
Dysregulated Tear Film Proteins in Macular Edema Due to the Neovascular Age-Related Macular Degeneration Are Involved in the Regulation of Protein Clearance, Inflammation, and Neovascularization


 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.2. Protein Purification and Precipitation
2.3. Preparation of Proteins for MALDI Identification
2.4. MALDI Analysis
2.5. Visual and Statistical Analysis
3. Results
4. Discussion
4.1. Oxidative Stress
4.2. Protein Clearance
4.3. Chronic Inflammation and Neovascularization
4.4. Anti-VEGF Treatment
4.5. Limitations of the Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, K.; Chen, Z.; Duan, F.; Liang, J.; Wu, K. Quantification of Tear Proteins by SDS-PAGE with an Internal Standard Protein: A New Method with Special Reference to Small Volume Tears. Graefes Arch. Clin. Exp. Ophthalmol. 2010, 248, 853–862. [Google Scholar] [CrossRef] [PubMed]
- Posa, A.; Bräuer, L.; Schicht, M.; Garreis, F.; Beileke, S.; Paulsen, F. Schirmer Strip vs. Capillary Tube Method: Non-Invasive Methods of Obtaining Proteins from Tear Fluid. Ann. Anat. 2013, 195, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Green-Church, K.B.; Nichols, K.K.; Kleinholz, N.M.; Zhang, L.; Nichols, J.J. Investigation of the Human Tear Film Proteome Using Multiple Proteomic Approaches. Mol. Vis. 2008, 14, 456–470. [Google Scholar]
- González, N.; Iloro, I.; Durán, J.A.; Elortza, F.; Suárez, T. Evaluation of Inter-Day and Inter-Individual Variability of Tear Peptide/Protein Profiles by MALDI-TOF MS Analyses. Mol. Vis. 2012, 18, 1572–1582. [Google Scholar]
- Rentka, A.; Koroskenyi, K.; Harsfalvi, J.; Szekanecz, Z.; Szucs, G.; Szodoray, P.; Kemeny-Beke, A. Evaluation of Commonly Used Tear Sampling Methods and Their Relevance in Subsequent Biochemical Analysis. Ann. Clin. Biochem. 2017, 54, 521–529. [Google Scholar] [CrossRef] [Green Version]
- Ablamowicz, A.F.; Nichols, J.J. Concentrations of MUC16 and MUC5AC Using Three Tear Collection Methods. Mol. Vis. 2017, 23, 529–537. [Google Scholar]
- Shevchenko, A.; Wilm, M.; Vorm, O.; Mann, M. Mass Spectrometric Sequencing of Proteins Silver-Stained Polyacrylamide Gels. Anal. Chem. 1996, 68, 850–858. [Google Scholar] [CrossRef]
- Winiarczyk, M.; Kaarniranta, K.; Winiarczyk, S.; Adaszek, Ł.; Winiarczyk, D.; Mackiewicz, J. Tear Film Proteome in Age-Related Macular Degeneration. Graefe’s Arch. Clin. Exp. Ophthalmol. 2018, 256, 1127–1139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tew, K.D. Redox in Redux: Emergent Roles for Glutathione S-Transferase P (GSTP) in Regulation of Cell Signaling and S-Glutathionylation. Biochem. Pharmacol. 2007, 73, 1257–1269. [Google Scholar] [CrossRef] [PubMed]
- Sreekumar, P.G.; Ferrington, D.A.; Kannan, R. Glutathione Metabolism and the Novel Role of Mitochondrial GSH in Retinal Degeneration. Antioxidants 2021, 10, 661. [Google Scholar] [CrossRef]
