1. Introduction
Bacterial outer membrane vesicles (OMVs) are extracellular vesicles with defined membrane structures that are actively secreted by Gram-negative bacteria [
1]. Their particle size distribution is approximately 20–250 nm, and they cannot replicate [
2]. OMVs contain various parent bacterial components that endow them with unique biological functions, including lipopolysaccharides, peptidoglycans, phospholipids, proteins, and nucleic acids (e.g., DNA and RNA) [
3,
4,
5,
6]. OMVs are crucial mediators of intercellular communication in bacteria–bacteria and bacteria–host interactions [
7,
8,
9,
10]. OMVs regulate various bacterial interactions, including biofilm formation, antibiotic resistance, and the killing of competing bacteria [
11]. OMVs also stimulate the activities of epithelial cells, endothelial cells, and various immune cells in the host [
12,
13]. Proteomic analyses of OMVs provide information on their protein composition, biogenesis, and function. The gradual development of liquid chromatography with tandem mass spectrometry (LC–MS/MS) over recent years has played a pivotal role in promoting the development of proteomics research into OMVs [
14,
15,
16]. Lee et al. used LC–MS/MS to demonstrate that OMVs derived from
Escherichia coli bacteria contain various outer membrane proteins and certain porins, such as OmpA, OmpF, and NmpC. These proteins have immunostimulatory activities and can induce leukocyte migration [
17]. Kwon et al. reported that OMVs derived from
Acinetobacter baumannii contained 132 proteins, of which 43 are cytoplasmic, 26 are outer membrane proteins, 8 are inner membrane proteins, 6 are periplasmic, and the rest have an unknown localization [
18]. Choi et al. reported that OMVs derived from
Pseudomonas aeruginosa contained 338 proteins [
19]. Among these proteins, the outer membrane proteins are mainly involved in the assembly of bacterial flagella, pore formation, the transport of specific substrates, and outer-membrane stability. The other proteins were found to be related to neutrophil-mediated killing.
The intestine is the largest immune organ in the body. The intestinal mucosal innate immune system, which comprises intestinal mucosal tissue, immune cells, and immune molecules, represents the body’s primary line of defense against pathogenic invasion. Upon interaction with pathogenic microorganisms, the intestinal mucosal innate immune system can elicit an immune response, thereby contributing to the body’s defense mechanisms [
20,
21]. As important innate immune cells, macrophages are among the immune cell models commonly used to study innate immune responses in vitro, and they play crucial roles in intestinal mucosal immunity.
Escherichia coli Nissle 1917 (EcN) is a Gram-negative probiotic that resides in the human intestine. The research has indicated that EcN positively contributes to regulating host immune function and treating intestinal diseases [
22]. Compared to EcN probiotics, EcN-derived OMVs exhibit a high propensity to disseminate, traverse the mucus layer, and migrate directly to the intestinal submucosal tissue to interact with host immune-system cells. Importantly, EcN-derived OMVs preserve the beneficial and immunogenic characteristics of the parent bacterium while mitigating the potential risks associated with administering live bacteria to the host. Canas et al. reported that EcN-derived OMVs mediated intestinal anti-inflammatory responses and barrier-protective effects in a mouse model of DSS-induced intestinal inflammation [
23]. Fábrega et al. demonstrated that the oral administration of EcN-derived OMVs in a mouse model effectively reduced pro-inflammatory factor levels, increased the expression of intestinal barrier markers, and consequently inhibited the progression of DSS-induced colitis [
24]. Alvarez et al. reported that EcN-derived OMVs activated intestinal mucosal immune and defense responses. Furthermore, these EcN-derived OMVs enhance barrier function by increasing the expression of tight junction proteins in intestinal epithelial cells [
25]. There have been few studies on the heterogeneity of EcN-derived OMVs in terms of size. Isolating EcN-derived OMVs of different sizes represents the first and most important step toward conducting research regarding their heterogeneity. Currently, fast, effective, and efficient methods for the refined preparation of OMVs of different sizes are lacking. Ultrafiltration is currently used as a method for the isolation of OMVs [
2]. It separates them based on their distinct physical characteristics using a combination of filter membranes of different sizes to facilitate the separation process. Inspired by the principles of ultrafiltration, we developed a membrane filtration method that is convenient and effective for separating EcN-derived OMVs of different sizes. This method has the advantages of not requiring any additional reagents, being simple to execute, being low-cost, being able to preserve the biological activity of OMVs, and being scalable for industrial mass production.
