1. Introduction
Immunoglobulins are widely used in practice [
1,
2]. There is a tendency to clarify the specific changes of erythrocytes when they are affected by hematin or pooled immunoglobulin G (IgG). The activating or damaging effects of pooled IgG or hematin, as well as the action of melittin as a toxic substance to the erythrocyte membrane, and also the combination of these chemical agents are realized in the erythrocyte through the membrane system of the cells. Pooled IgG is indicated for the treatment of primary humoral immunodeficiencies, such as congenital agammaglobulinemia, common variable immunodeficiency, X-linked agammaglobulinemia, Wiskott–Aldrich syndrome and severe combined immunodeficiencies. Melittin, as the major toxic component of the bee venom from
Apis mellifera, is used as a membrane-disruptive agent [
3] causing injury and hemolysis of the erythrocytes at appropriate concentrations. Melittin is used as a marker for peptide-permeability of erythrocyte membranes. Melittin as an ion-membrane-altering agent is presented, where at certain concentrations, it can act as a tetramer and not just as a monomer when applied at low concentrations affecting the erythrocyte membrane [
3,
4,
5]. Melittin is used as an agent altering the permeable properties of the membrane for ions. Melittin also has hemolytic properties.
The biophysical properties of membranes largely determine the course of physiological and biochemical processes in the cell and are an important link in the morphological changes due to altered properties of erythrocytes in vitro.
Erythrocytes are used as an object to study the properties of biological membranes, as they are easily accessible and have a simple structure. They possess only one membrane and no cell organelles. In the present study, erythrocytes were used as a model system for studying the electrokinetic properties of membranes, and the mechanism of the non-specific binding of polyvalent cations (lectin: FITC-concanavalin A) and pooled immunoglobulin G to the membrane was investigated. A different function of erythrocytes is expected after a change in their morphology after exposure to biomacromolecules. It has been shown that red blood cells undergo morphological transitions from discocytes with thin, doughnut-like shapes to echinocytes and spherocytes using quantitative phase imaging techniques [
6]. Melittin induces morphological changes in red blood cells called echinocytosis [
6]. Electrical charges on the surface of the cell membrane are an important factor in determining the permeability of the walls of blood vessels, the transport of ions and macromolecules and interactions in intermolecular recognition processes. The surface electrical charge density of membranes and subcellular organelles is decisive in their interaction with the extracellular environment and adhesion and aggregation processes. Electrostatic forces can affect the localization and orientation of integral membrane proteins. Surface charge asymmetry, which arises from ion asymmetry in the bilayer, affects the correct orientation of the protein in the membrane, but transmembrane electric fields affect its conformation. Therefore, the modification of the surface electrical properties through the binding of ions or molecules to the surface of the studied objects in many cases leads to a number of changes in the molecular organization of the membrane and its biochemical and biophysical properties [
7].
Therefore, the study and analysis of the electrokinetic characteristics of membranes and the possibility of influencing them are important in optimizing the conditions of interactions of membranes with other molecules and aggregation in some cells.
The biophysical state of the erythrocyte membrane is directly related to the surface charge, the presence of which can be calculated based on the electrophoretic mobility and zeta potential of the cell [
8,
9]. The registration of the movement of blood cells in a constant electric field under physiological conditions of the suspension medium allows an evaluation of the functions of the erythrocyte membrane. A pronounced change in the electrophoretic mobility of erythrocytes is found in normal and pathological conditions [
10,
11]. The electrokinetic potential of cells is genetically determined, and its decrease is associated with the aggregation of erythrocyte membranes. A number of pathological processes are associated with a modification of the electrophoretic mobility and zeta potential of erythrocytes. Electrophoretic mobility is a measure of the magnitude of the surface electrical charge of cells without or in the presence of physical factors or chemical agents. General biophysical regularities at the membrane level may cause changes in the electrophoretic mobility, zeta potential and surface electrical charge of erythrocytes upon exposure to pooled IgG and hematin, which needs more studies. When various chemical agents are applied to the erythrocyte membranes, which are released during inflammation and/or in the presence of pooled immunoglobulin G, as well as the membrane-disruptive melittin in the presence of IgG, molecular changes to the plasma membrane of the cell are observed. Erythrocyte membranes are the immediate target of the damaging action of a number of factors in the pathological process of the erythrocyte life cycle together with the initiation of lipid peroxidation as a universal mechanism of cell damage [
9,
12]. The application of additional biophysical studies on erythrocyte membranes after exposure to pooled IgG, hematin and melittin could be a contribution to clarifying the mechanism of membrane alteration. The physical parameters of the interaction of melittin with model membranes need to be investigated via electrochemical impedance spectroscopy [
13].
