

Molecular Dynamics Simulations of the Proteins Regulating Synaptic Vesicle Fusion

Abstract
1. Introduction
2. The SNARE Complex Assembly
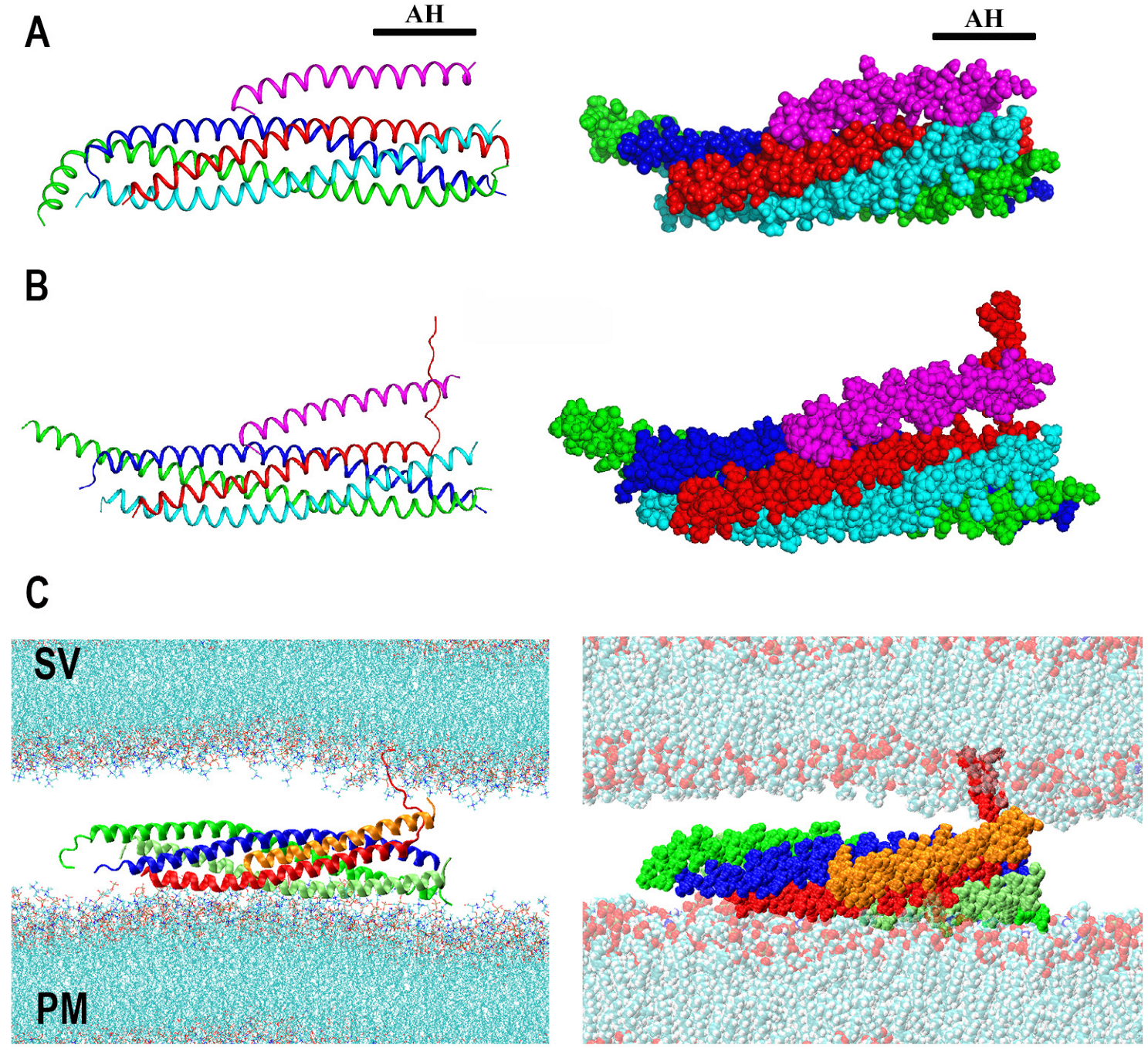
3. Cpx as a Dynamic Fusion Clamp

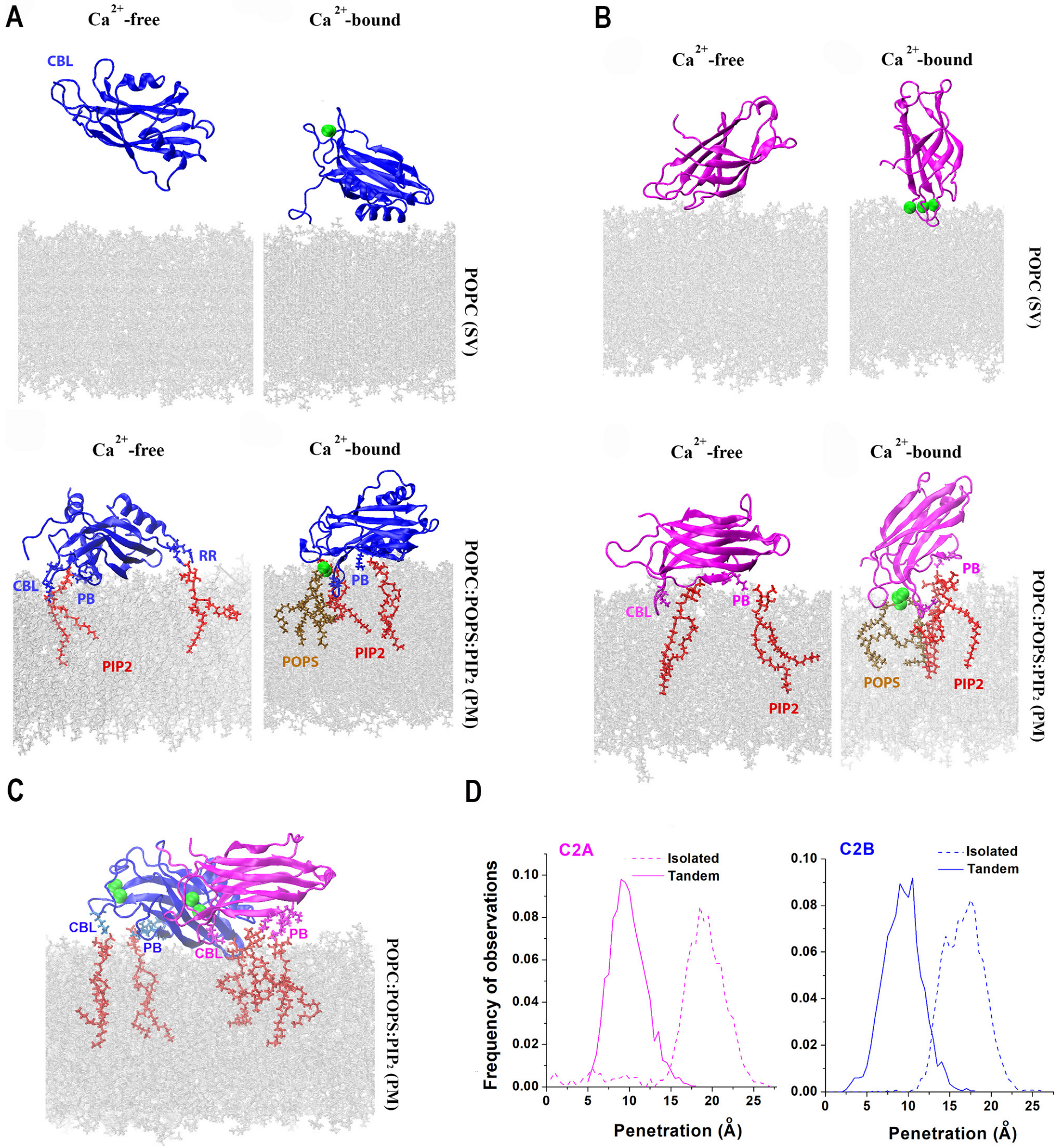
4. Syt1 and Its Interaction with Lipid Bilayers
5. The Prefusion Syt1-SNARE-Cpx Complex
6. Conclusions and Further Directions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Sudhof, T.C.; Rothman, J.E. Membrane fusion: Grappling with SNARE and SM proteins. Science 2009, 323, 474–477. [Google Scholar] [CrossRef] [PubMed]
- Sudhof, T.C. Neurotransmitter release: The last millisecond in the life of a synaptic vesicle. Neuron 2013, 80, 675–690. [Google Scholar] [CrossRef]
- Kaeser, P.S.; Regehr, W.G. Molecular mechanisms for synchronous, asynchronous, and spontaneous neurotransmitter release. Annu. Rev. Physiol. 2014, 76, 333–363. [Google Scholar] [CrossRef] [PubMed]
- Rizo, J.; Xu, J. The Synaptic Vesicle Release Machinery. Annu. Rev. Biophys. 2015, 44, 339–367. [Google Scholar] [CrossRef] [PubMed]
- Chapman, E.R. How does synaptotagmin trigger neurotransmitter release? Annu. Rev. Biochem. 2008, 77, 615–641. [Google Scholar] [CrossRef]
- Leitz, J.; Kavalali, E.T. Ca2+ Dependence of Synaptic Vesicle Endocytosis. Neuroscientist 2015, 22, 464–476. [Google Scholar] [CrossRef]
- Perin, M.S.; Brose, N.; Jahn, R.; Sudhof, T.C. Domain structure of synaptotagmin (p65). J. Biol. Chem. 1991, 266, 623–629. [Google Scholar] [CrossRef]
- Ubach, J.; Zhang, X.; Shao, X.; Sudhof, T.C.; Rizo, J. Ca2+ binding to synaptotagmin: How many Ca2+ ions bind to the tip of a C2-domain? EMBO J. 1998, 17, 3921–3930. [Google Scholar] [CrossRef]
- Fernandez-Chacon, R.; Konigstorfer, A.; Gerber, S.H.; Garcia, J.; Matos, M.F.; Stevens, C.F.; Brose, N.; Rizo, J.; Rosenmund, C.; Sudhof, T.C. Synaptotagmin I functions as a calcium regulator of release probability. Nature 2001, 410, 41–49. [Google Scholar] [CrossRef]
- Nyenhuis, S.B.; Karandikar, N.; Kiessling, V.; Kreutzberger, A.J.B.; Thapa, A.; Liang, B.; Tamm, L.K.; Cafiso, D.S. Conserved arginine residues in synaptotagmin 1 regulate fusion pore expansion through membrane contact. Nat. Commun. 2021, 12, 761. [Google Scholar] [CrossRef]
- Martens, S.; Kozlov, M.M.; McMahon, H.T. How synaptotagmin promotes membrane fusion. Science 2007, 316, 1205–1208. [Google Scholar] [CrossRef] [PubMed]
- Lynch, K.L.; Gerona, R.R.; Kielar, D.M.; Martens, S.; McMahon, H.T.; Martin, T.F. Synaptotagmin-1 utilizes membrane bending and SNARE binding to drive fusion pore expansion. Mol. Biol. Cell 2008, 19, 5093–5103. [Google Scholar] [CrossRef] [PubMed]
- Schupp, M.; Malsam, J.; Ruiter, M.; Scheutzow, A.; Wierda, K.D.; Sollner, T.H.; Sorensen, J.B. Interactions Between SNAP-25 and Synaptotagmin-1 Are Involved in Vesicle Priming, Clamping Spontaneous and Stimulating Evoked Neurotransmission. J. Neurosci. 2016, 36, 11865–11880. [Google Scholar] [CrossRef] [PubMed]