EcN-derived OMVs are pivotal in modulating intestinal immune responses. Compared to the probiotic EcN itself, EcN-derived OMVs offer unique advantages [
26]. There are relatively few studies addressing the heterogeneity of EcN-derived OMVs of varying sizes, which significantly restricts research into their clinical applications. In this study, we first used the gradient filtration method to isolate high-purity OMVs derived from EcN and then conducted proteomic analyses on the isolated EcN-derived OMVs. Second, we used the filtration method to obtain EcN-derived OMVs of three specific sizes: <50 nm, 50–100 nm, and 100–300 nm. We then conducted proteomic analyses on the EcN-derived OMVs of three specific sizes. Finally, we analyzed the interactions of EcN-derived OMVs of three specific sizes with RAW264.7 macrophages to characterize their differential effects. Our results showed that the isolated EcN-derived OMVs contained a total of 646 different proteins, and the EcN-derived OMVs of different sizes contained different protein types. Among them, we found that the 50–100 nm group contained the widest variety of proteins compared to those in the <50 nm and 100–300 nm groups. We also found that RAW264.7 macrophages secreted different amounts of IL-1β and IL-10 following co-incubation with the three EcN-derived OMV types. The 50–100 nm EcN-derived OMV group showed a stronger effect in terms of inducing inflammatory cytokine secretion than the other two groups. These findings confirmed that EcN-derived OMVs of varying sizes exhibited heterogeneous characteristics.
2. Materials and Methods
2.1. RAW264.7 Macrophage Culture
RAW264.7 macrophages were purchased from the Shanghai Cell Biology Institute of the Chinese Academy of Sciences (Shanghai, China). Fetal bovine serum (FBS) and Dulbecco’s Modified Eagle Medium (DMEM) were purchased from Thermo Fisher Scientific (Waltham, MA, USA). Penicillin-streptomycin solution was purchased from Beyotime Biotechnology (Shanghai, China). RAW264.7 macrophages were cultured in DMEM with 10% FBS and 1% penicillin-streptomycin. The cells were cultured in a CO2 incubator with a humidified atmosphere of 5% CO2/95% air at 37 °C.
2.2. Isolation of EcN-Derived OMVs and Size-Based Sorting
2.2.1. Isolation of EcN-Derived OMVs Through Gradient Filtration
The EcN bacterial stock was purchased from Baosai Biological Company (Hangzhou, China). EcN-derived OMVs were obtained via the gradient filtration protocol previously reported by our research group [
27]. We optimized the protocol for the gradient filtration and significantly improved the purity of the OMV samples by repeating the pretreatment and washing steps multiple times. The detailed protocol for the isolation of EcN-derived OMVs is described below. Aliquots of EcN glycerol cryopreservation solution (100 µL) were inoculated into 10 mL of sterile LB culture medium (Sevier Biotechnology, Wuhan, China) at a 1:100 ratio and cultured at 37 °C at 200 rpm for 8 h. Subsequently, 200 µL of this seed mixture was inoculated into 200 mL of sterile LB culture medium and cultured at 37 °C at 200 rpm for 18 h. Finally, 200 mL of EcN culture medium was collected. The EcN culture medium was then centrifuged at 15,000×
g at 4 °C to remove the bacterial cells and any sediments. The supernatant was subsequently filtered through a 0.45 µm filter (Millipore, Darmstadt, Germany) to remove residual cellular debris. The resulting supernatant was then filtered through a 0.3 µm filter membrane (Millipore). Finally, the collected filtrate was filtered and centrifuged in an ultrafiltration centrifuge tube (100 kDa, UFC9100; Millipore) at 5000×
g until the remaining volume was approximately 0.5 mL. Residual proteins were removed via three consecutive PBS washes (12 mL of PBS solution per wash). The ultrafiltration centrifuge tube was then centrifuged at 5000×
g for 30 min. The final precipitate (i.e., purified EcN-derived OMVs) was dissolved in 200 µL of PBS and stored at –80 °C for later use.
2.2.2. Size-Based Sorting of EcN-Derived OMVs of Varying Sizes
The EcN-derived OMV concentrate was first separated via membrane filtration with different pore sizes (100 nm, UFC30VV25; Millipore) at 5000× g for 5 min, with both upper solutions and lower filtrates collected to obtain <100 nm and 100–300 nm concentrates. The <100 nm concentrate was then passed through a 50 nm filter (50 nm, VMWP02500; Millipore) to separate the OMVs into <50 nm and 50–100 nm concentrates. The different OMV classes were then concentrated via ultrafiltration (100 kDa; Millipore) at 5000× g for 15 min.
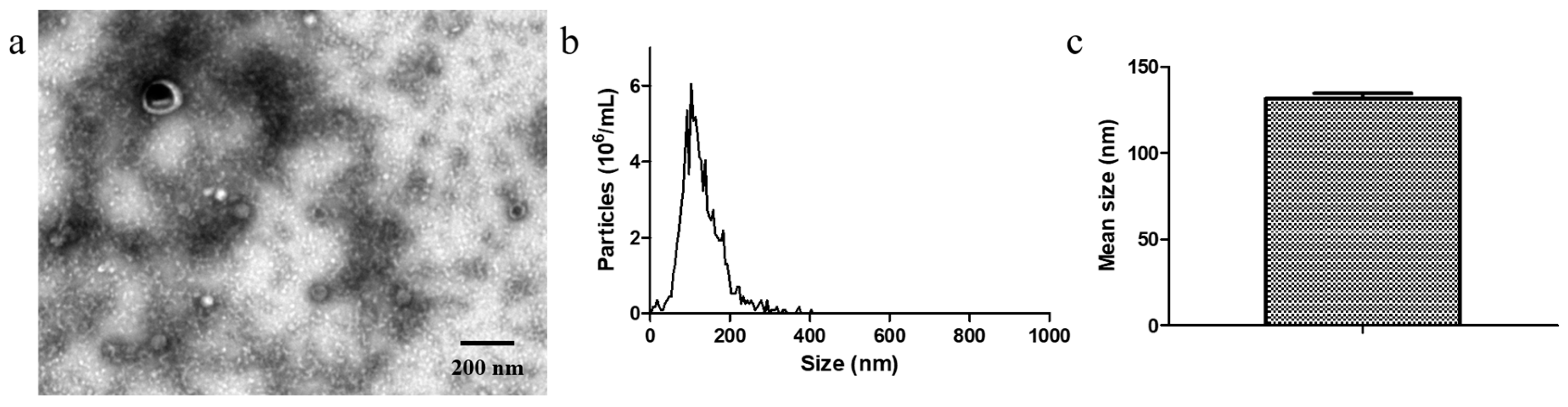
2.3. TEM of EcN-Derived OMVs
TEM images were acquired using a JEM1011 electron microscope (JEOL, Tokyo, Japan). The EcN-derived OMV samples were loaded onto TEM copper grids and dried for 20 min. The samples were negatively stained with 2% phosphotungstic acid for 10 min before TEM imaging, which was performed at 80 kV.
