
Clinical Relevance of Torque Teno Virus (TTV) in HIV/HCV Coinfected and HCV Monoinfected Patients Treated with Direct-Acting Antiviral Therapy

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Population and Sample Collection
2.1.1. HCV RNA Detection
2.1.2. Real-Time Quantitation of TTV DNA Load
2.2. TTV DNA Amplification and Phylogenetic Analysis
2.3. Quantification of Cytokines in Sera
2.4. Statistical Analysis
3. Results
3.1. Virological and Immunological Results
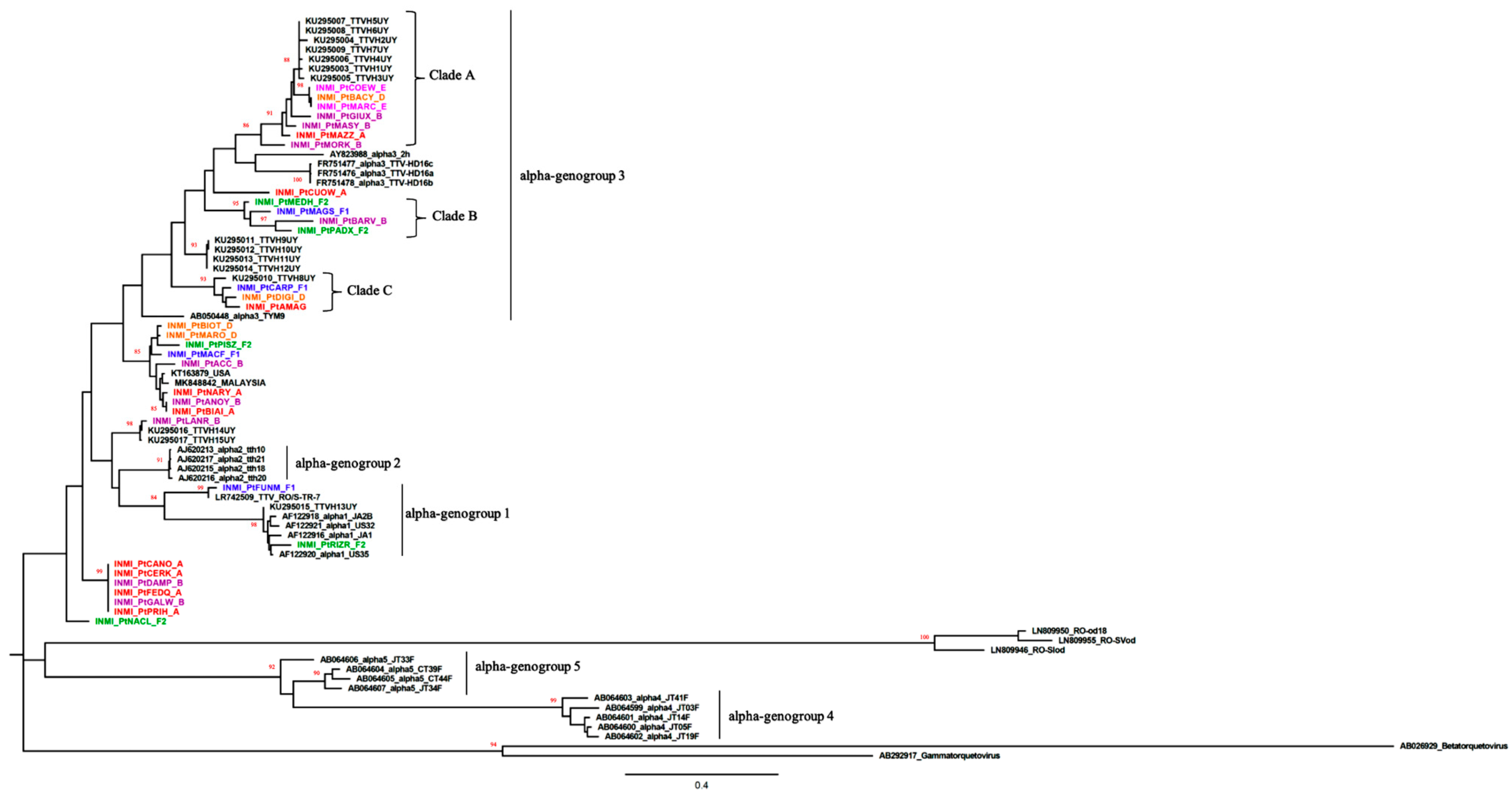
3.2. Phylogenetic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nishizawa, T.; Okamoto, H.; Konishi, K.; Yoshizawa, H.; Miyakawa, Y.; Mayumi, M. A novel DNA virus (TTV) associated with elevated transaminase levels in posttransfusion hepatitis of unknown etiology. Biochem. Biophys. Res. Commun. 1997, 241, 92–97. [Google Scholar] [CrossRef]
- Okamoto, H. History of discoveries and pathogenicity of TT viruses. Curr. Top. Microbiol. Immunol. 2009, 331, 1–20. [Google Scholar]
- Webb, B.; Rakibuzzaman, A.; Ramamoorthy, S. Torque teno viruses in health and disease. Virus Res. 2020, 285, 198013. [Google Scholar] [CrossRef]
- Biagini, P. Classification of TTV and related viruses (anelloviruses). Curr. Top. Microbiol. Immunol. 2009, 331, 21–33. [Google Scholar] [PubMed]
- Hsiao, K.L.; Wang, L.Y.; Lin, C.L.; Liu, H.F. New Phylogenetic Groups of Torque Teno Virus Identified in Eastern Taiwan Indigenes. PLoS ONE 2016, 11, e0149901. [Google Scholar] [CrossRef] [PubMed]
- De Vlaminck, I.; Khush, K.K.; Strehl, C.; Kohli, B.; Luikart, H.; Neff, N.F.; Okamoto, J.; Snyder, T.M.; Cornfield, D.N.; Nicolls, M.R.; et al. Temporal response of the human virome to immunosuppression and antiviral therapy. Cell 2013, 155, 1178–1187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poovorawan, Y.; Tangkijvanich, P.; Theamboonlers, A.; Hirsch, P. Transfusion transmissible virus TTV and its putative role in the etiology of liver disease. Hepatogastroenterology 2001, 48, 256–260. [Google Scholar] [PubMed]
- Okamoto, H.; Takahashi, M.; Nishizawa, T.; Ukita, M.; Fukuda, M.; Tsuda, F.; Miyakawa, Y.; Mayumi, M. Marked genomic heterogeneity and frequent mixed infection of TT virus demonstrated by PCR with primers from coding and noncoding regions. Virology 1999, 259, 428–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szládek, G.; Juhász, A.; Kardos, G.; Szoke, K.; Major, T.; Sziklai, I.; Tar, I.; Márton, I.; Kónya, J.; Gergely, L.; et al. High co-prevalence of genogroup 1 TT virus and human papillomavirus is associated with poor clinical outcome of laryngeal carcinoma. J. Clin. Pathol. 