- Pang, Z.P.; Shin, O.H.; Meyer, A.C.; Rosenmund, C.; Sudhof, T.C. A gain-of-function mutation in synaptotagmin-1 reveals a critical role of Ca2+-dependent soluble N-ethylmaleimide-sensitive factor attachment protein receptor complex binding in synaptic exocytosis. J. Neurosci. 2006, 26, 12556–12565. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Lai, Y.; Bacaj, T.; Zhao, M.; Lyubimov, A.Y.; Uervirojnangkoorn, M.; Zeldin, O.B.; Brewster, A.S.; Sauter, N.K.; Cohen, A.E.; et al. Architecture of the synaptotagmin-SNARE machinery for neuronal exocytosis. Nature 2015, 525, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Guan, Z.; Bykhovskaia, M.; Jorquera, R.A.; Sutton, R.B.; Akbergenova, Y.; Littleton, J.T. A synaptotagmin suppressor screen indicates SNARE binding controls the timing and Ca(2+) cooperativity of vesicle fusion. Elife 2017, 6, e28409. [Google Scholar] [CrossRef]
- Brewer, K.D.; Bacaj, T.; Cavalli, A.; Camilloni, C.; Swarbrick, J.D.; Liu, J.; Zhou, A.; Zhou, P.; Barlow, N.; Xu, J.; et al. Dynamic binding mode of a Synaptotagmin-1-SNARE complex in solution. Nat. Struct. Mol. Biol. 2015, 22, 555–564. [Google Scholar] [CrossRef]
- Jahn, R.; Fasshauer, D. Molecular machines governing exocytosis of synaptic vesicles. Nature 2012, 490, 201–207. [Google Scholar] [CrossRef]
- Park, Y.; Seo, J.B.; Fraind, A.; Perez-Lara, A.; Yavuz, H.; Han, K.; Jung, S.R.; Kattan, I.; Walla, P.J.; Choi, M.; et al. Synaptotagmin-1 binds to PIP(2)-containing membrane but not to SNAREs at physiological ionic strength. Nat. Struct. Mol. Biol. 2015, 22, 815–823. [Google Scholar] [CrossRef]
- Chen, X.; Tomchick, D.R.; Kovrigin, E.; Arac, D.; Machius, M.; Sudhof, T.C.; Rizo, J. Three-dimensional structure of the complexin/SNARE complex. Neuron 2002, 33, 397–409. [Google Scholar] [CrossRef]
- Reim, K.; Mansour, M.; Varoqueaux, F.; McMahon, H.T.; Sudhof, T.C.; Brose, N.; Rosenmund, C. Complexins regulate a late step in Ca2+-dependent neurotransmitter release. Cell 2001, 104, 71–81. [Google Scholar] [CrossRef]
- Xue, M.; Lin, Y.Q.; Pan, H.; Reim, K.; Deng, H.; Bellen, H.J.; Rosenmund, C. Tilting the balance between facilitatory and inhibitory functions of mammalian and Drosophila Complexins orchestrates synaptic vesicle exocytosis. Neuron 2009, 64, 367–380. [Google Scholar] [CrossRef]
- Chang, S.; Reim, K.; Pedersen, M.; Neher, E.; Brose, N.; Taschenberger, H. Complexin stabilizes newly primed synaptic vesicles and prevents their premature fusion at the mouse calyx of held synapse. J. Neurosci. 2015, 35, 8272–8290. [Google Scholar] [CrossRef]
- Huntwork, S.; Littleton, J.T. A complexin fusion clamp regulates spontaneous neurotransmitter release and synaptic growth. Nat. Neurosci. 2007, 10, 1235–1237. [Google Scholar] [CrossRef] [PubMed]