- Chen, W.-R.; Lan, Y.-W.; Chen, H.-L.; Chen, C.-M. AKR1A1 Deficiency Is Associated with High Risk of Alcohol-Induced Fatty Liver Syndrome. FASEB J. 2018, 32, 546.6. [Google Scholar] [CrossRef]
- Stomberski, C.T.; Anand, P.; Venetos, N.M.; Hausladen, A.; Zhou, H.-L.; Premont, R.T.; Stamler, J.S. AKR1A1 Is a Novel Mammalian S-Nitroso-Glutathione Reductase. J. Biol. Chem. 2019, 294, 18285–18293. [Google Scholar] [CrossRef]
- Morgan, C.A.; Hurley, T.D. Characterization of Two Distinct Structural Classes of Selective Aldehyde Dehydrogenase 1A1 Inhibitors. J. Med. Chem. 2015, 58, 1964–1975. [Google Scholar] [CrossRef] [Green Version]
- Kaarniranta, K.; Uusitalo, H.; Blasiak, J.; Felszeghy, S.; Kannan, R.; Kauppinen, A.; Salminen, A.; Sinha, D.; Ferrington, D. Mechanisms of Mitochondrial Dysfunction and Their Impact on Age-Related Macular Degeneration. Progress Retin. Eye Res. 2020, 100858. [Google Scholar] [CrossRef]
- Ferrington, D.A.; Sinha, D.; Kaarniranta, K. Defects in Retinal Pigment Epithelial Cell Proteolysis and the Pathology Associated with Age-Related Macular Degeneration. Progress Retin. Eye Res. 2016, 51, 69–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xi, Y.; Ju, R.; Wang, Y. Roles of Annexin A Protein Family in Autophagy Regulation and Therapy. Biomed. Pharmacother. 2020, 130, 110591. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.-F.; Huang, W.; Yi, H.-M.; Xiao, T.; Li, J.-Y.; Feng, J.; Yi, H.; Lu, S.-S.; Li, X.-H.; Lu, R.-H.; et al. Annexin A1-Suppressed Autophagy Promotes Nasopharyngeal Carcinoma Cell Invasion and Metastasis by PI3K/AKT Signaling Activation. Cell Death Dis. 2018, 9, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, I.J.; Bailey, L.M.; Aghakhani, M.R.; Moss, S.E.; Futter, C.E. EGF Stimulates Annexin 1-Dependent Inward Vesiculation in a Multivesicular Endosome Subpopulation. EMBO J. 2006, 25, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Datta, S.; Cano, M.; Ebrahimi, K.; Wang, L.; Handa, J.T. The Impact of Oxidative Stress and Inflammation on RPE Degeneration in Non-Neovascular AMD. Progress Retin. Eye Res. 2017, 60, 201–218. [Google Scholar] [CrossRef]
- Kauppinen, A.; Paterno, J.J.; Blasiak, J.; Salminen, A.; Kaarniranta, K. Inflammation and Its Role in Age-Related Macular Degeneration. Cell Mol. Life Sci. 2016, 73, 1765–1786. [Google Scholar] [CrossRef] [Green Version]
- D’Acquisto, F.; Merghani, A.; Lecona, E.; Rosignoli, G.; Raza, K.; Buckley, C.D.; Flower, R.J.; Perretti, M. Annexin-1 Modulates T-Cell Activation and Differentiation. Blood 2007, 109, 1095–1102. [Google Scholar] [CrossRef] [Green Version]
- Sanches, J.M.; Correia-Silva, R.D.; Duarte, G.H.B.; Fernandes, A.M.A.P.; Sánchez-Vinces, S.; Carvalho, P.O.; Oliani, S.M.; Bortoluci, K.R.; Moreira, V.; Gil, C.D. Role of Annexin A1 in NLRP3 Inflammasome Activation in Murine Neutrophils. Cells 2021, 10, 121. [Google Scholar] [CrossRef] [PubMed]
