in benign or malignant ovarian tumors. Gynecol. Oncol. 1993, 48, 308–313. [Google Scholar] [CrossRef]
- Yonenaga, Y.; Mori, A.; Onodera, H.; Yasuda, S.; Oe, H.; Fujimoto, A.; Tachibana, T.; Imamura, M. Absence of smooth muscle actin-positive pericyte coverage of tumor vessels correlates with hematogenous metastasis and prognosis of colorectal cancer patients. Oncology 2005, 69, 159–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho-Tin-Noé, B.; Demers, M.; Wagner, D.D. How platelets safeguard vascular integrity. J. Thromb. Haemost. 2011, 9 (Suppl. 1), 56–65. [Google Scholar] [CrossRef] [PubMed]
- Leyendecker, G.; Wildt, L. A new concept of endometriosis and adenomyosis: Tissue injury and repair (TIAR). Horm. Mol. Biol. Clin. Investig. 2011, 5, 125–142. [Google Scholar] [CrossRef] [Green Version]
- Donnez, J.; Chantraine, F.; Nisolle, M. The efficacy of medical and surgical treatment of endometriosis-associated infertility: Arguments in favour of a medico-surgical approach. Hum. Reprod. Update 2002, 8, 89–94. [Google Scholar] [CrossRef] [Green Version]
- Keckstein, J.; Saridogan, E.; Ulrich, U.A.; Sillem, M.; Oppelt, P.; Schweppe, K.W.; Krentel, H.; Janschek, E.; Exacoustos, C. The #Enzian classification: A comprehensive non-invasive and surgical description system for endometriosis. Acta Obstet. Gynecol. Scand. 2021, 100, 1165–1175. [Google Scholar]
- Koninckx, P.R.; Ussia, A.; Adamyan, L.; Wattiez, A.; Donnez, J. Deep endometriosis: Definition, diagnosis, and treatment. Fertil. Steril. 2012, 98, 564–571. [Google Scholar] [CrossRef]
- Gordts, S.; Grimbizis, G.; Campo, R. Symptoms and classification of uterine adenomyosis, including the place of hysteroscopy in diagnosis. Fertil. Steril. 2018, 109, 380–388.e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, S.-W. The pathogenesis of adenomyosis vis-à-vis endometriosis. J. Clin. Med. 2020, 9, 485. [Google Scholar] [CrossRef] [Green Version]
- Maruyama, S.; Imanaka, S.; Nagayasu, M.; Kimura, M.; Kobayashi, H. Relationship between adenomyosis and endometriosis; different phenotypes of a single disease? Eur. J. Obstet. Gynecol. Reprod. Biol. 2020, 253, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Donnez, J.; Stratopoulou, C.A.; Dolmans, M.-M. Uterine adenomyosis: From disease pathogenesis to a new medical approach using GnRH antagonists. Int. J. Environ. Res. Public Health 2021, 18, 9941. [Google Scholar] [CrossRef]
- Donnez, J.; Taylor, H.S.; Taylor, R.N.; Akin, M.D.; Tatarchuk, T.F.; Wilk, K.; Gotteland, J.P.; Lecomte, V.; Bestel, E. Treatment of endometriosis-associated pain with linzagolix, an oral gonadotropin-releasing hormone—Antagonist: A randomized clinical trial. Fertil. Steril. 2020, 114, 44–55. [Google Scholar] [CrossRef]
- Donnez, O.; Donnez, J. Gonadotropin-releasing hormone antagonist (linzagolix): A new therapy for uterine adenomyosis. Fertil. Steril. 2020, 114, 640–645. [Google Scholar] [CrossRef] [PubMed]
- Vannuccini, S.; Luisi, S.; Tosti, C.; Sorbi, F.; Petraglia, F. Role of medical therapy in the management of uterine adenomyosis. Fertil. Steril. 2018, 109, 398. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Healthy (a) | Endometriosis (b) | Adenomyosis (c) | p-Value |
|---|---|---|---|---|
| Age | Mean = 41.7 | Mean = 38.7 | Mean = 45.9 | |
| SD = 8.4 | SD = 8.2 | SD = 4.8 | NS | |
| Range = 19–56 | Range = 23–51 | Range = 34–54 | ||
| Menstrual Phase Proliferative Secretory Menstrual | 4 5 5 | 6 6 1 | 5 5 4 | |
| Parity | Mean = 2 | Mean = 1 | Mean = 1.5 | |
| SD = 1.6 | SD = 0.9 | SD = 1.2 | NS | |
| Range = 0–4 | Range = 0–2 | Range = 1–4 | ||
| Menorrhagia (%) | NE | 30.8 | 85.7 | (a, b) = NS (a, c) = NS ** (b, c) = 0.006 |
| Dysmenorrhea (%) | 14.3 | 61.5 | 57.1 | * (a, b) = 0.018 * (a, c) = 0.046 (b, c) = NS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stratopoulou, C.A.; Camboni, A.; Donnez, J.; Dolmans, M.-M. Identifying Common Pathogenic Features in Deep Endometriotic Nodules and Uterine Adenomyosis. J. Clin. Med. 2021, 10, 4585. https://doi.org/10.3390/jcm10194585
Stratopoulou CA, Camboni A, Donnez J, Dolmans M-M. Identifying Common Pathogenic Features in Deep Endometriotic Nodules and Uterine Adenomyosis. Journal of Clinical Medicine. 2021; 10(19):4585. https://doi.org/10.3390/jcm10194585
Chicago/Turabian StyleStratopoulou, Christina Anna, Alessandra Camboni, Jacques Donnez, and Marie-Madeleine Dolmans. 2021. "Identifying Common Pathogenic Features in Deep Endometriotic Nodules and Uterine Adenomyosis" Journal of Clinical Medicine 10, no. 19: 4585. https://doi.org/10.3390/jcm10194585
APA StyleStratopoulou, C. A., Camboni, A., Donnez, J., & Dolmans, M.-M. (2021). Identifying Common Pathogenic Features in Deep Endometriotic Nodules and Uterine Adenomyosis. Journal of Clinical Medicine, 10(19), 4585. https://doi.org/10.3390/jcm10194585