2.4. NTA of EcN-Derived OMVs
NTA was performed on a NanoSight NS300 instrument (Malvern, Worchestershire, UK). The EcN-derived OMV samples were warmed to room temperature and vortexed before being serially diluted to break apart any aggregates. A 10 µL volume of the resulting mixture was then diluted with PBS to achieve a concentration ranging from 1 × 107 to 1 × 109 particles/mL. The size distribution of the OMVs was then determined via a ZetaView PMX 110 particle matrix at 405 nm.
2.5. DLS of EcN-Derived OMVs of Varying Sizes
DLS was performed on a Nano ZS90 instrument (Malvern). The size distribution of the EcN-derived OMVs was characterized via DLS. A 10 µL volume of the resulting mixture was diluted with 1 mL of PBS and added to a DLS test cup to perform the determination.
2.6. BCA Protein Quantification of EcN-Derived OMVs
A bicinchoninic acid (BCA) protein assay kit (Beyotime Biotechnology, Shanghai, China) was used to prepare a standard curve per the manufacturer’s instructions. A fixed volume of each EcN-derived OMV sample was then mixed with radioimmunoprecipitation assay lysis solution. The mixture was oscillated at room temperature for 15 min. A 2 µL volume of each EcN-derived OMV sample lysate was obtained, and its absorbance (562 nm) was measured via a SpectraMax M5 full-wavelength microplate reader (Molecular Devices, Sunnyvale, CA, USA). The concentration of the EcN-derived OMVs was then determined according to the BCA standard curve and sample dilution factor.
2.7. Purification of EcN-Derived OMV Samples via Size-Exclusion Chromatography
Size-exclusion chromatography (SEC) was performed using the Exosupur® Purification Kit (Beijing Enzekantai Company, Beijing, China, ES914) according to the manufacturer’s instructions: first, the Exosupur® exclusion column was removed from the refrigerator, maintained at 4 °C, and stabilized in an upright orientation using a test tube rack. The column was then allowed to equilibrate to ambient temperature for 30 min. Subsequently, the column was rinsed with 20 mL of PBS buffer, which was twice the column’s volume. After the PBS was completely drained, 1 mL of the sample was added (if the sample was less than 1 mL, PBS was added until the total volume was 1 mL). After all samples were transferred into the sieve plate, 10 mL of the PBS eluent was added. Starting from the first drop of effluent, the effluent was collected using 1.5 mL centrifuge tubes. Each 500 µL of the effluent was marked as one fraction. Next, the OMVs were collected. It was recommended to discard fractions 1–4 and only retain fractions 5–7, as these fractions contained the highest concentration of OMVs. These collected fractions were combined into a single solution. The protein impurities predominantly appeared in the fractions collected after fraction 10, so fractions 10–15 were also collected and combined separately as a mixed protein sample. The collected OMVs and mixed protein solutions were concentrated via ultrafiltration centrifuge tube (100 kDa, UFC9100; Millipore). Subsequently, 200 µL of PBS was added to obtain an OMV concentrate and impure protein concentrate. These concentrates were subjected to quantitative analysis using the BCA protein quantification method.
2.8. SDS-PAGE of EcN-Derived OMVs
The SDS-PAGE gel preparation kit was purchased from Beyotime Biotechnology (Shanghai, China). The EcN-derived OMVs (10 µg) were boiled in 5× SDS-PAGE loading buffer for 10 min and then centrifuged (12,000× g), and the resulting supernatants were analyzed via SDS-PAGE on a 12% separating gel stained with G-250 protein-staining reagent (Baiolaibo Technology, Beijing, China).
2.9. Proteomic Study of EcN-Derived OMVs
Protein expression was verified using Wayen Biotechnology (Shanghai, China) via LC-MS/MS using label-free proteomic analysis, according to their established protocols [
28]. Proteomic experiments with EcN-derived OMV samples were performed in biological quadruplicate. Proteomic experiments with EcN-derived OMVs of three specific sizes were performed once each. The LC-MS-MS/MS protocol is described below.