- Trimbuch, T.; Rosenmund, C. Should I stop or should I go? The role of complexin in neurotransmitter release. Nat. Rev. Neurosci. 2016, 17, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Jorquera, R.A.; Huntwork-Rodriguez, S.; Akbergenova, Y.; Cho, R.W.; Littleton, J.T. Complexin controls spontaneous and evoked neurotransmitter release by regulating the timing and properties of synaptotagmin activity. J. Neurosci. 2012, 32, 18234–18245. [Google Scholar] [CrossRef]
- Dhara, M.; Yarzagaray, A.; Schwarz, Y.; Dutta, S.; Grabner, C.; Moghadam, P.K.; Bost, A.; Schirra, C.; Rettig, J.; Reim, K.; et al. Complexin synchronizes primed vesicle exocytosis and regulates fusion pore dynamics. J. Cell. Biol. 2014, 204, 1123–1140. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Brewer, K.D.; Perez-Castillejos, R.; Rizo, J. Subtle Interplay between synaptotagmin and complexin binding to the SNARE complex. J. Mol. Biol. 2013, 425, 3461–3475. [Google Scholar] [CrossRef]
- Tang, J.; Maximov, A.; Shin, O.H.; Dai, H.; Rizo, J.; Sudhof, T.C. A complexin/synaptotagmin 1 switch controls fast synaptic vesicle exocytosis. Cell 2006, 126, 1175–1187. [Google Scholar] [CrossRef]
- Cao, P.; Yang, X.; Sudhof, T.C. Complexin activates exocytosis of distinct secretory vesicles controlled by different synaptotagmins. J. Neurosci. 2013, 33, 1714–1727. [Google Scholar] [CrossRef]
- Zhou, Q.; Zhou, P.; Wang, A.L.; Wu, D.; Zhao, M.; Sudhof, T.C.; Brunger, A.T. The primed SNARE-complexin-synaptotagmin complex for neuronal exocytosis. Nature 2017, 548, 420–425. [Google Scholar] [CrossRef] [PubMed]
- Giraudo, C.G.; Eng, W.S.; Melia, T.J.; Rothman, J.E. A clamping mechanism involved in SNARE-dependent exocytosis. Science 2006, 313, 676–680. [Google Scholar] [CrossRef] [PubMed]
- Giraudo, C.G.; Garcia-Diaz, A.; Eng, W.S.; Chen, Y.; Hendrickson, W.A.; Melia, T.J.; Rothman, J.E. Alternative zippering as an on-off switch for SNARE-mediated fusion. Science 2009, 323, 512–516. [Google Scholar] [CrossRef] [PubMed]
- Cho, R.W.; Kummel, D.; Li, F.; Baguley, S.W.; Coleman, J.; Rothman, J.E.; Littleton, J.T. Genetic analysis of the Complexin trans-clamping model for cross-linking SNARE complexes in vivo. Proc. Natl. Acad. Sci. USA 2014, 111, 10317–10322. [Google Scholar] [CrossRef]
- Xue, M.; Reim, K.; Chen, X.; Chao, H.T.; Deng, H.; Rizo, J.; Brose, N.; Rosenmund, C. Distinct domains of complexin I differentially regulate neurotransmitter release. Nat. Struct. Mol. Biol. 2007, 14, 949–958. [Google Scholar] [CrossRef]
- Krishnakumar, S.S.; Li, F.; Coleman, J.; Schauder, C.M.; Kummel, D.; Pincet, F.; Rothman, J.E.; Reinisch, K.M. Re-visiting the trans insertion model for complexin clamping. Elife 2015, 4, e04463. [Google Scholar] [CrossRef] [PubMed]
- Krishnakumar, S.S.; Li, F.; Coleman, J.; Schauder, C.M.; Kummel, D.; Pincet, F.; Rothman, J.E.; Reinisch, K.M. Correction: Re-visiting the trans insertion model for complexin clamping. Elife 2017, 6, e31512. [Google Scholar] [CrossRef]
