Mechanisms of Membrane Curvature Generation in Membrane Traffic

Abstract
:1. Introduction
2. Lipids
2.1. Relationship between Asymmetric Lipid Distribution and Membrane Deformation
2.2. Phospholipid Flippases (P4-ATPases)

{kind=link}
| Class | Human | S. cerevisiae | C. elegans | A. thaliana |
|---|---|---|---|---|
| 1a | ATP8A1, ATP8A2 | DRS2 | TAT-1 | ALA1 |
| 1b | ATP8B1, ATP8B2, ATP8B3, ATP8B4 | - | TAT-2 | ALA2-12 |
| 2 | ATP9A, ATP9B | NEO1 | TAT-5, TAT-6 | |
| 3 | - | DNF1, DNF2 | - | |
| 4 | - | DNF3 | - | |
| 5 | ATP10A, ATP10B, ATP10D | - | TAT-3, TAT-4 | |
| 6 | ATP11A, ATP11B, ATP11C | - | - |
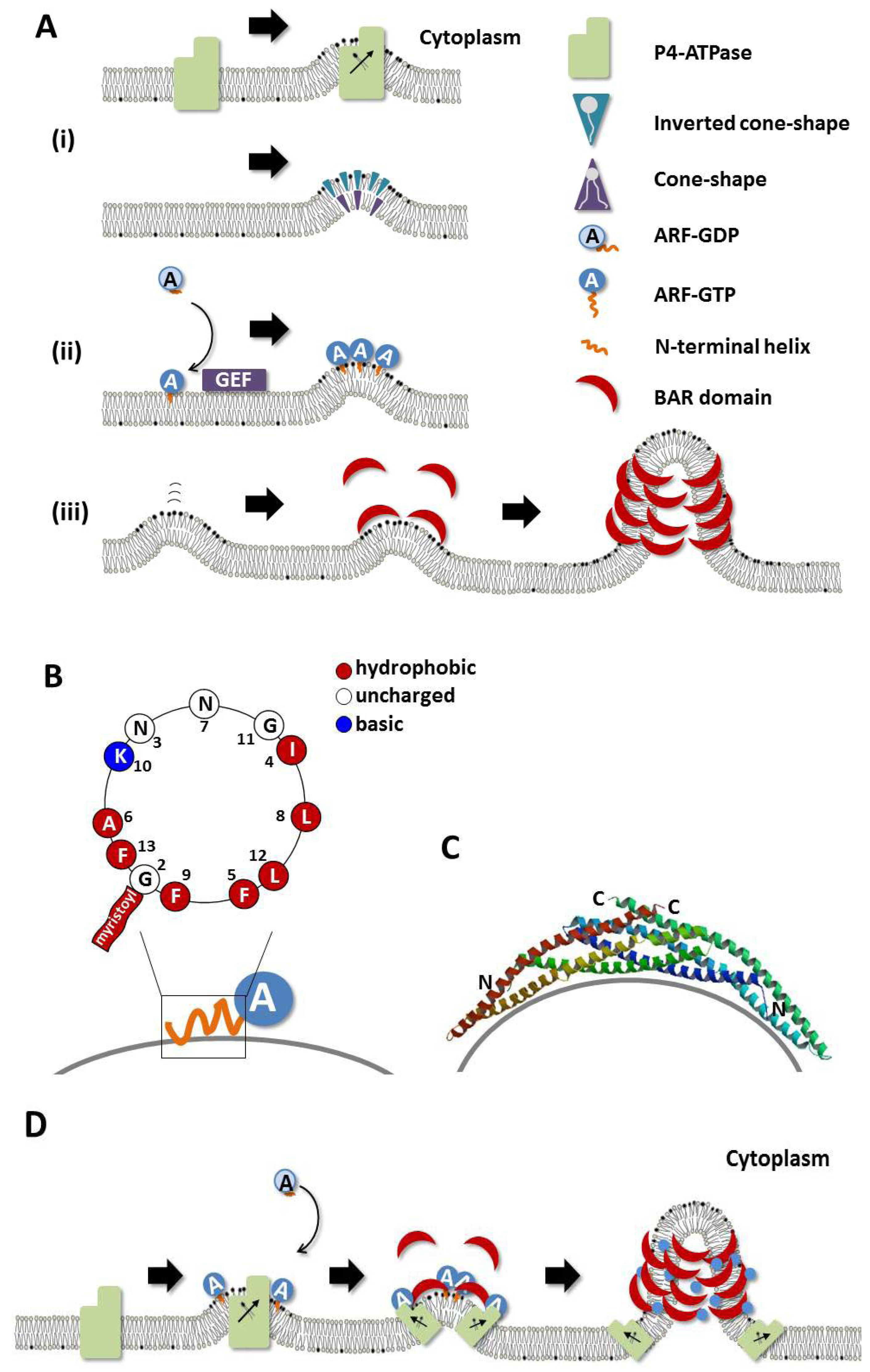
2.3. Lipid-Modifying Enzymes (LPAT and PLA2)
3. Proteins
3.1. Arf Family Small GTPases
3.2. BAR Domain-Containing Proteins
4. Interplay between Regulatory Proteins
4.1. Membrane Targeting of BAR Domain Proteins by Small GTPases
4.2. Arf Family Small GTPases and P4-ATPases
5. Conclusions
Acknowledgements
References
- Polishchuk, R.S.; Polishchuk, E.V.; Marra, P.; Alberti, S.; Buccione, R.; Luini, A.; Mironov, A.A. Correlative light-electron microscopy reveals the tubular-saccular ultrastructure of carriers operating between Golgi apparatus and plasma membrane. J. Cell Biol. 2000, 148, 45–58. [Google Scholar] [CrossRef]
- McMahon, H.T.; Gallop, J.L. Membrane curvature and mechanisms of dynamic cell membrane remodelling. Nature 2005, 438, 590–596. [Google Scholar] [CrossRef]
- Zimmerberg, J.; Kozlov, M.M. How proteins produce cellular membrane curvature. Nat. Rev. Mol. Cell Biol. 2006, 7, 9–19. [Google Scholar] [CrossRef]
- Schrier, S.L.; Zachowski, A.; Devaux, P.F. Mechanisms of amphipath-induced stomatocytosis in human erythrocytes. Blood 1992, 79, 782–786. [Google Scholar]
- Zwaal, R.F.; Schroit, A.J. Pathophysiologic implications of membrane phospholipid asymmetry in blood cells. Blood 1997, 89, 1121–1132. [Google Scholar]
- Van den Eijnde, S.M.; van den Hoff, M.J.; Reutelingsperger, C.P.; van Heerde, W.L.; Henfling, M.E.; Vermeij-Keers, C.; Schutte, B.; Borgers, M.; Ramaekers, F.C. Transient expression of phosphatidylserine at cell-cell contact areas is required for myotube formation. J. Cell Sci. 2001, 114, 3631–3642. [Google Scholar]
- Williamson, P.; Schlegel, R.A. Transbilayer phospholipid movement and the clearance of apoptotic cells. Biochim. Biophys. Acta 2002, 1585, 53–63. [Google Scholar] [CrossRef]
- Leventis, P.A.; Grinstein, S. The distribution and function of phosphatidylserine in cellular membranes. Annu. Rev. Biophys. 2010, 39, 407–427. [Google Scholar] [CrossRef]