Complex analysis of the electrokinetic properties of human erythrocytes in terms of dependence on Rhesus factor and other biophysical characteristics will allow us to obtain a general picture of the main mechanisms of action of hematin and pooled IgG on the functional activity of erythrocytes and the causes of the damaging effect of these biomacromolecules, of which actions on biological membranes have not been explained in detail. Rhesus (Rh) factor is a clinically significant blood group system in transfusion medicine [
14]. With different alterations to pooled IgG and hematin, as well as combinations of these agents, the action of melittin in the presence of pooled IgG on the erythrocyte membrane, evidenced by changes in the morphology of erythrocytes with biomacromolecules, hemolysis and acid-induced stability of erythrocytes, hematocrit and lipid peroxidation in the presence of these biological active agents is expected.
The disclosure of the biophysical mechanisms of changes in the erythrocyte membrane is extremely important in order to assess the state of interactions of hematin, pooled IgG and hematin, as well as pooled IgG and melittin, to evaluate the functional state of the cell and to develop methods for diagnosing the inflammatory process.
Based on the above, the aim of the present work was to study the electrokinetic properties of human erythrocytes and their interrelationship with the structural–functional state of the membrane in the norm and after influencing the functions with the biological active macromolecules pooled immunoglobulin G, hematin and melittin in vitro.
The present study will focus on the possibility of proving the use of electrokinetic parameters as a method for diagnosing inflammatory reactions in the body, related to the release of hematin and its interaction with pooled IgG in response to the applied impact (physical factors and chemical agents). The significance of biomacromolecule interactions with erythrocyte membranes provides a potential explanation of inflammatory diseases and anti-oxidative stress. The presented work would have future fundamental applications in biomedicine.
2. Materials and Methods
2.1. Materials
All chemicals used in the present study were of analytical grade. OCTAGAM® IVIG 5%, 2.5 gm in a 50 mL vial, (Human) Immune Globulin Intravenous—5% solution liquid preparation—was purchased from Octapharma International Services N.V., Octapharma N.V., De Frélaan 269/4, B-1180 Brussel. Human Immunoglobuline (50 g/L; min 95% IgG) 2.5 g/50 mL—Maltose 5 g—water injection contained the following subfractions: IgG1 62.6%, IgG2 30.1%, IgG3 6.1%, IgG4 1.2%. Melittin from bee venom, 65–85% (HPLC); FITC-CoA, concanavalin A FITC-labeled from Canavalia ensiformis (Jack bean) Type IV; Na2HPO4, sodium phosphate dibasic; KH2PO4, potassium phosphate monobasic were purchased from Sigma-Aldrich (St. Louis, MO, USA), and the chemicals were as follows: NaCl, KCl, NaOH; TCA (trichloroacetic acid); 2-thiobarbituric acid (TBA); NaN3, sodium azide; DMSO, dimethyl sulfoxide (methylsulfinylmethane); Hematin—Koch-Light Laboratories Ltd., Colnbrook, Bucks, UK. Propidium iodide was purchased from VECTOR Laboratories Inc. 6737 Mowry Ave Newark, CA 94560, USA. D (+)-Saccharose was purchased from Riedel-De Haën AG, Seelze, Hannover, Germany. Egg-yolk L-α-phosphatidylcholine (egg PC, 840051P) was from Avanti Polar Lipids Inc. Alabaster, AL, USA). Pentane CH3(CH2)3CH3 and hexane CH3(CH2)4CH3 of HPLC grade were supplied from Honeywell, Riedel-de Haën AG, Seelze, Hannover, Germany. Bidistilled water from a quartz distiller for the preparation of all aqueous solutions was used. All buffer media solutions were previously filtered through Whatman® membrane filters PTFE, cellulose nitrate, pore size 0.2 µm, diam. 47 mm, non-sterile, Whatman 7402-004, Whatman Article № 28420767, Sigma Aldrich, Merck KGaA, Darmstadt, Germany.