- Krishnakumar, S.S.; Radoff, D.T.; Kummel, D.; Giraudo, C.G.; Li, F.; Khandan, L.; Baguley, S.W.; Coleman, J.; Reinisch, K.M.; Pincet, F.; et al. A conformational switch in complexin is required for synaptotagmin to trigger synaptic fusion. Nat. Struct. Mol. Biol. 2011, 18, 934–940. [Google Scholar] [CrossRef]
- Kummel, D.; Krishnakumar, S.S.; Radoff, D.T.; Li, F.; Giraudo, C.G.; Pincet, F.; Rothman, J.E.; Reinisch, K.M. Complexin cross-links prefusion SNAREs into a zigzag array. Nat. Struct. Mol. Biol. 2011, 18, 927–933. [Google Scholar] [CrossRef]
- Radoff, D.T.; Dong, Y.; Snead, D.; Bai, J.; Eliezer, D.; Dittman, J.S. The accessory helix of complexin functions by stabilizing central helix secondary structure. Elife 2014, 3, e04553. [Google Scholar] [CrossRef]
- Chapman, E.R. A Ca(2+) Sensor for Exocytosis. Trends. Neurosci. 2018, 41, 327–330. [Google Scholar] [CrossRef]
- Brunger, A.T.; Choi, U.B.; Lai, Y.; Leitz, J.; White, K.I.; Zhou, Q. The pre-synaptic fusion machinery. Curr. Opin. Struct. Biol. 2019, 54, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Rizo, J. Mechanism of neurotransmitter release coming into focus. Protein. Sci. 2018, 27, 1364–1391. [Google Scholar] [CrossRef] [PubMed]
- Brunger, A.T.; Leitz, J.; Zhou, Q.; Choi, U.B.; Lai, Y. Ca(2+)-Triggered Synaptic Vesicle Fusion Initiated by Release of Inhibition. Trends. Cell Biol. 2018, 28, 631–645. [Google Scholar] [CrossRef] [PubMed]
- Mohrmann, R.; Dhara, M.; Bruns, D. Complexins: Small but capable. Cell. Mol. Life Sci. 2015, 72, 4221–4235. [Google Scholar] [CrossRef] [PubMed]
- Rizo, J. Molecular Mechanisms Underlying Neurotransmitter Release. Annu. Rev. Biophys. 2022, 51, 377–408. [Google Scholar] [CrossRef]
- Sutton, R.B.; Fasshauer, D.; Jahn, R.; Brunger, A.T. Crystal structure of a SNARE complex involved in synaptic exocytosis at 2.4 A resolution. Nature 1998, 395, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Durrieu, M.P.; Lavery, R.; Baaden, M. Interactions between neuronal fusion proteins explored by molecular dynamics. Biophys. J. 2008, 94, 3436–3446. [Google Scholar] [CrossRef]
- Bock, L.V.; Hutchings, B.; Grubmuller, H.; Woodbury, D.J. Chemomechanical regulation of SNARE proteins studied with molecular dynamics simulations. Biophys. J. 2010, 99, 1221–1230. [Google Scholar] [CrossRef]
- Knecht, V.; Grubmuller, H. Mechanical coupling via the membrane fusion SNARE protein syntaxin 1A: A molecular dynamics study. Biophys. J. 2003, 84, 1527–1547. [Google Scholar] [CrossRef]
- Blanchard, A.E.; Arcario, M.J.; Schulten, K.; Tajkhorshid, E. A highly tilted membrane configuration for the prefusion state of synaptobrevin. Biophys. J. 2014, 107, 2112–2121. [Google Scholar] [CrossRef] [PubMed]
- Delalande, O.; Ferey, N.; Grasseau, G.; Baaden, M. Complex molecular assemblies at hand via interactive simulations. J. Comput. Chem. 2009, 30, 2375–2387. [Google Scholar] [CrossRef] [PubMed]
- Lindau, M.; Hall, B.A.; Chetwynd, A.; Beckstein, O.; Sansom, M.S. Coarse-grain simulations reveal movement of the synaptobrevin C-terminus in response to piconewton forces. Biophys. J. 2012, 103, 959–969. [Google Scholar] [CrossRef] [PubMed]