- Emoto, K.; Kobayashi, T.; Yamaji, A.; Aizawa, H.; Yahara, I.; Inoue, K.; Umeda, M. Redistribution of phosphatidylethanolamine at the cleavage furrow of dividing cells during cytokinesis. Proc. Natl. Acad. Sci. USA 1996, 93, 12867–12872. [Google Scholar] [CrossRef]
- Sheetz, M.P.; Singer, S.J. Biological membranes as bilayer couples. A molecular mechanism of drug-erythrocyte interactions. Proc. Natl. Acad. Sci. USA 1974, 71, 4457–4461. [Google Scholar] [CrossRef]
- Devaux, P.F.; Herrmann, A.; Ohlwein, N.; Kozlov, M.M. How lipid flippases can modulate membrane structure. BBA-Biomembranes 2008, 1778, 1591–1600. [Google Scholar] [CrossRef]
- Papadopulos, A.; Vehring, S.; Lopez-Montero, I.; Kutschenko, L.; Stockl, M.; Devaux, P.F.; Kozlov, M.; Pomorski, T.; Herrmann, A. Flippase activity detected with unlabeled lipids by shape changes of giant unilamellar vesicles. J. Biol. Chem. 2007, 282, 15559–15568. [Google Scholar]
- Seigneuret, M.; Devaux, P.F. ATP-dependent asymmetric distribution of spin-labeled phospholipids in the erythrocyte membrane: Relation to shape changes. Proc. Natl. Acad. Sci. USA 1984, 81, 3751–3755. [Google Scholar] [CrossRef]
- Tang, X.; Halleck, M.S.; Schlegel, R.A.; Williamson, P. A subfamily of P-type ATPases with aminophospholipid transporting activity. Science 1996, 272, 1495–1497. [Google Scholar]
- Chen, C.Y.; Ingram, M.F.; Rosal, P.H.; Graham, T.R. Role for Drs2p, a P-type ATPase and potential aminophospholipid translocase, in yeast late Golgi function. J. Cell Biol. 1999, 147, 1223–1236. [Google Scholar] [CrossRef]
- Gomes, E.; Jakobsen, M.K.; Axelsen, K.B.; Geisler, M.; Palmgren, M.G. Chilling tolerance in arabidopsis involves ALA1, a member of a new family of putative aminophospholipid translocases. Plant Cell 2000, 12, 2441–2454. [Google Scholar] [Green Version]
- Daleke, D.L. Regulation of transbilayer plasma membrane phospholipid asymmetry. J. Lipid Res. 2003, 44, 233–242. [Google Scholar] [CrossRef]
- Pomorski, T.; Lombardi, R.; Riezman, H.; Devaux, P.F.; van Meer, G.; Holthuis, J.C.M. Drs2p-related P-type ATPases Dnf1p and Dnf2p are required for phospholipid translocation across the yeast plasma membrane and serve a role in endocytosis. Mol. Biol. Cell 2003, 14, 1240–1254. [Google Scholar] [CrossRef]
- Saito, K.; Fujimura-Kamada, K.; Furuta, N.; Kato, U.; Umeda, M.; Tanaka, K. Cdc50p, a protein required for polarized growth, associates with the Drs2p P-Type ATPase implicated in phospholipid translocation in saccharomyces cerevisiae. Mol. Biol. Cell 2004, 15, 3418–3432. [Google Scholar] [CrossRef]