2.9.1. Preprocessing of Proteomic Samples
Equal volumes of protein lysis solution [7 M urea (Sigma, St. Louis, MO, USA), 2% SDS (Sigma), and 1% protease inhibitor cocktail (Thermo Scientific, San Jose, CA, USA)] were added to the EcN-derived OMV samples, after which an ultrasonic cell disrupter (Xinzhi Biotechnology, Ningbo, China) was used for lysis. The samples were repeatedly sonicated for 2 s, with an intervening pause of 5 s, for a total of 1 min, before being lysed on ice for 2 h. The lysed product was then centrifuged at 13,000 rpm for 20 min at 4 °C. The supernatant was collected, and its concentration was measured via BCA quantification (Beyotime Biotechnology, Shanghai, China). The samples (50 µL) of this protein mixture were then added to 10 mM dithiothreitol (DTT; Sigma) and incubated at 37 °C for 1 h. The solutions were then centrifuged at 13,000 rpm for 1 h at 4 °C to collect precipitates. The collected precipitates were added to 1 mL aliquots of 100% acetone (Thermo Scientific) for purification. The samples were then centrifuged at 13,000 rpm for 30 min, the supernatant was discarded, and the precipitate was subjected to repeated washing. The precipitate was then dried at room temperature for 10 min, dissolved in PBS, and incubated with trypsin (Promega, Sunnyvale, CA, USA) at 37 °C overnight for enzymatic digestion. Following centrifugation for 30 min at 13,000 rpm, the peptide precipitate was obtained and desalted via a C18 desalting column (GL Sciences, Tokyo, Japan).
2.9.2. LC-MS/MS Analysis
The drained peptide sample was redissolved in the mobile phase and centrifuged at 20,000× g for 10 min, after which the supernatant was collected for injection-processing. The peptide sample was first separated via a nanoElute UPLC liquid-phase system (Brucker, Karlsruhe, Germany). Mobile phase A consisted of 0.1% formic acid (Aladdin, Los Angeles, CA, USA) in water, and the flow rate was 300 μL/min. Mobile phase B [0.1% formic acid (Aladdin) in acetonitrile (Thermo Scientific)] was run at 2% for the first 5 min, ramped to 22% over 45 min, and ramped to 35% over 50 min, before being rapidly increased to 80% for 55 min. The peptide fragments separated by the liquid phase were ionized via the CaptiveSpray source before being input into the timsTOF Pro 2 tandem mass spectrometer (Brucker) for mode detection. The main parameters used in the mass spectrometer were as follows: ion source voltage, 1.5 kV; ion mobility range, 0.76–1.29 V·S/cm2; first-level mass spectrometry scanning range, 452–1152 m/z; and peak intensity detected only at >2500. The 452–1152 m/z range was divided into four steps. Each step was divided into seven windows, and a total of 56 windows were used for continuous window fragmentation and information collection. The fragmentation mode was the charge injection device, and the fragmentation energy was 20–59 eV. The mass width of each window was 25. The cycle time of each DIA scan was 1.59 s.
2.9.3. Database Searching, Protein Identification, and Statistical Analyses
The obtained MS/MS data were analyzed via Spectronaut 17 (Biognosys, Schlieren, Switzerland). Database searches were performed via the UniProt database according to a previously reported protocol [
29]. For proteins derived from the UniProt database, the UniProt ID was first used to match the GO ID, and the corresponding protein information was retrieved from the UniProt-GOA database on the basis of the GO ID. The following search parameters were used: enzyme digestion of Trypsin, with a parent tolerance of 15 ppm; fragment tolerance of 0.5 Da; minimum peptide length of 5; false discovery rate of 1% at the peptide and protein levels; cysteine carbamidomethylation as fixed modification; and methionine oxidation and protein N-terminal acetylation as variable modifications. The identified proteins were classified according to their subcellular localization and molecular functions via CELLO (
http://cello.life.nctu.edu.tw/ (accessed on 1 September 2024)) and InterProScan (
http://www.ebi.ac.uk/interpro/ (accessed on 1 September 2024)), respectively [
30]. The COG protein functional classification method was used to classify the protein molecules involved in various biological processes.
2.10. Internalization of EcN-Derived OMVs of Diverse Sizes by RAW264.7 Macrophages
For the negative control group, 500 µL of PBS was combined with 0.5 µL of DiO membrane dye. In the experimental groups, 20 µg of the EcN-derived OMV samples was diluted in 500 µL of PBS and mixed with 0.5 µL of DiO membrane dye. All reaction mixtures underwent purification using an ultrafiltration centrifuge tube (100kDa) at 6000 rpm for 30 min to remove any unbound dye; two purification cycles were performed for each sample. Then, the precipitate was resuspended in 200 µL of PBS. RAW264.7 macrophages were co-cultured with the purified DiO-labeled EcN-derived OMVs of varying sizes for 16 h. The RAW264.7 macrophages were washed three times with PBS, followed by nuclear staining with Hoechst 33342 for 5 min. After washing with PBS, the RAW264.7 macrophages were added to the cell culture medium for fluorescence imaging.