A hematin (ferriprotoporphyrine IX) stock solution was prepared in DMSO to a final concentration of 2.5 mM [
15]. Pooled immunoglobulin G preparations were diluted in PBS to 10 mg/mL [
16]. All treated preparations were stored at 4 °C in the presence of 0.1% NaN
3.
2.2. Isolation of Erythrocytes
Erythrocytes collected from complete blood samples taken from healthy adult subjects were supplied by the National Centre of Hematology and Perfusion, Sofia, Bulgaria. Venom blood samples were taken from healthy human subjects, using EDTA-anticoagulant vacutainers for the study. The present study complied with the ethical regulations and legislation in both Europe and Bulgaria. The experiments were performed in compliance with “WMA Declaration of Helsinki, Ethical Principles for Medical Research Involving Human Subjects” [
17]. All subjects recruited for the study provided written informed consent. Ethic Committee Name: Ethics Committee of Sofia University “St. Kliment Ohridski”, Approval Code:RD-04-91, date 25 February 2022.
Erythrocytes were centrifuged in a microcentrifuge Eppendorf
® MiniSpin
® with standard Rotor, Eppendorf AG, Hamburg, Germany at 2500 rpm for 5 min and were washed 3 times with phosphate-buffered saline (10.1 mM Na
2HPO
4, 1.8 mM KH
2PO
4, 136.9 mM NaCl, pH 7.4). Erythrocytes (hematocrit 0.20) were used within 4 h after preparation. We used phosphate-buffered saline (PBS, pH 7.4), containing 137 mM NaCl, 10.1 mM Na
2HPO
4 and 1.8 mM KH
2PO
4 for the determination of hematocrit, hemolysis (Hb-release) and HCl-hemolysis, membrane transport, electrokinetic parameters, lipid peroxidation and fluorescence microscopy measurements, respectively, where the stock suspension of erythrocytes was then diluted to 1% hematocrit. Erythrocytes were treated with biomacromolecules for 1 h at 37 °C, at a mixing speed of 300 rpm (TMix Thermalmixer, Analytic Jena AG, Jena, Germany). The effect of fixed concentrations of pooled immunoglobulin G (10 mg/mL), hematin (80 µM [
15]) and melittin (0.2 µg/mL Mt; 0.5 µg/mL Mt; 2 µg/mL Mt and 5 µg/mL Mt) treatments on biophysical parameters (average value ± SD) of erythrocytes with a Rhesus-positive factor and Rhesus-negative factor were investigated.
2.3. Hematocrit and Hemolysis Tests
We determined hematocrit via microcentrifugation (NF 048 NÜVE SANAYİ MALZEMELERİ İMALAT VE TİCARET A.Ş bench-top centrifuge, Akyurt—Ankara, Türkiye. Hematocrit adjustment was performed via blood centrifugation (12,500×
g for 2 min), plasma removal and addition of cells in the desired ratio to obtain 20% (
v/
v). The samples were aliquoted into Eppendorf tubes and homogenized. Tubes (Heparin Disposable Pipettes, containing 50 microliters, 67 mm length, made in USA, by Drumond Scientific Company, distributed by Rudolph GRAVE AB Stockholm, Sweden) of precise bore glass were filled with the human erythrocyte suspension, one end was sealed and the tubes were centrifuged for the constant packing of cells [
9]. The relative heights of the packed cell column and the total fluid were measured using a special optical device.