- Zhou, X.; Graham, T.R. Reconstitution of phospholipid translocase activity with purified Drs2p, a type-IV P-type ATPase from budding yeast. Proc. Natl. Acad. Sci. USA 2009, 106, 16586–16591. [Google Scholar] [CrossRef]
- Zachowski, A.; Henry, J.P.; Devaux, P.F. Control of transmembrane lipid asymmetry in chromaffin granules by an ATP-dependent protein. Nature 1989, 340, 75–76. [Google Scholar] [CrossRef]
- Anne-Francoise, R.; Lars, N.; Fabrice, R.; Morten Krog, L.; Jean-Louis, B.; Simon, T. The C. elegans P4-ATPase TAT-1 regulates lysosome biogenesis and endocytosis. Traffic 2009, 10, 88–100. [Google Scholar] [CrossRef]
- Chen, B.; Jiang, Y.; Zeng, S.; Yan, J.; Li, X.; Zhang, Y.; Zou, W.; Wang, X. Endocytic sorting and recycling require membrane phosphatidylserine asymmetry maintained by TAT-1/CHAT-1. PLoS Genet. 2010, 6, e1001235. [Google Scholar] [CrossRef]
- Wehman, A.M.; Poggioli, C.; Schweinsberg, P.; Grant, B.D.; Nance, J. The P4-ATPase TAT-5 inhibits the budding of extracellular vesicles in C. elegans embryos. Curr. Biol. 2011, 21, 1951–1959. [Google Scholar] [CrossRef]
- Graham, T.R. Flippases and vesicle-mediated protein transport. Trends Cell Biol. 2004, 14, 670–677. [Google Scholar] [CrossRef]
- Natarajan, P.; Wang, J.; Hua, Z.; Graham, T.R. Drs2p-coupled aminophospholipid translocase activity in yeast Golgi membranes and relationship to in vivo function. Proc. Natl. Acad. Sci. USA 2004, 101, 10614–10619. [Google Scholar] [CrossRef]
- Gall, W.E.; Geething, N.C.; Hua, Z.; Ingram, M.F.; Liu, K.; Chen, S.I.; Graham, T.R. Drs2p-dependent formation of exocytic clathrin-coated vesicles in vivo. Curr. Biol. 2002, 12, 1623–1627. [Google Scholar] [CrossRef]
- Alder-Baerens, N.; Lisman, Q.; Luong, L.; Pomorski, T.; Holthuis, J.C.M. Loss of P4 ATPases Drs2p and Dnf3p disrupts aminophospholipid transport and asymmetry in yeast post-Golgi secretory vesicles. Mol. Biol. Cell 2006, 17, 1632–1642. [Google Scholar] [CrossRef]
- Poulsen, L.R.; Lopez-Marques, R.L.; McDowell, S.C.; Okkeri, J.; Licht, D.; Schulz, A.; Pomorski, T.; Harper, J.F.; Palmgren, M.G. The arabidopsis P4-ATPase ALA3 localizes to the Golgi and requires a {beta}-Subunit to function in lipid translocation and secretory vesicle formation. Plant Cell 2008, 20, 658–676. [Google Scholar] [CrossRef]
- Protein Data Bank, Research Collaboratory for Structural Bioinformatics (RCSB PDB) web site. Available online: www.pdb.org (accessed on 29 February 2012).