2005, 58, 402–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garbuglia, A.R.; Iezzi, T.; Capobianchi, M.R.; Pignoloni, P.; Pulsoni, A.; Sourdis, J.; Pescarmona, E.; Vitolo, D.; Mandelli, F. Detection of TT virus in lymph node biopsies of B-cell lymphoma and Hodgkin’s disease, and its association with EBV infection. Int. J. Immunopathol. Pharmacol. 2003, 16, 109–118. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, F.; Shan, T.L.; Deng, X.; Delwart, E.; Feng, X.P. A novel species of torque teno mini virus (TTMV) in gingival tissue from chronic periodontitis patients. Sci. Rep. 2016, 6, 26739. [Google Scholar] [CrossRef] [Green Version]
- Ng, T.F.F.; Dill, J.A.; Camus, A.C.; Delwart, E.; Van Meir, E.G. Two new species of betatorqueviruses identified in a human melanoma that metastasized to the brain. Oncotarget 2017, 8, 105800–105808. [Google Scholar] [CrossRef]
- Bal, A.; Oriol, G.; Josset, L.; Generenaz, L.; Sarkozy, C.; Sesques, P.; Salles, G.; Morfin, F.; Lina, B.; Becker, J.; et al. Metagenomic Investigation of Torque Teno Mini Virus-SH in Hematological Patients. Front. Microbiol. 2019, 10, 1898. [Google Scholar] [CrossRef]
- Spandole-Dinu, S.; Cimponeriu, D.G.; Crăciun, A.M.; Radu, I.; Nica, S.; Toma, M.; Alexiu, O.A.; Iorga, C.S.; Berca, L.M.; Nica, R. Prevalence of human anelloviruses in Romanian healthy subjects and patients with common pathologies. BMC Infect. Dis. 2018, 18, 334. [Google Scholar] [CrossRef] [PubMed]
- Rivanera, D.; Lozzi, M.A.; Idili, C.; Lilli, D. Prevalence of TT virus infection in Italian dialysis patients. Pathol. Biol. (Paris) 2009, 57, 97–100. [Google Scholar] [CrossRef]
- Irshad, M.; Mandal, K.; Singh, S.; Agarwal, S.K. Torque teno virus infection in hemodialysis patients in North India. Int. Urol. Nephrol. 2010, 42, 1077–1083. [Google Scholar] [CrossRef] [PubMed]
- Massaú, A.; Martins, C.; Nachtigal, G.C.; Araújo, A.B.; Rossetti, M.L.; Niel, C.; da Silva, C.M. The high prevalence of Torque teno virus DNA in blood donors and haemodialysis patients in southern Brazil. Mem. Inst. Oswaldo Cruz 2012, 107, 684–686. [Google Scholar] [CrossRef] [PubMed]