The hemolysis test at different concentrations of NaCl to determine the degree of hemolysis was used. The erythrocyte hemolysis test was fixed [
18] by adding erythrocytes to a series of hypotonic solutions with decreasing NaCl concentrations (0.9–0.3% NaCl) at 5% Hct, with incubation for 30 min at 25 °C with gentle mixing (speed of 300 rpm using TMix, Analytic Jena AG, Jena, Germany). Afterward, 1.5 mL of the samples with different salt concentrations were diluted with the samples of the erythrocyte suspensions previously incubated with biomacromolecus (1 h at 37 °C) at 25 °C for 30 min. The erythrocytes incubated with different hypotonic solutions were then centrifuged (12,000×
g for 1 min), and after that, the supernatants were removed. The hemoglobin of each supernatant was measured from the absorbance at λ = 576 nm immediately after centrifugation [
19] using a BOECO Spectrophotometer S-200 (VIS), Boeckel + Co (GmbH + Co) KG, Hamburg, Germany. The Abs (absorbance) was calculated by plotting the relationship between the absorbance at 576 nm, respectively, versus the absorbance at the appropriate concentration of the NaCl solution.
The value of
was calculated as follows:
where
is the absorbance Abs
(λ=576) of the release of hemoglobin from erythrocytes without or in the presence of different concentrations of biomacromolecules, measured at λ = 576 nm and
is the absorbance from the release of hemoglobin in erythrocytes without or in the presence of biomacromolecules, suspended into 0.9% NaCl solution, respectively.
2.4. Acid—Hemolytic Stability Assay
The acid-induced resistance of human erythrocytes has been studied in isosmotic media (8.5% NaCl) in the presence of 0.004 N HCl as a hemolytic agent. Erythrograms were obtained according to the classical method [
20]. The method of erythrograms is described in detail. In a blood sample obtained by diluting the blood with physiological solution about 10
3 fold, an acid hemolytic is placed, of which the chemical composition and concentration can be determined very precisely. Hydrochloric acid is used as this hemolytic. From the moment the hemolytic is introduced, the transparency of the sample is periodically measured, which gradually increases and after some time reaches a relatively high and constant value. The dependence of transparency on time is an integral curve that reflects the transition from the initial transparency of the sample to the final one. According to Terskov and Gitelzon [
20,
21], this integral curve is not sufficiently representative of the hemolytic process, and therefore, the authors replaced it with the first and time derivative, which expresses the dependence of the instantaneous rate of hemolysis on time. Since the measurements of the transparency of the sample during the hemolytic process are made only at individual moments, both the integral and the derivative curve have a polygonal character. The derived curve is called the acid erythrogram [
21]. The study of the hemolytic process is conveniently performed spectrophotometrically. The duration of the hemolytic process depends on the properties of the erythrocyte membranes, on the type and concentration of the hemolytic and on the temperature of the sample during hemolysis.
The difference between the absorbance of the erythrocyte membranes at the beginning of the process of hemolysis and the final value of absorbance represents the change in the absorbance coefficient, and we approximated its value as 100 per cent. The changes between two following absorbance values are represented as the percent of the whole change in absorbance. The upper values are proportional to the average velocity of the hemolytic process during every separate interval of 30 s between the 2 following measurements. The graph dependence of the changes in acid-resistance (partial hemolysis, hemolyzed erythrocyte fraction (%)) in time during 10 min represents the acid-induced erythrogram. Consequently, we used the acid erythrogram representing the 1st derivative of the optical density kinetic curve and characteristics of the erythrocyte population heterogeneity as in [
20,
22].