- Yazid, S.; Gardner, P.J.; Carvalho, L.; Chu, C.J.; Flower, R.J.; Solito, E.; Lee, R.W.J.; Ali, R.R.; Dick, A.D. Annexin-A1 Restricts Th17 Cells and Attenuates the Severity of Autoimmune Disease. J. Autoimmun. 2015, 58, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kang, G.-Y.; Bang, J.Y.; Choi, A.J.; Yoon, J.; Lee, W.-C.; Choi, S.; Yoon, S.; Kim, H.C.; Baek, J.-H.; Park, H.S.; et al. Exosomal Proteins in the Aqueous Humor as Novel Biomarkers in Patients with Neovascular Age-Related Macular Degeneration. J. Proteome Res. 2014, 13, 581–595. [Google Scholar] [CrossRef] [PubMed]
- Crabb, J.W.; Miyagi, M.; Gu, X.; Shadrach, K.; West, K.A.; Sakaguchi, H.; Kamei, M.; Hasan, A.; Yan, L.; Rayborn, M.E.; et al. Drusen Proteome Analysis: An Approach to the Etiology of Age-Related Macular Degeneration. Proc. Natl. Acad. Sci. USA 2002, 99, 14682–14687. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Quigley, H.A.; Pease, M.E.; Yang, Y.; Qian, J.; Valenta, D.; Zack, D.J. Changes in Gene Expression in Experimental Glaucoma and Optic Nerve Transection: The Equilibrium between Protective and Detrimental Mechanisms. Investig. Ophthalmol. Vis. Sci. 2007, 48, 5539–5548. [Google Scholar] [CrossRef] [PubMed]
- Iaccarino, L.; Ghirardello, A.; Canova, M.; Zen, M.; Bettio, S.; Nalotto, L.; Punzi, L.; Doria, A. Anti-Annexins Autoantibodies: Their Role as Biomarkers of Autoimmune Diseases. Autoimmun. Rev. 2011, 10, 553–558. [Google Scholar] [CrossRef]
- Cañas, F.; Simonin, L.; Couturaud, F.; Renaudineau, Y. Annexin A2 Autoantibodies in Thrombosis and Autoimmune Diseases. Thromb. Res. 2015, 135, 226–230. [Google Scholar] [CrossRef]
- Adamus, G.; Chew, E.Y.; Ferris, F.L.; Klein, M.L. Prevalence of Anti-Retinal Autoantibodies in Different Stages of Age-Related Macular Degeneration. BMC Ophthalmol. 2014, 14, 154. [Google Scholar] [CrossRef] [Green Version]
- Joachim, S.C.; Bruns, K.; Lackner, K.J.; Pfeiffer, N.; Grus, F.H. Analysis of IgG Antibody Patterns against Retinal Antigens and Antibodies to α-Crystallin, GFAP, and α-Enolase in Sera of Patients with “Wet” Age-Related Macular Degeneration. Graefe’s Arch. Clin. Exp. Ophthalmol. 2006, 245, 619. [Google Scholar] [CrossRef]
- Adamus, G.; Aptsiauri, N.; Guy, J.; Heckenlively, J.; Flannery, J.; Hargrave, P.A. The Occurrence of Serum Autoantibodies against Enolase in Cancer-Associated Retinopathy. Clin. Immunol. Immunopathol. 1996, 78, 120–129. [Google Scholar] [CrossRef]
- Dot, C.; Guigay, J.; Adamus, G. Anti-α-Enolase Antibodies in Cancer-Associated Retinopathy with Small Cell Carcinoma of the Lung. Am. J. Ophthalmol. 2005, 139, 746–747. [Google Scholar] [CrossRef] [PubMed]