- Fang, Q.; Zhao, Y.; Herbst, A.D.; Kim, B.N.; Lindau, M. Positively Charged Amino Acids at the SNAP-25 C Terminus Determine Fusion Rates, Fusion Pore Properties, and Energetics of Tight SNARE Complex Zippering. J. Neurosci. 2015, 35, 3230–3239. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Pluhackova, K.; Wassenaar, T.A.; Bockmann, R.A. Synaptobrevin Transmembrane Domain Dimerization Studied by Multiscale Molecular Dynamics Simulations. Biophys. J. 2015, 109, 760–771. [Google Scholar] [CrossRef]
- Marrink, S.J.; Risselada, H.J.; Yefimov, S.; Tieleman, D.P.; de Vries, A.H. The MARTINI force field: Coarse grained model for biomolecular simulations. J. Phys. Chem. B 2007, 111, 7812–7824. [Google Scholar] [CrossRef]
- Sharma, S.; Lindau, M. Molecular mechanism of fusion pore formation driven by the neuronal SNARE complex. Proc. Natl. Acad. Sci. USA 2018, 115, 12751–12756. [Google Scholar] [CrossRef]
- Mostafavi, H.; Thiyagarajan, S.; Stratton, B.S.; Karatekin, E.; Warner, J.M.; Rothman, J.E.; O’Shaughnessy, B. Entropic forces drive self-organization and membrane fusion by SNARE proteins. Proc. Natl. Acad. Sci. USA 2017, 114, 5455–5460. [Google Scholar] [CrossRef]
- Fortoul, N.; Bykhovskaia, M.; Jagota, A. Coarse-Grained Model for Zippering of SNARE from Partially Assembled States. J. Phys. Chem. B. 2018, 122, 10834–10840. [Google Scholar] [CrossRef]
- Fortoul, N.; Singh, P.; Hui, C.-Y.; Bykhovskaia, M.; Jagota, A. Coarse-Grained Model of the Snare Complex Determines the Number of Snares Required for Docking. Biophys. J. 2015, 108, 154a. [Google Scholar] [CrossRef]
- McDargh, Z.A.; Polley, A.; O’Shaughnessy, B. SNARE-mediated membrane fusion is a two-stage process driven by entropic forces. FEBS. Lett. 2018, 592, 3504–3515. [Google Scholar] [CrossRef] [PubMed]
- Ayton, G.S.; Voth, G.A. Multiscale simulation of protein mediated membrane remodeling. Semin. Cell Dev. Biol. 2010, 21, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Ayton, G.S.; Tepper, H.L.; Mirijanian, D.T.; Voth, G.A. A new perspective on the coarse-grained dynamics of fluids. J. Chem. Phys. 2004, 120, 4074–4088. [Google Scholar] [CrossRef] [PubMed]
- Ayton, G.S.; Voth, G.A. Systematic multiscale simulation of membrane protein systems. Curr. Opin. Struct. Biol. 2009, 19, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, G.; Nandekar, P.P.; Yu, X.; Wade, R.C. On the application of the MARTINI coarse-grained model to immersion of a protein in a phospholipid bilayer. J. Chem. Phys. 2015, 143, 243139. [Google Scholar] [CrossRef]
- Lamberg, A.; Taniguchi, T. Coarse-grained computational studies of supported bilayers: Current problems and their root causes. J. Phys. Chem. B. 2014, 118, 10643–10652. [Google Scholar] [CrossRef]