- Ataei, B.; Emami Naeini, A.; Khorvash, F.; Yazdani, M.R.; Javadi, A.A. Prevalence of transfusion transmitted virus infection in hemodialysis patients and injection drug users compared to healthy blood donors in isfahan, iran. Gastroenterol. Res. Pract. 2012, 2012, 671927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okamoto, H.; Nishizawa, T.; Kato, N.; Ukita, M.; Ikeda, K.; Iizuka, H.; Miyakawa, Y.; Mayumi, M. Molecular cloning and characterization of a novel DNA virus (TTV) associated with posttransfusion hepatitis of unknow etiology. Hepatol. Res. 1998, 10, 1–16. [Google Scholar] [CrossRef]
- Asim, M.; Singla, R.; Gupta, R.K.; Kar, P. Clinical & molecular characterization of human TT virus in different liver diseases. Indian J. Med. Res. 2010, 131, 545–554. [Google Scholar]
- Matsumoto, A.; Yeo, A.E.; Shih, J.W.; Tanaka, E.; Kiyosawa, K.; Alter, H.J. Transfusion-associated TT virus infection and its relationship to liver disease. Hepatology 1999, 30, 283–288. [Google Scholar] [CrossRef] [PubMed]
- Kato, H.; Mizokami, M.; Orito, E.; Ohno, T.; Hayashi, K.; Nakano, T.; Kato, T.; Tanaka, Y.; Sugauchi, F.; Mukaide, M.; et al. Lack of association between TTV viral load and aminotransferase levels in patients with hepatitis C or non-B-C. Scand. J. Infect. Dis. 2000, 32, 259–262. [Google Scholar]
- Irshad, M.; Sharma, Y.; Dhar, I.; Singh, J.; Joshi, Y.K. Transfusion-transmitted virus in association with hepatitis A-E viral infections in various forms of liver diseases in India. World J. Gastroenterol. 2006, 12, 2432–2436. [Google Scholar] [CrossRef] [PubMed]
- Focosi, D.; Antonelli, G.; Pistello, M.; Maggi, F. Torquetenovirus: The human virome from bench to bedside. Clin. Microbiol. Infect. 2016, 22, 589–593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maggi, F.; Focosi, D.; Albani, M.; Lanini, L.; Vatteroni, M.L.; Petrini, M.; Ceccherini-Nelli, L.; Pistello, M.; Bendinelli, M. Role of hematopoietic cells in the maintenance of chronic human torquetenovirus plasma viremia. J. Virol. 2010, 84, 6891–6893. [Google Scholar] [CrossRef] [Green Version]
- Zheng, H.; Ye, L.; Fang, X.; Li, B.; Wang, Y.; Xiang, X.; Kong, L.; Wang, W.; Zeng, Y.; Ye, L.; et al. Torque teno virus (SANBAN isolate) ORF2 protein suppresses NF-kappaB pathways via interaction with IkappaB kinases. J. Virol. 2007, 81, 11917–11924. [Google Scholar] [CrossRef] [Green Version]
- Shibayama, T.; Masuda, G.; Ajisawa, A.; Takahashi, M.; Nishizawa, T.; Tsuda, F.; Okamoto, H. Inverse relationship between the titre of TT virus DNA and the CD4 cell count in patients infected with HIV. Aids 2001, 15, 563–570. [Google Scholar] [CrossRef] [Green Version]