2.11. Effects of EcN-Derived OMVs of Varying Sizes on Inflammatory Cytokine Secretion by RAW264.7 Macrophages
RAW264.7 macrophages were seeded into a 24-well plate at a concentration of 1 × 105/well. The cells were cultured at 37 °C and 5% CO2 until 70–80% confluence. A 0.95 mL aliquot of culture medium was added to each well to replace the extracted culture medium, and 50 µL aliquots of EcN-derived OMVs of different sizes at standardized concentrations of 80 µg/mL were added to final concentrations of 4 µg/mL in each well. After the RAW264.7 cells were cocultured with different sizes of EcN-derived OMVs for 24 h, the culture media were collected and centrifuged at 1500× g for 10 min at 4 °C, and the supernatants were collected. ELISA kits (Sevier Biotechnology, Wuhan, China) were used to measure the concentrations of IL-1β and IL-10 according to the manufacturer’s instructions.
2.12. Statistical Analysis
Statistical analyses were performed via one-way ANOVA. All the data are presented as the mean ± SDs of three independent experiments. Significance was assessed at p < 0.05 (*), p < 0.01 (**), and p < 0.001 (***). All the statistical analyses were performed via GraphPad Prism software 5.0 (San Diego, CA, USA).
4. Discussion
OMVs are extracellular vesicles secreted by Gram-negative bacteria with diameters ranging from 20 to 250 nm. OMVs contain biologically active molecules such as proteins, lipids, and nucleic acids, which are secreted by parent bacteria [
35]. Since OMVs can transport substances derived from their parent bacteria, they may serve as effective delivery vehicles for transferring bacterial components to host cells during interactions. This mechanism has the potential to induce an immune response in the host [
36]. As a novel secretion system, OMVs have many advantages. First, OMVs can selectively encapsulate the contents of parent bacteria, such as lipids, proteins, and LPS. Second, OMVs can protect their contents from being degraded by extracellular enzymes, ensuring that they can safely reach target cells. Finally, OMVs are equipped with surface ligands that interact with host cell receptors, thereby directly eliciting host immune responses. OMVs have significant potential for application in vaccine research and development, drug delivery systems, and antitumor therapeutics [
2,
6]. Regarding their use in vaccines, OMVs are highly immunogenic but cannot replicate; thus, they can be used as vaccine antigen candidates as well as vaccine adjuvants [
37,
38]. Probiotic-derived OMVs can be used as drug carriers to prevent and treat corresponding diseases; they are inanimate substances that benefit the host’s health and can avoid potential harm to the host caused by live bacteria while retaining the beneficial and immunogenic properties of the parent bacteria. Regarding antitumor preparations, when combined with other treatment modalities, OMVs can enhance tumor immunotherapy and prevent tumor recurrence and metastasis [
39]. EcN is a Gram-negative probiotic that is widely used in clinical treatment because of its ability to significantly alleviate intestinal diseases [
40]. OMVs secreted by EcN serve as carriers of signaling molecules and can trigger pro-inflammatory and anti-inflammatory responses in RAW264.7 macrophages, thereby regulating the biological functions of these cells [
26,
33]. Nevertheless, the heterogeneity of EcN-derived OMVs of varying sizes has received limited scholarly attention.
Presently, frequently employed techniques for separating OMVs include ultracentrifugation, size-exclusion chromatography (SEC), ultrafiltration, and density gradient centrifugation [
5,
41]. Each of these methods presents distinct advantages and disadvantages. Notably, ultracentrifugation is a widely utilized technique that separates OMVs from cellular debris, bacteria, and other contaminants on the basis of the applied centrifugal force. However, this method is associated with high equipment costs, and the high centrifugal force may damage the structural integrity and functionality of the OMVs. The SEC method achieves the separation of particles on the basis of size through utilizing porous polymer gel packing. While the SEC method offers high separation purity, it is primarily suitable for the purification of samples with small volumes. In contrast, the ultrafiltration technique employs membranes with varying molecular weight cutoffs to selectively separate and collect vesicles. Ultrafiltration has the highest recovery efficiency. Density gradient centrifugation facilitates the distribution of OMVs across specific density layers via ultra-high-speed centrifugation. This method achieves the highest separation purity, but the operation is cumbersome. This study investigated the heterogeneity of EcN-derived OMVs of different sizes. The main objective was to efficiently obtain EcN-derived OMV samples of different sizes that satisfied the experimental requirements. Conventional methods fail to produce EcN-derived OMVs of different sizes. Inspired by the experimental principles of ultrafiltration, we developed a membrane filtration method to obtain EcN-derived OMVs of varying sizes. In this study, we first used the gradient filtration previously developed by our research group specifically for the separation of EcN-derived OMVs, and used SEC to assess the purity of the EcN-derived OMV samples. The experimental findings indicated that the purity of the EcN-derived OMVs obtained via gradient filtration was comparable to that achieved through SEC. Our approach to optimizing the gradient filtration method, through the incorporation of pretreatment steps and the repeated washing of the samples, proved to be highly effective. We then used the membrane filtration method to separate the high-purity isolated EcN-derived OMVs into three major size groups (<50 nm, 50–100 nm, and 100–300 nm), which were confirmed using TEM and DLS. The significant advantages of the membrane filtration method are that it is fast, convenient, and suitable for achieving industrial-scale yields for mass production, and it maintains the biological functional activity of the isolated OMVs.