- Bao, H.; Das, D.; Courtney, N.A.; Jiang, Y.; Briguglio, J.S.; Lou, X.; Roston, D.; Cui, Q.; Chanda, B.; Chapman, E.R. Dynamics and number of trans-SNARE complexes determine nascent fusion pore properties. Nature 2018, 554, 260–263. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Shen, Q.T.; Kiel, A.; Wang, J.; Wang, H.W.; Melia, T.J.; Rothman, J.E.; Pincet, F. SNARE proteins: One to fuse and three to keep the nascent fusion pore open. Science 2012, 335, 1355–1359. [Google Scholar] [CrossRef]
- Sinha, R.; Ahmed, S.; Jahn, R.; Klingauf, J. Two synaptobrevin molecules are sufficient for vesicle fusion in central nervous system synapses. Proc. Natl. Acad. Sci. USA 2011, 108, 14318–14323. [Google Scholar] [CrossRef]
- Van den Bogaart, G.; Holt, M.G.; Bunt, G.; Riedel, D.; Wouters, F.S.; Jahn, R. One SNARE complex is sufficient for membrane fusion. Nat. Struct. Mol. Biol. 2010, 17, 358–364. [Google Scholar] [CrossRef]
- Zdanowicz, R.; Kreutzberger, A.; Liang, B.; Kiessling, V.; Tamm, L.K.; Cafiso, D.S. Complexin Binding to Membranes and Acceptor t-SNAREs Explains Its Clamping Effect on Fusion. Biophys. J. 2017, 113, 1235–1250. [Google Scholar] [CrossRef] [PubMed]
- Bykhovskaia, M.; Jagota, A.; Gonzalez, A.; Vasin, A.; Littleton, J.T. Interaction of the complexin accessory helix with the C-terminus of the SNARE complex: Molecular-dynamics model of the fusion clamp. Biophys. J. 2013, 105, 679–690. [Google Scholar] [CrossRef] [PubMed]
- Brady, J.; Vasin, A.; Bykhovskaia, M. The Accessory Helix of Complexin Stabilizes a Partially Unzippered State of the SNARE Complex and Mediates the Complexin Clamping Function In Vivo. eNeuro 2021, 8, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Vasin, A.; Volfson, D.; Littleton, J.T.; Bykhovskaia, M. Interaction of the Complexin Accessory Helix with Synaptobrevin Regulates Spontaneous Fusion. Biophys. J. 2016, 111, 1954–1964. [Google Scholar] [CrossRef]
- Bykhovskaia, M. Molecular Dynamics Simulations of the SNARE Complex. Methods. Mol. Biol. 2019, 1860, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Fuson, K.L.; Montes, M.; Robert, J.J.; Sutton, R.B. Structure of human synaptotagmin 1 C2AB in the absence of Ca2+ reveals a novel domain association. Biochemistry 2007, 46, 13041–13048. [Google Scholar] [CrossRef]
- Herrick, D.Z.; Kuo, W.; Huang, H.; Schwieters, C.D.; Ellena, J.F.; Cafiso, D.S. Solution and membrane-bound conformations of the tandem C2A and C2B domains of synaptotagmin 1: Evidence for bilayer bridging. J. Mol. Biol. 2009, 390, 913–923. [Google Scholar] [CrossRef]
- Kuo, W.; Herrick, D.Z.; Cafiso, D.S. Phosphatidylinositol 4,5-bisphosphate alters synaptotagmin 1 membrane docking and drives opposing bilayers closer together. Biochemistry 2011, 50, 2633–2641. [Google Scholar] [CrossRef]
- Nyenhuis, S.B.; Thapa, A.; Cafiso, D.S. Phosphatidylinositol 4,5 Bisphosphate Controls the cis and trans Interactions of Synaptotagmin 1. Biophys. J. 2019, 117, 247–257. [Google Scholar] [CrossRef]