- Thom, K.; Petrik, J. Progression towards AIDS leads to increased Torque teno virus and Torque teno minivirus titers in tissues of HIV infected individuals. J. Med. Virol. 2007, 79, 1–7. [Google Scholar] [CrossRef]
- Nasser, T.F.; Brajão de Oliveira, K.; Reiche, E.M.; Amarante, M.K.; Pelegrinelli Fungaro, M.H.; Watanabe, M.A. Detection of TT virus in HIV-1 exposed but uninfected individuals and in HIV-1 infected patients and its influence on CD4+ lymphocytes and viral load. Microb. Pathog. 2009, 47, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Elesinnla, A.R.; Adeleye, I.A.; Ayolabi, C.I.; Bessong, P.O. Prevalence of torque viruses in HIV-infected and non-HIV-infected Nigerian subjects: Analysis of near-full-length genome sequences. Arch. Virol. 2020, 165, 571–582. [Google Scholar] [CrossRef]
- Devalle, S.; Rua, F.; Morgado, M.G.; Niel, C. Variations in the frequencies of torque teno virus subpopulations during HAART treatment in HIV-1-coinfected patients. Arch. Virol. 2009, 154, 1285–1291. [Google Scholar] [CrossRef]
- Reshetnyak, V.I.; Maev, I.V.; Burmistrov, A.I.; Chekmazov, I.A.; Karlovich, T.I. Torque teno virus in liver diseases: On the way towards unity of view. World J. Gastroenterol. 2020, 26, 1691–1707. [Google Scholar] [CrossRef] [PubMed]
- Zur Hausen, H.; de Villiers, E.M. TT viruses: Oncogenic or tumor-suppressive properties? Curr. Top. Microbiol. Immunol. 2009, 331, 109–116. [Google Scholar]
- Wyles, D.L.; Sulkowski, M.S.; Dieterich, D. Management of Hepatitis C/HIV Coinfection in the Era of Highly Effective Hepatitis C Virus Direct-Acting Antiviral Therapy. Clin. Infect. Dis. 2016, 63 (Suppl. 1), S3–S11. [Google Scholar] [CrossRef] [PubMed]
- Pereira, G.L.; Tarragô, A.M.; Neves, W.L.L.; da Silva Neto, P.V.; de Souza, P.S.; Dos Santos Affonso, J.; de Sousa, K.S.; da Silva, J.A.; Costa, A.G.; da Silva Victoria, F.; et al. Immunological Dynamics Associated with Direct-Acting Antiviral Therapies in Naive and Experimented HCV Chronic-Infected Patients. Mediat. Inflamm. 2019, 2019, 4738237. [Google Scholar] [CrossRef] [PubMed]
- Pericot-Valverde, I.; Heo, M.; Akiyama, M.J.; Norton, B.L.; Agyemang, L.; Niu, J.; Litwin, A.H. Factors and HCV treatment outcomes associated with smoking among people who inject drugs on opioid agonist treatment: Secondary analysis of the PREVAIL randomized clinical trial. BMC Infect. Dis. 2020, 20, 928. [Google Scholar] [CrossRef] [PubMed]