- Tarricone, C.; Xiao, B.; Justin, N.; Walker, P.A.; Rittinger, K.; Gamblin, S.J.; Smerdon, S.J. The structural basis of Arfaptin-mediated cross-talk between Rac and Arf signalling pathways. Nature 2001, 411, 215–219. [Google Scholar]
- Farge, E.; Ojcius, D.M.; Subtil, A.; Dautry-Varsat, A. Enhancement of endocytosis due to aminophospholipid transport across the plasma membrane of living cells. Am. J. Physiol. Cell Physiol. 1999, 276, C725–733. [Google Scholar]
- Paulusma, C.C.; Oude Elferink, R.P.J. The type 4 subfamily of P-type ATPases, putative aminophospholipid translocases with a role in human disease. BBA-Mol. Basis Dis. 2005, 1741, 11–24. [Google Scholar] [CrossRef]
- Paulusma, C.C.; Groen, A.; Kunne, C.; Ho-Mok, K.S.; Spijkerboer, A.L.; Rudi de Waart, D.; Hoek, F.J.; Vreeling, H.; Hoeben, K.A.; van Marle, J.; et al. Atp8b1 deficiency in mice reduces resistance of the canalicular membrane to hydrophobic bile salts and impairs bile salt transport. Hepatology 2006, 44, 195–204. [Google Scholar] [CrossRef]
- Wang, L.; Beserra, C.; Garbers, D.L. A novel aminophospholipid transporter exclusively expressed in spermatozoa is required for membrane lipid asymmetry and normal fertilization. Dev. Biol. 2004, 267, 203–215. [Google Scholar] [CrossRef]
- Dhar, M.S.; Sommardahl, C.S.; Kirkland, T.; Nelson, S.; Donnell, R.; Johnson, D.K.; Castellani, L.W. Mice heterozygous for Atp10c, a putative amphipath, represent a novel model of obesity and type 2 diabetes. J. Nutr. 2004, 134, 799–805. [Google Scholar]
- Folmer, D.E.; Elferink, R.P.J.O.; Paulusma, C.C. P4 ATPases-Lipid flippases and their role in disease. BBA-Mol. Cell Biol. L. 2009, 1791, 628–635. [Google Scholar]
- Flamant, S.; Pescher, P.; Lemercier, B.; Clément-Ziza, M.; Képès, F.; Fellous, M.; Milon, G.; Marchal, G.; Besmond, C. Characterization of a putative type IV aminophospholipid transporter P-type ATPase. Mamm. Genome 2003, 14, 21–30. [Google Scholar] [CrossRef]
- Siggs, O.M.; Arnold, C.N.; Huber, C.; Pirie, E.; Xia, Y.; Lin, P.; Nemazee, D.; Beutler, B. The P4-type ATPase ATP11C is essential for B lymphopoiesis in adult bone marrow. Nat. Immunol. 2011, 12, 434–440. [Google Scholar] [CrossRef]
- Yabas, M.; Teh, C.E.; Frankenreiter, S.; Lal, D.; Roots, C.M.; Whittle, B.; Andrews, D.T.; Zhang, Y.; Teoh, N.C.; Sprent, J.; et al. ATP11C is critical for the internalization of phosphatidylserine and differentiation of B lymphocytes. Nat. Immunol. 2011, 12, 441–449. [Google Scholar] [CrossRef]
- Hammond, K.; Reboiras, M.D.; Lyle, I.G.; Jones, M.N. Characterisation of phosphatidylcholine/ phosphatidylinositol sonicated vesicles. Effects of phospholipid composition on vesicle size. Biochim. Biophys. Acta 1984, 774, 19–25. [Google Scholar] [CrossRef]
- Sprong, H.; van der Sluijs, P.; van Meer, G. How proteins move lipids and lipids move proteins. Nat. Rev. Mol.Cell Biol. 2001, 2, 504–513. [Google Scholar]
- Scales, S.J.; Scheller, R.H. Cell biology: Lipid membranes shape up. Nature 1999, 401, 123–124. [Google Scholar]
- Brown, W.J.; Chambers, K.; Doody, A. Phospholipase A2 (PLA2) enzymes in membrane trafficking: Mediators of membrane shape and function. Traffic 2003, 4, 214–221. [Google Scholar] [CrossRef]
- Schmidt, J.A.; Brown, W.J. Lysophosphatidic acid acyltransferase 3 regulates Golgi complex structure and function. J. Cell Biol. 2009, 186, 211–218. [Google Scholar] [CrossRef]
- Drecktrah, D.; Chambers, K.; Racoosin, E.L.; Cluett, E.B.; Gucwa, A.; Jackson, B.; Brown, W.J. Inhibition of a Golgi complex lysophospholipid acyltransferase induces membrane tubule formation and retrograde trafficking. Mol. Biol. Cell 2003, 14, 3459–3469. [Google Scholar] [CrossRef]