2.5. Microelectrophoresis
The electrophoretic mobility (EPM) measurements were performed using the microelectrophoresis technique with the OPTON Cytopherometer (Feintechnik Ges, m.b.H., Zeiss-Opton, Oberkochen, Germany). Electrophoretic mobility was measured in a rectangular chamber and platinum electrodes at a constant electric field of 5 mA and temperature of 25 ± 0.1 °C. The movement of erythrocytes over a known distance (16 μm) was timed for both forward and backward (reversed field) runs. The erythrocytes were observed under a light microscope connected to a Sony video camera (Video Camera Head CH–1400 CE, Sony Corporation, Japan) providing 2000 times magnification and a JVC monitor (Victor Company of Japan, LTD., Yokohama, Japan). The results were expressed by means of the EPM (per 10
−8 m
2 V
−1s
−1) ± standard deviations (SD) for each sample. The electrical conductivity and viscosity of the different erythrocyte suspensions were measured using a Thermo Fisher Scientific CyberScan PC 510 (Oakton
® Instruments/Eutech Instruments Pte Ltd., Singapore) pH/Conductivity meter and a Rheo (VEB MLW Prüfgeräte–WERK, MEDINGEN/SITZ FREITAL/GDR, Typ 202, Germany) viscometer, respectively. Values represent the mean of three replications (54–96 erythrocytes). The electrokinetic (zeta) potential (ζ) was calculated from the electrophoretic mobility,
u, using the Helmholtz–Smoluchowski equation [
23]:
where
is in units of mV,
is the relative dielectric permittivity of the aqueous phase,
is the permittivity of free space and
is the viscosity of the PBS: 136.9 mM NaCl, 10.1 mM Na
2HPO
4, 1.8 mM KH
2PO
4, pH 7.4 (pH 7.4 at 25 °C) as in [
24].
The electrostatic potential in the aqueous phase of the erythrocyte membrane surface and charge density (
σ) is given by the following:
where
is in mV,
at 25 °C,
,
23 mol
−1 is the Avogadro constant). The surface electrical charge of erythrocytes is expressed in C/m
2. The values of surface charge density were calculated according to the assumption that
[
24]. The electrophoretic mobility of the cells gives information about the dynamics of the surface electrical charge on the outer membrane surface, i.e., the average amount of electrical charges that is generated on the outer surface of the cell membrane of the erythrocyte was calculated. The surface electrical charge was calculated from the zeta potential of the cell by making the approximation that the electrokinetic potential is approximately equal to the surface or electrostatic potential of the cell according to the Gouy–Chapman theory. This explicitly states that the value of the zeta potential depends on the ionic strength. Therefore, erythrocytes were placed in a medium with an isotonic ionic strength to follow the change in the electrokinetic parameters of the erythrocyte membrane in the presence of pooled immunoglobulin G, hematin and a combination of pooled immunoglobulin G and melittin. The Gouy–Chapman theory was used in the calculation of the surface charge used in the determination of the surface electrical charge. The surface electrical charge of erythrocytes was determined to obtain a complete evaluation of the change in surface electrostatics without and in the presence of biomacromolecules in the suspending medium.