- Werner, C.R.; Schwarz, J.M.; Egetemeyr, D.P.; Beck, R.; Malek, N.P.; Lauer, U.M.; Berg, C.P. Second-generation direct-acting-antiviral hepatitis C virus treatment: Efficacy, safety, and predictors of SVR12. World J. Gastroenterol. 2016, 22, 8050–8059. [Google Scholar] [CrossRef] [PubMed]
- Visco-Comandini, U.; Lapa, D.; Lionetti, R.; Taibi, C.; Loiacono, L.; Montalbano, M.; Capobianchi, M.R.; D’Offizi, G.; Garbuglia, A.R. Significance of detectable HCV RNA below the limit of quantification in patients treated with DAAs using standard and ultrasensitive protocols. J. Med. Virol. 2018, 90, 1264–1271. [Google Scholar] [CrossRef] [PubMed]
- Kulifaj, D.; Durgueil-Lariviere, B.; Meynier, F.; Munteanu, E.; Pichon, N.; Dubé, M.; Joannes, M.; Essig, M.; Hantz, S.; Barranger, C.; et al. Development of a standardized real time PCR for Torque teno viruses (TTV) viral load detection and quantification: A new tool for immune monitoring. J. Clin. Virol. 2018, 105, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Kulifaj, D.; Tilloy, V.; Scaon, E.; Guerin, E.; Essig, M.; Pichon, N.; Hantz, S.; De Bernardi, A.; Joannes, M.; Barranger, C.; et al. Viral metagenomics analysis of kidney donors and recipients: Torque teno virus genotyping and prevalence. J. Med. Virol. 2020, 92, 3301–3311. [Google Scholar] [CrossRef]
- Cancela, F.; Ramos, N.; Mirazo, S.; Mainardi, V.; Gerona, S.; Arbiza, J. Detection and molecular characterization of Torque Teno Virus (TTV) in Uruguay. Infect. Genet. Evol. 2016, 44, 501–506. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT; Nucleic Acids Symposium Series; Information Retrieval Ltd.: London, UK, 1999; Volume 41, pp. 95–98. [Google Scholar]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Hallett, R.L.; Clewley, J.P.; Bobet, F.; McKiernan, P.J.; Teo, C.G. Characterization of a highly divergent TT virus genome. J. Gen. Virol. 2000, 81 Pt 9, 2273–2279. [Google Scholar] [CrossRef]
- Muljono, D.H.; Nishizawa, T.; Tsuda, F.; Takahashi, M.; Okamoto, H. Molecular epidemiology of TT virus (TTV) and characterization of two novel TTV genotypes in Indonesia. Arch. Virol. 2001, 146, 1249–1266. [Google Scholar] [CrossRef]
- ICTV Report. Available online: https://talk.ictvonline.org/ictv-reports/ictv_9th_report/ssdna-viruses-2011/w/ssdna_viruses/139/anelloviridae (accessed on 18 December 2020).