OMVs contain biologically active molecules, including proteins, lipids, polysaccharides, DNA, and RNA derived from parent bacterial cells. Elucidating the molecular composition of OMVs is crucial for comprehending their function in bacterial growth or interaction with the host. The analysis of OMV components mainly employs proteomic technologies to elucidate their protein composition. Proteomic research on the OMVs of Gram-negative pathogenic bacteria has identified many protein constituents associated with bacterial pathogenesis and immune modulation [
42,
43]. In this study, our proteomics results revealed that the isolated EcN-derived OMVs contained 646 total protein types. There is a paucity of published research concerning proteomic analyses of EcN-derived OMVs of different sizes. Nice et al. reported that the
Aggregatibacter actinomycetemcomitans strain JP2 produces two types of vesicles throughout growth, with varying sizes and toxin compositions [
44]. Turner et al. reported that the size of
Helicobacter pylori-derived OMVs determines their protein content, as fewer and less diverse bacterial proteins are contained within smaller OMVs than those contained within larger OMVs [
45]. In addition, there are related reports on the heterogeneity of EVs derived from animal cells. Extracellular vesicles (EVs) released by tumor cells have received considerable attention in recent years, as they contain biomarkers such as tumor-specific proteins and nucleic acids (i.e., RNA and DNA) [
46,
47]. Liu et al. researched the effect of EV size on protein profiles. They isolated EVs of different sizes from cultured media with 22Rv1 prostate cancer cells via ExoTIC devices with two key pore sizes (30 and 50 nm), and reported that EVs of different sizes contained different protein types [
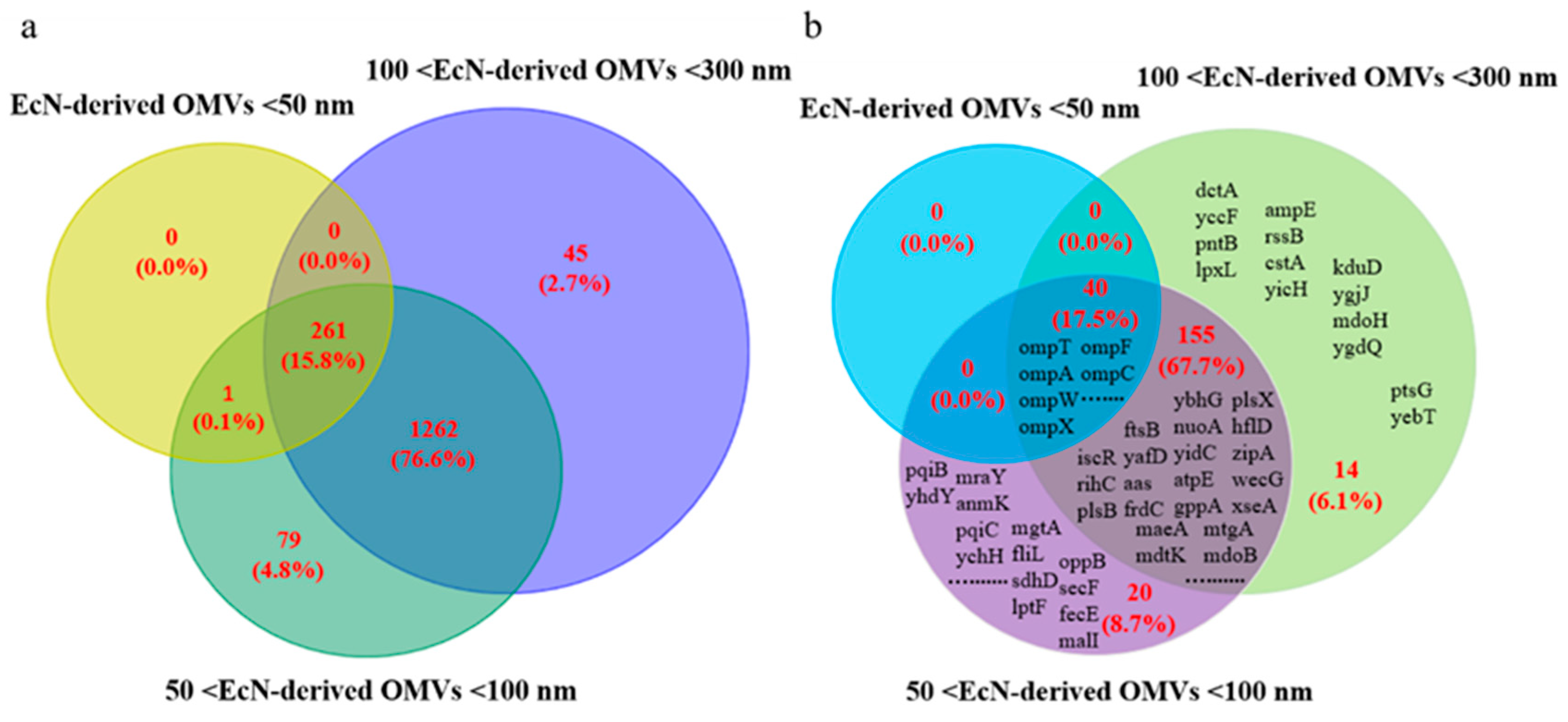
34]. In this study, we used LC-MS/MS analysis to perform proteomic analyses of EcN-derived OMVs of three specific sizes: <50 nm, 50–100 nm, and 100–300 nm. Our results revealed that the <50 nm group contained significantly fewer protein types (262) than the 50–100 nm (1603) and 100–300 nm (1568) groups did. We conducted a categorical analysis of 261 proteins commonly present in EcN-derived OMVs of three specific sizes. They were classified into five groups: cytoplasmic (44.1%, 115/261), periplasmic (32.2%, 84/261), outer membrane (13.8%, 36/261), inner membrane (1.5%, 4/261), and extracellular (8.4%, 22/261). Given that the EcN-derived OMVs of three specific sizes were all secreted by the parent bacteria, it was entirely plausible that they shared certain common proteins. The results indicated that the EcN-derived OMVs of three specific sizes possessed a relatively rich variety of common proteins, which suggested that they might perform similar biological functions. As we found in this study, the