- Morohoshi, K.; Goodwin, A.M.; Ohbayashi, M.; Ono, S.J. Autoimmunity in Retinal Degeneration: Autoimmune Retinopathy and Age-Related Macular Degeneration. J. Autoimmun. 2009, 33, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Hikage, F.; Lennikov, A.; Mukwaya, A.; Lachota, M.; Ida, Y.; Utheim, T.P.; Chen, D.F.; Huang, H.; Ohguro, H. NF-ΚB Activation in Retinal Microglia Is Involved in the Inflammatory and Neovascularization Signaling in Laser-Induced Choroidal Neovascularization in Mice. Exp. Cell Res. 2021, 403, 112581. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.K.; Occelli, L.M.; Binette, F.; Petersen-Jones, S.M.; Nasonkin, I.O. Transplantation of Human Embryonic Stem Cell-Derived Retinal Tissue in the Subretinal Space of the Cat Eye. Stem Cells Dev. 2019, 28, 1151–1166. [Google Scholar] [CrossRef] [PubMed]
- Hambright, D.; Park, K.-Y.; Brooks, M.; McKay, R.; Swaroop, A.; Nasonkin, I.O. Long-Term Survival and Differentiation of Retinal Neurons Derived from Human Embryonic Stem Cell Lines in Un-Immunosuppressed Mouse Retina. Mol. Vis. 2012, 18, 920–936. [Google Scholar]
- Stifter, J.; Ulbrich, F.; Goebel, U.; Böhringer, D.; Lagrèze, W.A.; Biermann, J. Neuroprotection and Neuroregeneration of Retinal Ganglion Cells after Intravitreal Carbon Monoxide Release. PLoS ONE 2017, 12, e0188444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ambati, J.; Fowler, B.J. Mechanisms of Age-Related Macular Degeneration. Neuron 2012, 75, 26–39. [Google Scholar] [CrossRef] [Green Version]
- Gutmann, D.A.P.; Ward, A.; Urbatsch, I.L.; Chang, G.; van Veen, H.W. Understanding Polyspecificity of Multidrug ABC Transporters: Closing in on the Gaps in ABCB1. Trends Biochem. Sci. 2010, 35, 36–42. [Google Scholar] [CrossRef] [Green Version]
- Hodges, L.M.; Markova, S.M.; Chinn, L.W.; Gow, J.M.; Kroetz, D.L.; Klein, T.E.; Altman, R.B. Very Important Pharmacogene Summary: ABCB1 (MDR1, P-Glycoprotein). Pharmacogenet Genom. 2011, 21, 152–161. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.-K.; Frank, T. Teaching an Old Dog New Tricks: Reactivated Developmental Signaling Pathways Regulate ABCB1 and Chemoresistance in Cancer. Cancer Drug Resist. 2021, 4. [Google Scholar] [CrossRef]
- Eastlake, K.; Heywood, W.E.; Banerjee, P.; Bliss, E.; Mills, K.; Khaw, P.T.; Charteris, D.; Limb, G.A. Comparative Proteomic Analysis of Normal and Gliotic PVR Retina and Contribution of Müller Glia to This Profile. Exp. Eye Res. 2018, 177, 197–207. [Google Scholar] [CrossRef]
- Mirzaei, M.; Pushpitha, K.; Deng, L.; Chitranshi, N.; Gupta, V.; Rajput, R.; Mangani, A.B.; Dheer, Y.; Godinez, A.; McKay, M.J.; et al. Upregulation of Proteolytic Pathways and Altered Protein Biosynthesis Underlie Retinal Pathology in a Mouse Model of Alzheimer’s Disease. Mol. Neurobiol. 2019, 56, 6017–6034. [Google Scholar] [CrossRef] [PubMed]