- Christensen, J.K.; Eugen-Olsen, J.; SŁrensen, M.; Ullum, H.; Gjedde, S.B.; Pedersen, B.K.; Nielsen, J.O.; Krogsgaard, K. Prevalence and prognostic significance of infection with TT virus in patients infected with human immunodeficiency virus. J. Infect. Dis. 2000, 181, 1796–1799. [Google Scholar] [CrossRef]
- Madsen, C.D.; Eugen-Olsen, J.; Kirk, O.; Parner, J.; Kaae Christensen, J.; Brasholt, M.S.; Ole Nielsen, J.; Krogsgaard, K. TTV viral load as a marker for immune reconstitution after initiation of HAART in HIV-infected patients. HIV Clin. Trials 2002, 3, 287–295. [Google Scholar]
- Nishizawa, Y.; Tanaka, E.; Orii, K.; Rokuhara, A.; Ichijo, T.; Yoshizawa, K.; Kiyosawa, K. Clinical impact of genotype 1 TT virus infection in patients with chronic hepatitis C and response of TT virus to alpha-interferon. J. Gastroenterol. Hepatol. 2000, 15, 1292–1297. [Google Scholar] [PubMed]
- García-Álvarez, M.; Berenguer, J.; Alvarez, E.; Guzmán-Fulgencio, M.; Cosín, J.; Miralles, P.; Catalán, P.; López, J.C.; Rodríguez, J.M.; Micheloud, D.; et al. Association of torque teno virus (TTV) and torque teno mini virus (TTMV) with liver disease among patients coinfected with human immunodeficiency virus and hepatitis C virus. Eur. J. Clin. Microbiol. Infect. Dis. 2013, 32, 289–297. [Google Scholar] [CrossRef]
- AbuOdeh, R.; Al-Mawlawi, N.; Al-Qahtani, A.A.; Bohol, M.F.; Al-Ahdal, M.N.; Hasan, H.A.; AbuOdeh, L.; Nasrallah, G.K. Detection and genotyping of torque teno virus (TTV) in healthy blood donors and patients infected with HBV or HCV in Qatar. J. Med. Virol. 2015, 87, 1184–1191. [Google Scholar] [CrossRef]
- Maggi, F.; Andreoli, E.; Lanini, L.; Fornai, C.; Vatteroni, M.; Pistello, M.; Presciuttini, S.; Bendinelli, M. Relationships between total plasma load of torquetenovirus (TTV) and TTV genogroups carried. J. Clin. Microbiol. 2005, 43, 4807–4810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Deng, X.; Da Costa, A.C.; Bruhn, R.; Deeks, S.G.; Delwart, E. Virome analysis of antiretroviral-treated HIV patients shows no correlation between T-cell activation and anelloviruses levels. J. Clin. Virol. 2015, 72, 106–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocchi, J.; Ricci, V.; Albani, M.; Lanini, L.; Andreoli, E.; Macera, L.; Pistello, M.; Ceccherini-Nelli, L.; Bendinelli, M.; Maggi, F. Torquetenovirus DNA drives proinflammatory cytokines production and secretion by immune cells via toll-like receptor 9. Virology 2009, 394, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Simonetta, F.; Pradier, A.; Masouridi-Levrat, S.; van Delden, C.; Giostra, E.; Morard, I.; Mueller, N.; Muellhaupt, B.; Valli, P.V.; Semmo, N.; et al. Torque Teno Virus Load and Acute Rejection After Orthotopic Liver Transplantation. Transplantation 2017, 