- Kirchhausen, T. Three ways to make a vesicle. Nat. Rev. Mol. Cell Biol. 2000, 1, 187–198. [Google Scholar] [CrossRef]
- Kahn, R.A.; Cherfils, J.; Elias, M.; Lovering, R.C.; Munro, S.; Schurmann, A. Nomenclature for the human Arf family of GTP-binding proteins: ARF, ARL, and SAR proteins. J. Cell Biol. 2006, 172, 645–650. [Google Scholar] [CrossRef]
- Nakayama, K.; Wakatsuki, S. The structure and function of GGAs, the traffic controllers at the TGN sorting crossroads. Cell Struct. Func. 2003, 28, 431–442. [Google Scholar] [CrossRef]
- Robinson, M.S. Adaptable adaptors for coated vesicles. Trends Cell Biol 2004, 14, 167–174. [Google Scholar] [CrossRef]
- Bonifacino, J.S. The GGA proteins: Adaptors on the move. Nat. Rev. Mol. Cell Biol. 2004, 5, 23–32. [Google Scholar] [CrossRef]
- Whatmore, J.; Morgan, C.P.; Cunningham, E.; Collison, K.S.; Willison, K.R.; Cockcroft, S. ADP-ribosylation factor 1-regulated phospholipase D activity is localized at the plasma membrane and intracellular organelles in HL60 cells. Biochem. J. 1996, 320, 785–794. [Google Scholar]
- Honda, A.; Nogami, M.; Yokozeki, T.; Yamazaki, M.; Nakamura, H.; Watanabe, H.; Kawamoto, K.; Nakayama, K.; Morris, A.J.; Frohman, M.A.; Kanaho, Y. Phosphatidylinositol 4-phosphate 5-kinase alpha is a downstream effector of the small G protein ARF6 in membrane ruffle formation. Cell 1999, 99, 521–532. [Google Scholar] [CrossRef]
- Godi, A.; Pertile, P.; Meyers, R.; Marra, P.; di Tullio, G.; Iurisci, C.; Luini, A.; Corda, D.; de Matteis, M.A. ARF mediates recruitment of PtdIns-4-OH kinase-beta and stimulates synthesis of PtdIns(4,5)P2 on the Golgi complex. Nat. Cell Biol. 1999, 1, 280–287. [Google Scholar] [CrossRef]
- Shin, H.-W.; Nakayama, K. Dual control of membrane targeting by PtdIns(4)P and ARF. Trends Biochem. Sci. 2004, 29, 513–515. [Google Scholar] [CrossRef]
- Donaldson, J.G. Phospholipase D in endocytosis and endosomal recycling pathways. BBA-Mol. Cell Biol. L. 2009, 1791, 845–849. [Google Scholar]
- Goldberg, J. Structural and functional analysis of the ARF1-ARFGAP complex reveals a role for coatomer in GTP hydrolysis. Cell 1999, 96, 893–902. [Google Scholar] [CrossRef]
- Gillingham, A.K.; Munro, S. The small G proteins of the Arf family and their regulators. Ann. Rev. Cell Dev. Biol. 2007, 23, 579–611. [Google Scholar]
- Bielli, A.; Haney, C.J.; Gabreski, G.; Watkins, S.C.; Bannykh, S.I.; Aridor, M. Regulation of Sar1 NH2 terminus by GTP binding and hydrolysis promotes membrane deformation to control COPII vesicle fission. J. Cell Biol. 2005, 171, 919–924. [Google Scholar] [CrossRef]
- Lee, M.C.S.; Orci, L.; Hamamoto, S.; Futai, E.; Ravazzola, M.; Schekman, R. Sar1p N-terminal helix initiates membrane curvature and completes the fission of a COPII vesicle. Cell 2005, 122, 605–617. [Google Scholar] [CrossRef]
- Krauss, M.; Jia, J.Y.; Roux, A.; Beck, R.; Wieland, F.T.; de Camilli, P.; Haucke, V. Arf1-GTP-induced tubule formation suggests a function of Arf family proteins in curvature acquisition at sites of vesicle budding. J. Biol. Chem. 2008, 283, 27717–27723. [Google Scholar]
- Beck, R.; Sun, Z.; Adolf, F.; Rutz, C.; Bassler, J.; Wild, K.; Sinning, I.; Hurt, E.; Brugger, B.; Bethune, J.; Wieland, F. Membrane curvature induced by Arf1-GTP is essential for vesicle formation. Proc. Natl. Acad. Sci. USA 2008, 105, 11731–11736. [Google Scholar]
- Lundmark, R.; Doherty, G.J.; Vallis, Y.; Peter, B.J.; McMahon, H.T. Arf family GTP loading is activated by, and generates, positive membrane curvature. Biochem. J. 2008, 414, 189–194. [Google Scholar] [CrossRef]