- Bykhovskaia, M. Calcium binding promotes conformational flexibility of the neuronal Ca(2+) sensor synaptotagmin. Biophys. J. 2015, 108, 2507–2520. [Google Scholar] [CrossRef]
- Wu, Z.; Schulten, K. Synaptotagmin’s role in neurotransmitter release likely involves Ca(2+)-induced conformational transition. Biophys. J. 2014, 107, 1156–1166. [Google Scholar] [CrossRef] [PubMed]
- Prasad, R.; Zhou, H.X. Membrane Association and Functional Mechanism of Synaptotagmin-1 in Triggering Vesicle Fusion. Biophys. J. 2020, 119, 1255–1265. [Google Scholar] [CrossRef]
- Bykhovskaia, M. SNARE complex alters the interactions of the Ca(2+) sensor synaptotagmin 1 with lipid bilayers. Biophys. J. 2021, 120, 642–661. [Google Scholar] [CrossRef] [PubMed]
- Guillen, J.; Ferrer-Orta, C.; Buxaderas, M.; Perez-Sanchez, D.; Guerrero-Valero, M.; Luengo-Gil, G.; Pous, J.; Guerra, P.; Gomez-Fernandez, J.C.; Verdaguer, N.; et al. Structural insights into the Ca2+ and PI(4,5)P2 binding modes of the C2 domains of rabphilin 3A and synaptotagmin 1. Proc. Natl. Acad. Sci. USA 2013, 110, 20503–20508. [Google Scholar] [CrossRef] [PubMed]
- Perez-Lara, A.; Thapa, A.; Nyenhuis, S.B.; Nyenhuis, D.A.; Halder, P.; Tietzel, M.; Tittmann, K.; Cafiso, D.S.; Jahn, R. PtdInsP2 and PtdSer cooperate to trap synaptotagmin-1 to the plasma membrane in the presence of calcium. Elife 2016, 5, e15886. [Google Scholar] [CrossRef]
- Hui, E.; Bai, J.; Chapman, E.R. Ca2+-triggered simultaneous membrane penetration of the tandem C2-domains of synaptotagmin I. Biophys. J. 2006, 91, 1767–1777. [Google Scholar] [CrossRef]
- Wang, P.; Wang, C.T.; Bai, J.; Jackson, M.B.; Chapman, E.R. Mutations in the effector binding loops in the C2A and C2B domains of synaptotagmin I disrupt exocytosis in a nonadditive manner. J. Biol. Chem. 2003, 278, 47030–47037. [Google Scholar] [CrossRef]
- Lin, C.C.; Seikowski, J.; Perez-Lara, A.; Jahn, R.; Hobartner, C.; Walla, P.J. Control of membrane gaps by synaptotagmin-Ca2+ measured with a novel membrane distance ruler. Nat. Commun. 2014, 5, 5859. [Google Scholar] [CrossRef]
- Hui, E.; Johnson, C.P.; Yao, J.; Dunning, F.M.; Chapman, E.R. Synaptotagmin-mediated bending of the target membrane is a critical step in Ca(2+)-regulated fusion. Cell. 2009, 138, 709–721. [Google Scholar] [CrossRef]
- McMahon, H.T.; Kozlov, M.M.; Martens, S. Membrane curvature in synaptic vesicle fusion and beyond. Cell 2010, 140, 601–605. [Google Scholar] [CrossRef]
- Lai, A.L.; Huang, H.; Herrick, D.Z.; Epp, N.; Cafiso, D.S. Synaptotagmin 1 and SNAREs form a complex that is structurally heterogeneous. J. Mol. Biol. 2011, 405, 696–706. [Google Scholar] [CrossRef]
- Rizo, J.; Sari, L.; Qi, Y.; Im, W.; Lin, M.M. All-atom molecular dynamics simulations of Synaptotagmin-SNARE-complexin complexes bridging a vesicle and a flat lipid bilayer. Elife 2022, 11, e76356. [Google Scholar] [CrossRef] [PubMed]