101, e219–e221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Görzer, I.; Jaksch, P.; Kundi, M.; Seitz, T.; Klepetko, W.; Puchhammer-Stöckl, E. Pre-transplant plasma Torque Teno virus load and increase dynamics after lung transplantation. PLoS ONE 2015, 10, e0122975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strassl, R.; Doberer, K.; Rasoul-Rockenschaub, S.; Herkner, H.; Görzer, I.; Kläger, J.P.; Schmidt, R.; Haslacher, H.; Schiemann, M.; Eskandary, F.A.; et al. Torque Teno Virus for Risk Stratification of Acute Biopsy-Proven Alloreactivity in Kidney Transplant Recipients. J. Infect. Dis. 2019, 219, 1934–1939. [Google Scholar] [CrossRef]


{kind=link}
| All Patients n (%) | HCV Mono-Infected n (%) | HIV/HCV Coinfected n (%) | |
|---|---|---|---|
| Overall | 92 (100.0%) | 29 (31.5%) | 63 (68.5%) |
| Gender | |||
| Male | 81 (88.0%) | 21 (72.4%) | 60 (95.2%) |
| Female | 11 (19%) | 8 (27.6%) | 3 (4.8%) |
| TTV DNA-PCR positive | 73 (79.3%) | 27 (93.1%) | 46 (73%) |
| HCV treatment response | |||
| SVR | 46 (50.0%) | 7 (24.1%) | 39 (61.9%) |
| Untreated | 24 (26.1%) | 11 (37.9%) | 13 (20.6%) |
| DAA failure | 22 (23.9%) | 11 (37.9%) | 11 (17.5%) |
| Cirrhosis | 28 (30.4%) | 10 (34.5%) | 18 (28.6%) |
| HCV genotype | |||
| 1a | 42 (45.7%) | 9 (31.0%) | 33 (52.4%) |
| 1b | 15 (16.3%) | 7 (24.1%) | 8 (12.7%) |
| 2 | 9 (5.4%) | 3 (10.3%) | 2 (3.2%) |
| 3 | 13 (14.1%) | 6 (20.7%) | 7 (11.1%) |
| 4 | 3 (3.3%) | 3 (10.3%) | 11 (17.5%) |
| Other/unknown | 3 (3.3%) | 1 (3.4%) | 2 (3.2%) |
| Age (median, IQ-range) | 53 (49–56) | 53 (48–59) | 53 (49–56) |
| TTV DNA, copies/mL (median, IQ range) | 2259 (50–31,446) | 29,715 (6578–55,841) | 778 (13–5777) |
| CD4 T cell count * (median, IQ range) | 611 (359–950) |
| TTV DNA PCR Pos (%) | p-Value * | TTV DNA PCR cp/mL, Median (IQ Range) | p-Value * | |
|---|---|---|---|---|
| Gender | ||||
| Male | 63/81 (77.8%) | 1351 (39–31,071) | ||
| Female | 10/11 (90.9%) | 0.449 | 31,446 (10,067–396,782) | 0.894 |
| HIV | ||||
| HIV/HCV coinfected | 46/63 (63.0%) | 881 (15–5818) | ||
| HCV monoinfected | 27/29 (93.1%) | 0.029 | 31,446 (6857–114,056) | 0.232 ** |
| HCV status | ||||
| SVR | 30 /46 (65.2%) | 469 (10–4124) | ||
| Untreated | 24/24 (100.0%) | <0.001 *** | 19,884 (5977–333,534) | 0.04 **** (**) |
| DAA failure | 19/22 (86.4%) | 0.06 *** | 4915 (450–64,535) | |
| Cirrhosis | ||||
| Yes | 23/28 (82.1%) | 6934 (558–80,913) | ||
| No | 50/64 (78.1%) | 0.784 | 1419 (42–28,914) | 0.523 |
| HCV genotype | ||||
| 1a | 34/42 (45.7%) | 4227 (155–30,609) | ||
| 1b | 13/15 (86.7%) | 8079 (575–55,841) | ||
| 2 | 3/5 (60.0%) | 50 (10–264,129) | ||
| 3 | 11/13 (84.6%) | 2259 (32–249,452) | ||
| 4 | 9/14 (64.3%) | 248 (10–17,882) | ||
| Other/unknown | 3/3 (100.0%) | 0.727 | 10,067 (1079–10,067) | 0.366 (1a vs. oth) ** |