2.6. Measurements of Proton Transport
The maintenance of cellular homeostasis in the conditions of great differences in the chemical composition of the cytoplasm and the environment is ensured by the barrier functions of the membrane and the selective transport of substances in the cell. Erythrocytes can be considered systems in a non-equilibrium steady state. Under normal conditions, anion transport across the membrane is maintained in a certain equilibrium state. When the erythrocytes are transferred from saline to an isotonic sucrose solution of low ionic strength, a spike in the curve is observed at the beginning of the experiment, which is due to the escape of the intracellular solution. To some extent, of course, there is an antiport of chloride and bicarbonate ions dissolved in the medium (the suspension is in continuous contact with air). However, the time constants of this process are too large. In unbuffered solutions, the adaptive process is too fast and the new C-state is reached in less than 2 min. The equilibrium of the cells is mainly determined by the change in the external pH. This process can be explained, at least in part, by the minimal amount of OH
− and H
+ ions that must be transported from inside to outside the cell to establish equilibrium. The following processes are possible: (1) OH
−/Cl
− antiport or H
+/Cl
− cotransport; (2) Cl
− antiport with divalent ions. The transfer of erythrocytes into solutions with altered electrolyte content induces passive ion transport processes. According to Glaser and coworker’s model [
25] of ionic states in human erythrocytes, a quasi-equilibrium “C-state” exists, characterized by an equilibrium of all permeant anions and protons, but maintaining the initial Na
+ and K
+ contents. The kinetics of reaching this C-state is determined based on anion fluxes and components of the pH-equilibrium and is therefore limited by membrane permeability to them, as well as driving forces. In the presence of CO
2, HCO
3− and carboanhydrase, the equilibrium for Cl
− ions and pH is mediated by the highly efficient Jacobs–Stewart cycle. In most cases, the concentration of carboanhydrase, CO
2 and HCO
3− in the experimental solutions is much lower than that under physiological conditions. Therefore, under in vitro conditions, the unimpaired transport of Cl
− ions via the Jacobs–Stewart cycle cannot be expected, and the electroneutrality of the equilibrium process can only be maintained via the co-transport of Cl
− and H
+ or via their antiport against other anions OH
−.
The experimental study on the anion–proton co-transport was based on the measurement of net proton flows associated with erythrocyte Band 3-mediated net anion transfer. An erythrocyte suspension (500 µL, Hct = 20%) of previously incubated erythrocyte membranes (without or in the presence of biomacromolecules at 37 °C for an hour incubation and preincubation of pooled immunoglobulin G (10 mg/mL) and 80 µM hematin) was added to 50 mL of unbuffered isotonic sucrose medium, and the time course of the pH of the medium was registered for 10 min. The changes in pH are presented as a percent of the control. All experiments were carried out at 25 °C.
Erythrocytes suspended in hypotonic sucrose solution can be characterized by the exchange of inorganic anions of chloride and carbonate in connection with the pH equilibration, which occurs in a minutes. Water transport in an erythrocyte suspension equilibrates the osmotic gradient in less than one second [
25]. Erythrocytes are characterized by a “C-state” with a stable quasi-stationary state for hours. We measured the extracellular proton concentration (H
+ext) as a function of time in seconds promoted by the treatment of erythrocytes with biomacromolecules as in [
26]. The suspending medium of 0.3 M sucrose (NaOH), pH 7.4, to maintain its buffer capacity constant over the pH range covered in the erythrocyte experiments was used [
25].
Proton efflux began by mixing 500 µL of the erythrocyte suspended into 50 mL of 0.3 M sucrose (NaOH), pH 7.4. The pH of the erythrocyte suspension without or in the presence of different concentrations of biomacromolecules at the same time was measured using a Thermo Fisher Scientific CyberScan PC 510 (Oakton® Instruments/Eutech Instruments Pte Ltd., Singapore) pH/conductivity meter.
The results of the proton efflux alterations in extracellular media in the presence of different concentrations of biomacromolecules were obtained from membrane transport measurements in erythrocyte suspending media every 20 s during 10 min with gentle mixing. The value of
(%) was calculated from the following:
where
and
are the pH values of erythrocytes in biomacromolecule-free medium and in the presence of biomacromolecule concentrations, respectively.