EcN-derived OMVs of three specific sizes had obvious therapeutic effects on RAW264.7 macrophages and significantly promoted the secretion of the pro-inflammatory factor IL-1β and the anti-inflammatory factor IL-10 by RAW264.7 macrophages. Although our study provides limited proteomic data on EcN-derived OMVs of three distinct sizes, these experimental findings substantiate that EcN-derived OMVs of varying sizes present differences in protein composition. Future research will involve a comprehensive analysis of proteomic data from EcN-derived OMVs of different sizes, with a focus on investigating the types and characteristics of these differential proteins in greater depth.
Most inflammatory responses result from innate immunity, which is mainly caused through the activation of pro-inflammatory signaling cascades by macrophages and leukocytes. The inflammatory cytokines produced by this cascade comprise small soluble proteins produced by these immune cells; these cytokines regulate the host’s immune and inflammatory responses. Inflammatory cytokines involved in inflammatory responses are mainly divided into two categories, namely, pro-inflammatory factors (e.g., TNF-α, IL-6, and IL-1β) and anti-inflammatory factors (e.g., IL-4 and IL-10) [
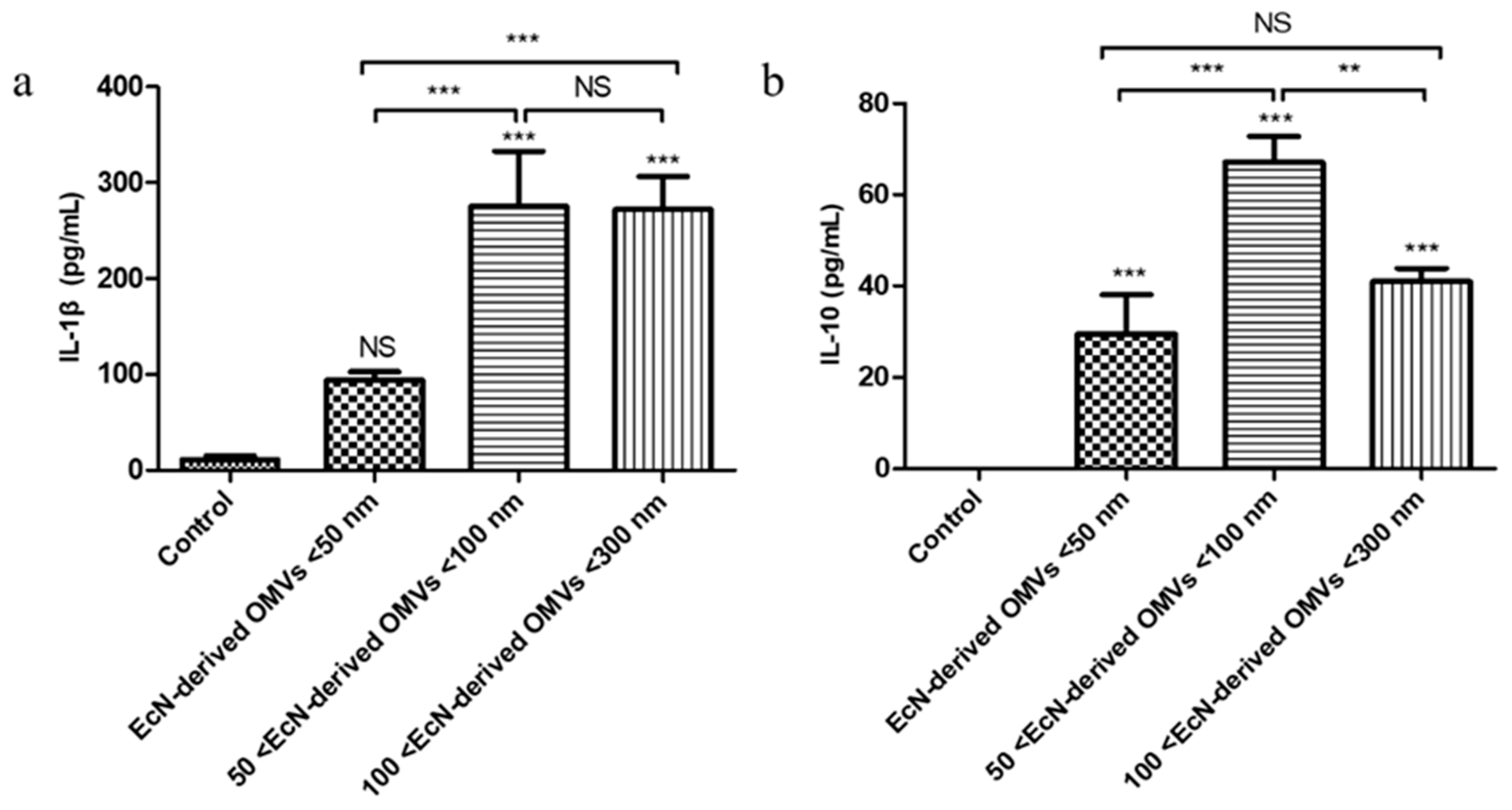
48]. Among them, IL-1β can stimulate immune cells to secrete pro-inflammatory factors such as TNF-α, which plays a pro-inflammatory role. TNF-α is an important regulatory factor in the initiation of immunity and inflammation. IL-10 is an anti-inflammatory factor that can play an anti-inflammatory role. EcN-derived OMVs can stimulate RAW264.7 macrophages to secrete the pro-inflammatory factors TNF-α, IL-6, and IL-1β, and the anti-inflammatory factor IL-10 [
26]. In this study, the concentrations of the pro-inflammatory cytokine IL-1β produced by RAW264.7 macrophages in the three types of EcN-derived OMVs of different sizes (<50 nm, 50–100 nm, and 100–300 nm) were 93.7 ± 8.6 pg/mL, 274.9 ± 58.1 pg/mL, and 271.9 ± 34.1 pg/mL, respectively. Moreover, the concentration of anti-inflammatory IL-10 produced by RAW264.7 macrophages in the 50–100 nm group was the highest (67.1 ± 5.7 pg/mL), and the concentrations of IL-10 in the <50 nm and 100–300 nm groups were 29.5 ± 8.6 pg/mL and 40.9 ± 2.9 pg/mL, respectively. Our results revealed that the therapeutic effects of EcN-derived OMVs of different sizes differed; the 50–100 nm group had a stronger therapeutic effect than the other two groups. We analyzed the reasons for the different experimental phenomena observed regarding the macrophage function mentioned above and speculate that these differences may be attributable to the distinct protein compositions of EcN-derived OMVs of varying sizes. Proteomic analyses of the EcN-derived OMVs of three specific sizes revealed that, in addition to the 261 common