- Ni, S.; Ye, M.; Huang, T. Short Stature Homeobox 2 Methylation as a Potential Noninvasive Biomarker in Bronchial Aspirates for Lung Cancer Diagnosis. Oncotarget 2017, 8, 61253–61263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ang, W.J.; Zunaina, E.; Norfadzillah, A.J.; Raja-Norliza, R.O.; Julieana, M.; Ab-Hamid, S.A.; Mahaneem, M. Evaluation of Vascular Endothelial Growth Factor Levels in Tears and Serum among Diabetic Patients. PLoS ONE 2019, 14. [Google Scholar] [CrossRef] [PubMed]
- Rentka, A.; Hársfalvi, J.; Berta, A.; Köröskényi, K.; Szekanecz, Z.; Szücs, G.; Szodoray, P.; Kemény-Beke, Á. Vascular Endothelial Growth Factor in Tear Samples of Patients with Systemic Sclerosis. Mediators Inflamm. 2015, 2015. [Google Scholar] [CrossRef] [PubMed]
- Klaassen, I.; Van Noorden, C.J.F.; Schlingemann, R.O. Molecular Basis of the Inner Blood-Retinal Barrier and Its Breakdown in Diabetic Macular Edema and Other Pathological Conditions. Progress Retin. Eye Res. 2013, 34, 19–48. [Google Scholar] [CrossRef] [PubMed]
- Schlingemann, R.O. Role of Growth Factors and the Wound Healing Response in Age-Related Macular Degeneration. Graefes Arch. Clin. Exp. Ophthalmol. 2004, 242, 91–101. [Google Scholar] [CrossRef]
- El-Mollayess, G.M.; Noureddine, B.N.; Bashshur, Z.F. Bevacizumab and Neovascular Age Related Macular Degeneration: Pathogenesis and Treatment. Semin. Ophthalmol. 2011, 26, 69–76. [Google Scholar] [CrossRef]
- Gawęcki, M.; Jaszczuk-Maciejewska, A.; Jurska-Jaśko, A.; Kneba, M.; Grzybowski, A. Transfoveal Micropulse Laser Treatment of Central Serous Chorioretinopathy within Six Months of Disease Onset. J. Clin. Med. 2019, 8, 1398. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
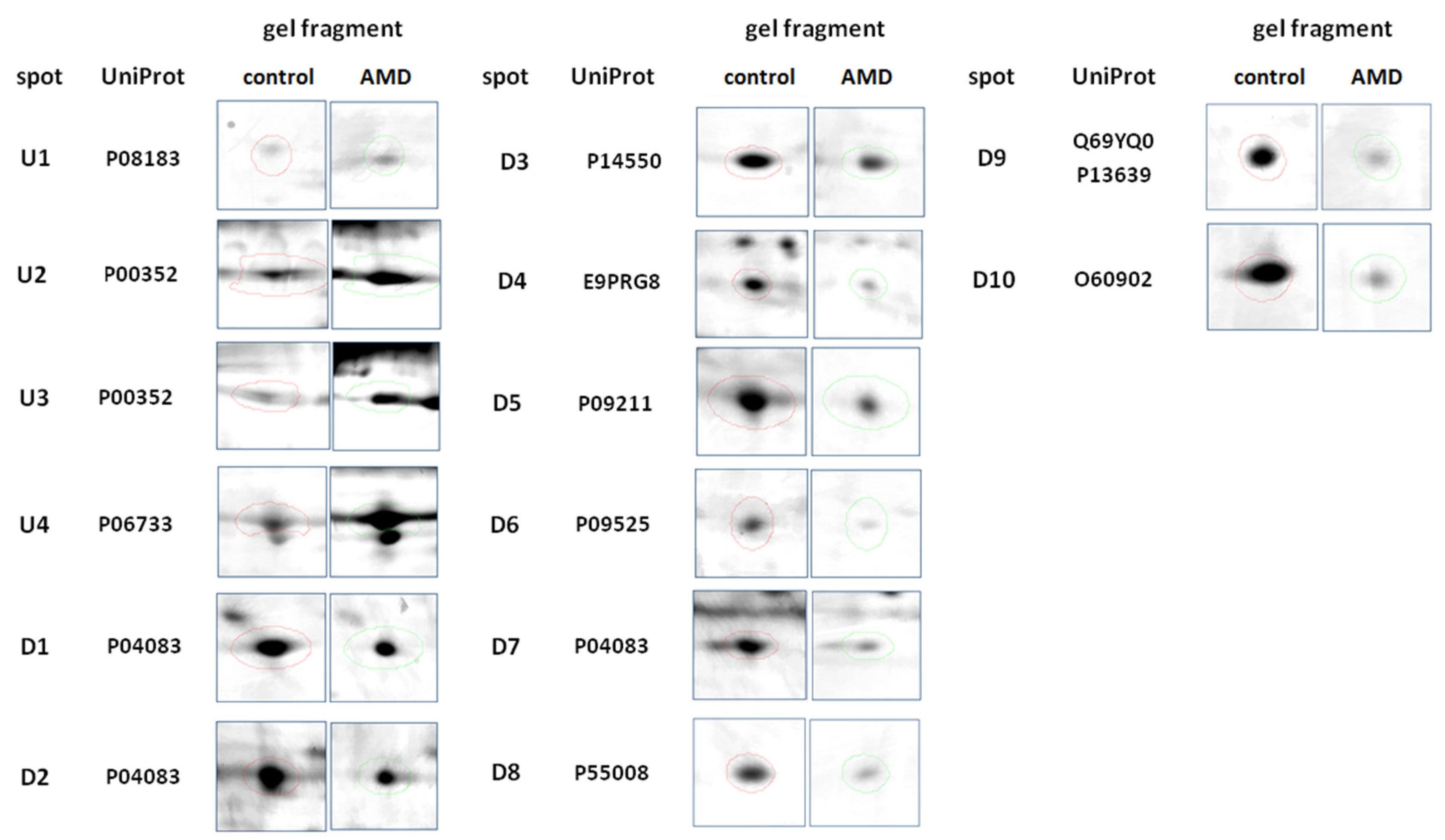
| ID | Protein | Accession Number (UniProtKB) | Species | Score | Match | MW (Da) | pI | Seq. Cov (%) | Rt | p-Value |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | ATP-dependent translocase ABCB1 | P08183 | H. sapiens | 87 | 11 | 141,788 | 9.06 | 9 | 2.193 | 0.025 |
| 5 | Annexin A1 | P04083 | H. sapiens | 96 | 12 | 38,918 | 6.57 | 41 | 0.664 | 0.026 |
| 6 | Annexin A1 | P04083 | H. sapiens | 59 | 9 | 38,918 | 6.57 | 30 | 0.575 | 0.017 |
| 8 | Aldo-keto reductase family 1 member A1 | P14550 | H. sapiens | 146 | 15 | 36,892 | 6.32 | 48 | 0.638 | 0.029 |
| 10 | Retinal dehydrogenase 1 | P00352 | H. sapiens | 75 | 9 | 55,454 | 6.30 | 24 | 2.027 | 0.011 |
| 12 | Uncharacterized protein C11orf98 | E9PRG8 | H. sapiens | 76 | 5 | 14,225 | 11.53 | 38 | 0.560 | 0.008 |
| 15 | Glutathione S-transferase P | P09211 | H. sapiens | 89 | 8 | 23,569 | 5.43 | 50 | 0.529 | 0.007 |
| 23 | Retinal dehydrogenase 1 | P00352 | H. sapiens | 121 | 14 | 55,454 | 6.30 | 41 | 1.991 | 0.015 |
| 24 | Alpha-enolase | P06733 | H. sapiens | 67 | 11 | 47,481 | 7.01 | 29 | 1.476 | 0.022 |
| 11 | Annexin A4 | P09525 | H. sapiens | 94 | 14 | 36,088 | 5.84 | 14 | 0.393 | 0.003 |