- Long, K.R.; Yamamoto, Y.; Baker, A.L.; Watkins, S.C.; Coyne, C.B.; Conway, J.F.; Aridor, M. Sar1 assembly regulates membrane constriction and ER export. J. Cell Biol. 2010, 190, 115–128. [Google Scholar] [CrossRef]
- Farsad, K.; Ringstad, N.; Takei, K.; Floyd, S.R.; Rose, K.; de Camilli, P. Generation of high curvature membranes mediated by direct endophilin bilayer interactions. J. Cell Biol. 2001, 155, 193–200. [Google Scholar] [CrossRef]
- Gallop, J.L.; Jao, C.C.; Kent, H.M.; Butler, P.J.; Evans, P.R.; Langen, R.; McMahon, H.T. Mechanism of endophilin N-BAR domain-mediated membrane curvature. EMBO J. 2006, 25, 2898–2910. [Google Scholar] [CrossRef]
- Ford, M.G.; Mills, I.G.; Peter, B.J.; Vallis, Y.; Praefcke, G.J.; Evans, P.R.; McMahon, H.T. Curvature of clathrin-coated pits driven by epsin. Nature 2002, 419, 361–366. [Google Scholar]
- Takei, K.; Slepnev, V.I.; Haucke, V.; de Camilli, P. Functional partnership between amphiphysin and dynamin in clathrin-mediated endocytosis. Nat. Cell Biol. 1999, 1, 33–39. [Google Scholar] [CrossRef]
- Peter, B.J.; Kent, H.M.; Mills, I.G.; Vallis, Y.; Butler, P.J.G.; Evans, P.R.; McMahon, H.T. BAR domains as sensors of membrane curvature: The amphiphysin BAR structure. Science 2004, 303, 495–499. [Google Scholar]
- Habermann, B. The BAR-domain family of proteins: a case of bending and binding? EMBO Rep. 2004, 5, 250–255. [Google Scholar] [CrossRef]
- Itoh, T.; de Camilli, P. BAR, F-BAR (EFC) and ENTH/ANTH domains in the regulation of membrane-cytosol interfaces and membrane curvature. BBA-Mol. Cell Biol. L. 2006, 1761, 897–912. [Google Scholar]
- Masuda, M.; Mochizuki, N. Structural characteristics of BAR domain superfamily to sculpt the membrane. Semin. Cell Dev. Biol. 2010, 21, 391–398. [Google Scholar]
- Suetsugu, S.; Toyooka, K.; Senju, Y. Subcellular membrane curvature mediated by the BAR domain superfamily proteins. Semi. Cell Dev. Biol. 2010, 21, 340–349. [Google Scholar]
- Qualmann, B.; Koch, D.; Kessels, M.M. Let’s go bananas: Revisiting the endocytic BAR code. EMBO J. 2011, 30, 3501–3515. [Google Scholar] [CrossRef]
- Frost, A.; Unger, V.M.; de Camilli, P. The BAR domain superfamily: Membrane-molding macromolecules. Cell 2009, 137, 191–196. [Google Scholar] [CrossRef]
- Farsad, K.; De Camilli, P. Mechanisms of membrane deformation. Curr. Opin. Cell Biol. 2003, 15, 372–381. [Google Scholar] [CrossRef]
- Shimada, A.; Niwa, H.; Tsujita, K.; Suetsugu, S.; Nitta, K.; Hanawa-Suetsugu, K.; Akasaka, R.; Nishino, Y.; Toyama, M.; Chen, L.; et al. Curved EFC/F-BAR-domain dimers are joined end to end into a filament for membrane invagination in endocytosis. Cell 2007, 129, 761–772. [Google Scholar] [CrossRef]
- Frost, A.; Perera, R.; Roux, A.; Spasov, K.; Destaing, O.; Egelman, E.H.; de Camilli, P.; Unger, V.M. Structural basis of membrane invagination by F-BAR domains. Cell 2008, 132, 807–817. [Google Scholar] [CrossRef]
- Wang, Q.; Navarro, M.V.A.S.; Peng, G.; Molinelli, E.; Goh, S.L.; Judson, B.L.; Rajashankar, K.R.; Sondermann, H. Molecular mechanism of membrane constriction and tubulation mediated by the F-BAR protein Pacsin/Syndapin. Proc. Natl. Acad. Sci. USA 2009, 106, 12700–12705. [Google Scholar]
- Henne, W.M.; Boucrot, E.; Meinecke, M.; Evergren, E.; Vallis, Y.; Mittal, R.; McMahon, H.T. FCHo proteins are nucleators of clathrin-mediated endocytosis. Science 2010, 328, 1281–1284. [Google Scholar] [CrossRef]
- Wu, M.; Huang, B.; Graham, M.; Raimondi, A.; Heuser, J.E.; Zhuang, X.; de Camilli, P. Coupling between clathrin-dependent endocytic budding and F-BAR-dependent tubulation in a cell-free system. Nat. Cell Biol. 2010, 12, 902–908. [Google Scholar] [CrossRef]