| Age | ||||
| ≥55 | 29/33 (87.9%) | 6935 (566–28,114) | ||
| <55 | 44/59 (60.3%) | 0.181 | 1514 (18–36,679) | 0.216 ** |
| CD4 T cell count | ||||
| <500 | 16/19 (84.2%) | 469 (10–5337) | ||
| ≥500 | 26/40 (65.0%) | 0.218 | 1232 (159–11,399) | 0.311 |
| TTV DNA PCR Positive n. /Total (%) | TTV DNA PCR Copies/mLMedian (IQ Range) | |
|---|---|---|
| Group A | 12/22 (54.5%) | 95 (10–2204) |
| Group B | 16/22 (72.7%) | 830 (126–2590) |
| Group C | 5/6 (83.3%) | 348 (33–4677) |
| Group D | 5/7 (71.4%) | 10,067 (10–29,715) |
| Group E | 11/11 (100.0%) | 52,781 (8050–137,494) |
| Group F1 | 13/13 (100.0%) | 12,999 (2581–305,659) |
| Group F2 | 11/11 (100.0%) | 31,446 (7136–396,782) |
| TTV DNA PCR Pos Median (IQ Range) | TTV DNA PCR Neg Median (IQ Range) | p-Value * | |
|---|---|---|---|
| IL-6 | 2.87 (1.70–4.56) | 2.07 (1.82–5.77) | 0.810 |
| IL-4 | 0.00 (0.00–0.29) | 0.56 (0.00–1.96) | 0.200 |
| CD14 | 8577 (6822–10,273) | 9589 (7573–10,842) | 0.615 |
| IL-6 Median (IQ Range) | IL-4 * Median (IQ Range) | CD14 Median (IQ Range) | |
|---|---|---|---|
| Group A | 2.01 (1.37–5.46) | 1.24 (0.00–2.33) ** | 9009 (734–11,450) |
| Group B | 3.18 (2.58–4.64) | 0.00 (0.00–0.13) *** | 9326 (7814–10,928) |
| Group C | 1.91 (0.00–3.38) | ND | ND |
| Group D | 4.60 (1.47–10.13) | 0.28 (0.03–1.13) | 8625 (7405–10,782) |
| Group E | 2.96 (1.31–6.28) | 0.00 (0.00–0.40) | 8724 (6344–10,778) |
| Group F1 | 2.71 (1.62–3.67) | 0.00 (0.00–0.87) | 8717 (8132–13,177) |
| Group F2 | 3.68 (2.25–7.60) | 0.18 (0.02–0.59) | 8577 (8300–12,459) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lapa, D.; Del Porto, P.; Minosse, C.; D’Offizi, G.; Antinori, A.; Capobianchi, M.R.; Visco-Comandini, U.; McPhee, F.; Garbuglia, A.R.; Zaccarelli, M. Clinical Relevance of Torque Teno Virus (TTV) in HIV/HCV Coinfected and HCV Monoinfected Patients Treated with Direct-Acting Antiviral Therapy. J. Clin. Med. 2021, 10, 2092. https://doi.org/10.3390/jcm10102092
Lapa D, Del Porto P, Minosse C, D’Offizi G, Antinori A, Capobianchi MR, Visco-Comandini U, McPhee F, Garbuglia AR, Zaccarelli M. Clinical Relevance of Torque Teno Virus (TTV) in HIV/HCV Coinfected and HCV Monoinfected Patients Treated with Direct-Acting Antiviral Therapy. Journal of Clinical Medicine. 2021; 10(10):2092. https://doi.org/10.3390/jcm10102092
Chicago/Turabian StyleLapa, Daniele, Paola Del Porto, Claudia Minosse, Gianpiero D’Offizi, Andrea Antinori, Maria Rosaria Capobianchi, Ubaldo Visco-Comandini, Fiona McPhee, Anna Rosa Garbuglia, and Mauro Zaccarelli. 2021. "Clinical Relevance of Torque Teno Virus (TTV) in HIV/HCV Coinfected and HCV Monoinfected Patients Treated with Direct-Acting Antiviral Therapy" Journal of Clinical Medicine 10, no. 10: 2092. https://doi.org/10.3390/jcm10102092