| 21 | Annexin A1 | P04083 | H. sapiens | 76 | 11 | 38,918 | 6.57 | 35 | 0.213 | 0.008 |
| 31 | Allograft inflammatory factor 1 | P55008 | H. sapiens | 77 | 5 | 16,693 | 5.97 | 34 | 0.560 | 0.026 |
| 33 | Cytospin-A or Elongation factor 2 | Q69YQ0 P13639 | H. sapiens | 113 88 | 16 12 | 124,925 96,246 | 5.52 6.41 | 16 12 | 0.560 | 0.037 |
| 32 | Short stature homeobox protein 2 | O60902 | H. sapiens | 65 | 5 | 35,160 | 8.99 | 12 | 0.529 | 0.041 |
| Identified Protein | Upregulation or Downregulation | Fold Relative to Healthy Controls | Standard Deviation (SD) |
|---|---|---|---|
| Retinal dehydrogenase 1 | Up | 2.072 1.991 | 0.011 0.015 |
| ATP-dependent translocase ABCB1 | Up | 2.193 | 0.025 |
| Alpha-enolase | Up | 1.476 | 0.022 |
| Annexin A1 | Down | 0.664 0.575 0.213 | 0.026 0.017 0.008 |
| Annexin A4 | Down | 0.393 | 0.003 |
| Aldo-keto reductase family 1 member A1 | Down | 0.638 | 0.029 |
| Uncharacterized protein C11orf98 | Down | 0.560 | 0.008 |
| Glutathione S-transferase P | Down | 0.529 | 0.007 |
| Allograft inflammatory factor 1 | Down | 0.560 | 0.026 |
| Cytospin-A or Elongation factor 2 | Down | 0.560 | 0.037 |
| Short stature homeobox protein 2 | Down | 0.529 | 0.041 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Winiarczyk, M.; Winiarczyk, D.; Michalak, K.; Kaarniranta, K.; Adaszek, Ł.; Winiarczyk, S.; Mackiewicz, J. Dysregulated Tear Film Proteins in Macular Edema Due to the Neovascular Age-Related Macular Degeneration Are Involved in the Regulation of Protein Clearance, Inflammation, and Neovascularization. J. Clin. Med. 2021, 10, 3060. https://doi.org/10.3390/jcm10143060
Winiarczyk M, Winiarczyk D, Michalak K, Kaarniranta K, Adaszek Ł, Winiarczyk S, Mackiewicz J. Dysregulated Tear Film Proteins in Macular Edema Due to the Neovascular Age-Related Macular Degeneration Are Involved in the Regulation of Protein Clearance, Inflammation, and Neovascularization. Journal of Clinical Medicine. 2021; 10(14):3060. https://doi.org/10.3390/jcm10143060
Chicago/Turabian StyleWiniarczyk, Mateusz, Dagmara Winiarczyk, Katarzyna Michalak, Kai Kaarniranta, Łukasz Adaszek, Stanisław Winiarczyk, and Jerzy Mackiewicz. 2021. "Dysregulated Tear Film Proteins in Macular Edema Due to the Neovascular Age-Related Macular Degeneration Are Involved in the Regulation of Protein Clearance, Inflammation, and Neovascularization" Journal of Clinical Medicine 10, no. 14: 3060. https://doi.org/10.3390/jcm10143060
APA StyleWiniarczyk, M., Winiarczyk, D., Michalak, K., Kaarniranta, K., Adaszek, Ł., Winiarczyk, S., & Mackiewicz, J. (2021). Dysregulated Tear Film Proteins in Macular Edema Due to the Neovascular Age-Related Macular Degeneration Are Involved in the Regulation of Protein Clearance, Inflammation, and Neovascularization. Journal of Clinical Medicine, 10(14), 3060. https://doi.org/10.3390/jcm10143060