- Dawson, J.C.; Legg, J.A.; Machesky, L.M. Bar domain proteins: A role in tubulation, scission and actin assembly in clathrin-mediated endocytosis. Trends Cell Biol. 2006, 16, 493–498. [Google Scholar] [CrossRef]
- Ferguson, S.; Raimondi, A.; Paradise, S.; Shen, H.; Mesaki, K.; Ferguson, A.; Destaing, O.; Ko, G.; Takasaki, J.; Cremona, O.; O’Toole, E.; de Camilli, P. Coordinated actions of actin and BAR proteins upstream of dynamin at endocytic clathrin-coated pits. Dev. Cell 2009, 17, 811–822. [Google Scholar] [CrossRef]
- Nie, Z.; Hirsch, D.S.; Luo, R.; Jian, X.; Stauffer, S.; Cremesti, A.; Andrade, J.; Lebowitz, J.; Marino, M.; Ahvazi, B.; et al. A BAR domain in the N terminus of the Arf GAP ASAP1 affects membrane structure and trafficking of epidermal growth factor receptor. Curr. Biol. 2006, 16, 130–139. [Google Scholar] [CrossRef]
- Zhu, G.; Chen, J.; Liu, J.; Brunzelle, J.S.; Huang, B.; Wakeham, N.; Terzyan, S.; Li, X.; Rao, Z.; Li, G.; Zhang, X.C. Structure of the APPL1 BAR-pH domain and characterization of its interaction with Rab5. EMBO J. 2007, 26, 3484–3493. [Google Scholar]
- Soulet, F.; Yarar, D.; Leonard, M.; Schmid, S.L. SNX9 regulates dynamin assembly and is required for efficient clathrin-mediated endocytosis. Mol. Biol. Cell 2005, 16, 2058–2067. [Google Scholar] [CrossRef]
- Yarar, D.; Waterman-Storer, C.M.; Schmid, S.L. SNX9 couples actin assembly to phosphoinositide signals and is required for membrane remodeling during endocytosis. Dev. Cell 2007, 13, 43–56. [Google Scholar] [CrossRef]
- Pylypenko, O.; Lundmark, R.; Rasmuson, E.; Carlsson, S.R.; Rak, A. The PX-BAR membrane-remodeling unit of sorting nexin 9. EMBO J. 2007, 26, 4788–4800. [Google Scholar] [CrossRef]
- Cozier, G.E.; Carlton, J.; McGregor, A.H.; Gleeson, P.A.; Teasdale, R.D.; Mellor, H.; Cullen, P.J. The phox homology (PX) domain-dependent, 3-phosphoinositide-mediated association of sorting nexin-1 with an early sorting endosomal compartment is required for its ability to regulate epidermal growth factor receptor degradation. J. Biol. Chem. 2002, 277, 48730–48736. [Google Scholar]
- Carlton, J.; Bujny, M.; Peter, B.J.; Oorschot, V.M.J.; Rutherford, A.; Mellor, H.; Klumperman, J.; McMahon, H.T.; Cullen, P.J. Sorting nexin-1 mediates tubular endosome-to-TGN transport through coincidence sensing of high-curvature membranes and 3-phosphoinositides. Curr. Biol. 2004, 14, 1791–1800. [Google Scholar] [CrossRef]
- Carlton, J.G.; Bujny, M.V.; Peter, B.J.; Oorschot, V.M.; Rutherford, A.; Arkell, R.S.; Klumperman, J.; McMahon, H.T.; Cullen, P.J. Sorting nexin-2 is associated with tubular elements of the early endosome, but is not essential for retromer-mediated endosome-to-TGN transport. J. Cell Sci. 2005, 118, 4527–4539. [Google Scholar] [CrossRef]
- Cullen, P.J. Endosomal sorting and signalling: an emerging role for sorting nexins. Nat. Rev. Mol. Cell Biol. 2008, 9, 574–582. [Google Scholar] [CrossRef]
- Van Weering, J.R.; Verkade, P.; Cullen, P.J. SNX-BAR proteins in phosphoinositide-mediated, tubular-based endosomal sorting. Semin. Cell Dev. Biol. 2010, 21, 371–380. [Google Scholar]
- Van Weering, J.R.; Verkade, P.; Cullen, P.J. SNX-BAR-mediated endosome tubulation is co-ordinated with endosome maturation. Traffic 2012, 13, 94–107. [Google Scholar] [CrossRef]
- Man, Z.; Kondo, Y.; Koga, H.; Umino, H.; Nakayama, K.; Shin, H.W. Arfaptins are localized to the trans-Golgi by interaction with Arl1, but Not Arfs. J. Biol. Chem. 2011, 286, 11569–11578. [Google Scholar]