proteins shared among the three groups, the 50–100 nm group contained 1341 unique proteins that were distinct from those in the <50 nm group. Similarly, the 100–300 nm group contained 1307 unique proteins that were distinct from those in the <50 nm group. These experimental results suggest that the 50–100 nm and 100–300 nm groups might have stronger therapeutic effects on RAW264.7 macrophages than the <50 nm group. Furthermore, a comparative analysis between the 50–100 nm group and the 100–300 nm group revealed that, in addition to sharing 1523 common proteins, each group also contained distinct proteins. The 50–100 nm group contained 80 unique proteins, whereas the 100–300 nm group contained 45 unique proteins. This variation in protein composition within each group might account for the differential therapeutic effects observed between the 50–100 nm group and the 100–300 nm group on RAW264.7 macrophages. In summary, these differences in macrophage function are consistent with the differences in the proteomic information of the EcN-derived OMVs of varying sizes that we measured. In the macrophage function experiment, we only measured changes in the representative pro-inflammatory cytokine IL-1β and the anti-inflammatory cytokine IL-10. Subsequent experiments should be conducted to further analyze the changes in other pro-inflammatory factors, such as TNF-α, IL-6, and other anti-inflammatory factors, such as IL-4.
Although our experimental findings suggest that EcN-derived OMVs of varying sizes are heterogeneous in terms of protein composition and interactions with RAW264.7 macrophages, additional research is necessary to establish more comprehensive conclusions. Our preliminary proteomic results suggest that EcN-derived OMVs of different sizes present distinct protein compositions. The composition of other components in EcN-derived OMVs of varying sizes, such as lipopolysaccharides, phospholipids, DNA, and RNA, is currently unknown. This study aimed to elucidate the differences in the therapeutic effects of EcN-derived OMVs of varying sizes on RAW264.7 macrophages through examining the distinct protein compositions associated with these EcN-derived OMVs. Importantly, this may represent only one contributing factor. The variations in other constituents contained in EcN-derived OMVs of different sizes may also influence their effects on RAW264.7 macrophages. Further investigation into these unidentified components is necessary in future research. In addition, the therapeutic efficacy of EcN-derived OMVs of varying sizes on macrophages in mouse models remains unclear and requires further validation. However, notwithstanding these limitations, our results provide direct experimental evidence that EcN-derived OMVs of different sizes exhibit heterogeneous properties.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}