- Shaw, D.E.; Dror, R.O.; Grossman, J.P.; Mackenzie, K.M.; Bank, J.A.; Young, C.; Deneroff, M.M.; Batson, B.; Bowers, K.J.; Chow, E.; et al. Millisecond-Scale Molecular Dynamics Simulations on Anton. In Proceedings of the Conference on High Performance Computing Networking Storage and Analysis; ACM: New York, NY, USA, 2009; Volume SC09. [Google Scholar]
- Dror, R.O.; Jensen, M.O.; Shaw, D.E. Elucidating membrane protein function through long-timescale molecular dynamics simulation. Annu. Int. Conf. IEEE Eng. Med. Biol. Soc. 2009, 2009, 2340–2342. [Google Scholar] [PubMed]
- Shaw, D.E.; Maragakis, P.; Lindorff-Larsen, K.; Piana, S.; Dror, R.O.; Eastwood, M.P.; Bank, J.A.; Jumper, J.M.; Salmon, J.K.; Shan, Y.; et al. Atomic-Level Characterization of the Structural Dynamics of Proteins. Science 2010, 330, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Dror, R.O.; Dirks, R.M.; Grossman, J.; Xu, H.; Shaw, D.E. Biomolecular Simulation: A Computational Microscope for Molecular Biology. Annu. Rev. Biophys. 2012, 41, 429–452. [Google Scholar] [CrossRef]
- Baker, R.W.; Jeffrey, P.D.; Zick, M.; Phillips, B.P.; Wickner, W.T.; Hughson, F.M. A direct role for the Sec1/Munc18-family protein Vps33 as a template for SNARE assembly. Science 2015, 349, 1111–1114. [Google Scholar] [CrossRef]
- Ma, L.; Rebane, A.A.; Yang, G.; Xi, Z.; Kang, Y.; Gao, Y.; Zhang, Y. Munc18-1-regulated stage-wise SNARE assembly underlying synaptic exocytosis. Elife 2015, 4, e09580. [Google Scholar] [CrossRef]
- Montecucco, C.; Schiavo, G.; Pantano, S. SNARE complexes and neuroexocytosis: How many, how close? Trends Biochem. Sci. 2005, 30, 367–372. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Main Focus | Methodology | References |
|---|---|---|
| SNARE bundle dynamics | AAMD | [48,49,52] |
| SNARE TM domains embedded in lipids | AAMD | [50,51] |
| CGMD, Martini force field | [53] | |
| SNARE zippering under the forces exerted by the PM-SV repulsion | CGMD, Martini force field | [52,57] |
| CGMD, customized force fields | [58,59,60,61] | |
| Cpx fusion clamp | AAMD | [72,73,74] |
| Syt1 interdomain rotations | AAMD | [16,80] |
| Syt1 interactions with lipids | AAMD | [81,82,83] |
| Syt1-SNARE-Cpx prefusion complex between PM and SV | AAMD | [83,92] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bykhovskaia, M. Molecular Dynamics Simulations of the Proteins Regulating Synaptic Vesicle Fusion. Membranes 2023, 13, 307. https://doi.org/10.3390/membranes13030307
Bykhovskaia M. Molecular Dynamics Simulations of the Proteins Regulating Synaptic Vesicle Fusion. Membranes. 2023; 13(3):307. https://doi.org/10.3390/membranes13030307
Chicago/Turabian StyleBykhovskaia, Maria. 2023. "Molecular Dynamics Simulations of the Proteins Regulating Synaptic Vesicle Fusion" Membranes 13, no. 3: 307. https://doi.org/10.3390/membranes13030307
APA StyleBykhovskaia, M. (2023). Molecular Dynamics Simulations of the Proteins Regulating Synaptic Vesicle Fusion. Membranes, 13(3), 307. https://doi.org/10.3390/membranes13030307