- Lu, L.; Horstmann, H.; Ng, C.; Hong, W. Regulation of Golgi structure and function by ARF-like protein 1 (Arl1). J. Cell Sci. 2001, 114, 4543–4555. [Google Scholar]
- Lu, L.; Tai, G.; Hong, W. Autoantigen Golgin-97, an effector of Arl1 GTPase, participates in traffic from the endosome to the trans-Golgi network. Mol. Biol. Cell 2004, 15, 4426–4443. [Google Scholar] [CrossRef]
- Shin, H.-W.; Kobayashi, H.; Kitamura, M.; Waguri, S.; Suganuma, T.; Uchiyama, Y.; Nakayama, K. Roles of ARFRP1 (ADP-ribosylation factor-related protein 1) in post-Golgi membrane trafficking. J. Cell Sci. 2005, 118, 4039–4048. [Google Scholar] [CrossRef]
- Nishimoto-Morita, K.; Shin, H.W.; Mitsuhashi, H.; Kitamura, M.; Zhang, Q.; Johannes, L.; Nakayama, K. Differential effects of depletion of ARL1 and ARFRP1 on membrane trafficking between the trans-Golgi network and endosomes. J. Biol. Chem. 2009, 284, 10583–10592. [Google Scholar]
- Miaczynska, M.; Christoforidis, S.; Giner, A.; Shevchenko, A.; Uttenweiler-Joseph, S.; Habermann, B.; Wilm, M.; Parton, R.G.; Zerial, M. APPL proteins link Rab5 to nuclear signal transduction via an endosomal compartment. Cell 2004, 116, 445–456. [Google Scholar] [CrossRef]
- Hua, Z.; Fatheddin, P.; Graham, T.R. An essential subfamily of Drs2p-related P-type ATPases is required for protein trafficking between Golgi complex and endosomal/vacuolar system. Mol. Biol. Cell 2002, 13, 3162–3177. [Google Scholar] [CrossRef]
- Liu, K.; Surendhran, K.; Nothwehr, S.F.; Graham, T.R. P4-ATPase requirement for AP-1/Clathrin function in protein transport from the trans-golgi network and early endosomes. Mol. Biol. Cell 2008, 19, 3526–3535. [Google Scholar] [CrossRef]
- Chantalat, S.; Park, S.K.; Hua, Z.; Liu, K.; Gobin, R.; Peyroche, A.; Rambourg, A.; Graham, T.R.; Jackson, C.L. The Arf activator Gea2p and the P-type ATPase Drs2p interact at the Golgi in saccharomyces cerevisiae. J. Cell Sci. 2004, 117, 711–722. [Google Scholar] [CrossRef]
- Jochum, A.; Jackson, D.; Schwarz, H.; Pipkorn, R.; Singer-Kruger, B. Yeast Ysl2p, homologous to Sec7 domain guanine nucleotide exchange factors, functions in endocytosis and maintenance of vacuole integrity and interacts with the Arf-like small GTPase Arl1p. Mol. Cell. Biol. 2002, 22, 4914–4928. [Google Scholar] [CrossRef]
- Wicky, S.; Schwarz, H.; Singer-Kruger, B. Molecular interactions of yeast neo1p, an essential member of the drs2 family of aminophospholipid translocases, and its role in membrane trafficking within the endomembrane system. Mol. Cell. Biol. 2004, 24, 7402–7418. [Google Scholar] [CrossRef]
- Takatsu, H.; Baba, K.; Shima, T.; Umino, H.; Kato, U.; Umeda, M.; Nakayama, K.; Shin, H.W. ATP9B, a P4-ATPase (a putative aminophospholipid translocase), localizes to the trans-golgi network in a cdc50 protein-independent manner. J. Biol. Chem. 2011, 286, 38159–38167. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Shin, H.-W.; Takatsu, H.; Nakayama, K. Mechanisms of Membrane Curvature Generation in Membrane Traffic. Membranes 2012, 2, 118-133. https://doi.org/10.3390/membranes2010118
Shin H-W, Takatsu H, Nakayama K. Mechanisms of Membrane Curvature Generation in Membrane Traffic. Membranes. 2012; 2(1):118-133. https://doi.org/10.3390/membranes2010118
Chicago/Turabian StyleShin, Hye-Won, Hiroyuki Takatsu, and Kazuhisa Nakayama. 2012. "Mechanisms of Membrane Curvature Generation in Membrane Traffic" Membranes 2, no. 1: 118-133. https://doi.org/10.3390/membranes2010118
APA StyleShin, H.-W., Takatsu, H., & Nakayama, K. (2012). Mechanisms of Membrane Curvature Generation in Membrane Traffic. Membranes, 2(1), 118-133. https://doi.org/10.3390/membranes2010118