2.7. Lipid Peroxidation Determined with Thiobarbituric Acid Reactive Substances (TBARS)
Malondialdehyde content was measured according to [
27,
28] with modifications. The erythrocyte suspensions (500 μL erythrocytes in PBS, pH 7.4, 2 mM NaN
3, Hct = 20%) without or in the presence of biomacromolecules after an incubation and preincubation time period were homogenized in 400 μL of 28% trichloroacetic acid (TCA) and centrifuged at 12,500 ×
g for 4 min. After centrifugation, 1 mL of supernatant was mixed with 500 μL of 1% thiobarbituric acid (TBA) in 1% NaOH, and the mixture was incubated in boiling water for 30 min. The suspending medium was centrifuged at 12,500×
g for 2 min, and the absorbance of λ = 532 nm was measured using a BOECO Spectrophotometer S-200 (VIS), Boeckel + Co (GmbH + Co) KG, Hamburg, (Germany) to determine the MDA content. The thiobarbituric acid-reactive substance (TBARS) molar concentration,
c, was calculated as follows:
where
is the absorbance,
stands for the molar absorption coefficient of H
2O
2,
and
represents the optical path length. Afterwards, the reaction was stopped by cooling the samples in an ice bath. MDA reacts in the TBA test to generate a colored product. In acid solution, the product absorbs light at 532 nm. MDA content was estimated by using an extinction coefficient of 154 mmol L
−1 cm
−1. The lipid peroxidation of erythrocyte membranes was determined based on the production of TBARS and expressed in μmol L
−1 [
27].
2.8. Fluorescence Microscopy Studies
The samples were observed using a Zeiss Axioscope 5 microscope with a fluorescence LED Illumination Colibri 3 (Carl Zeiss Microscopy GmbH, Jena, Germany) at 1000× magnification (objectives 100× and eyepieces 10×) under immersion, with appropriate excitation filters for propidium iodide (PI) staining solution and fluorescein isothiocyanate (FITC): blue, 478–495 nm. The results were documented with an Axiocam 202 mono digital camera (Carl Zeiss Microscopy GmbH, Jena, Germany) and ZEN 2.5 (blue edition) software (Carl Zeiss Microscopy GmbH, Jena, Germany). Fluorescein isothiocyanate (FITC)-labeled lectin concanavalin A (CoA) was used. Concanavalin A-FITC labeled from Canavalia ensiformis (Jack bean) Type IV, FITC content of 3.6 mol/mol lectin (Mol.Wt of lectin approx. 102,000) has an affinity for terminal α-D-mannosyl and α-D-glycosyl residues. ConA-FITC inhibitory carbohydrates are α-methylmannoside and α-methylglucoside. FITC-Con A (MW 102,000 Da) labeling solutions were dissolved in distilled water at 2 mg/mL as a stock solution in dark tubes. They were diluted with PBS, pH 7.4, to 40 µg/mL before use. Erythrocyte membranes were transferred into the PI staining solution. After 60 min of incubation at 37 °C, erythrocytes (2 × 106 cells/mL) were washed with PBS (15 min 12,000× g, Micro22R, Andreas Hettich GmbH & Co. KG, Tuttlingen, Germany)) and examined under via epifluorescence using a Zeiss Axioscope 5 (Carl Zeiss Microscopy GmbH, Jena, Germany).
Fluorescence microscopy of erythrocytes with FITC-labeled CoA samples followed the protocol: microscopic preparations made of all samples in duplicate, 20 μL of each sample, in separate Eppendorf tubes, were placed in the resuspended sediment with erythrocytes in 100 μL PBS, pH 7.4. Then, 15 μL of propidium iodide solution (PI, 1.0 mg/mL solution in water, Invitrogene, Thermo Fisher Scientific Inc., Eugene, OR, USA, cat. P3566) was added to each. Samples were incubated for 30 min at 37 °C and then centrifuged at 14,000 rpm for 5 min. The pellet of each sample was resuspended in 35 μL of PBS buffer, and the entire amount was spotted onto a glass slide coated with poly-L-lysine (poly-L-lysine is coated glass slides, Poly-Prep Slides, Sigma-Aldrich, Merck, Saint Louis, MO, USA, cat. P0425-72EA) and covered with cover glass. The preparations were placed in a humid chamber (a box at the bottom of which there were several layers of filter paper wetted with PBS), wrapped with aluminum foil. The preparations were incubated at +4 °C, overnight. For better stabilization of the fluorescent signal in erythrocytes without and in the presence of biomacromolecules, they were allowed to stain better for the formation of a luminescent halo around the erythrocyte, which does not form immediately after incubation with FITC-concanavalin A. The cells were faintly stained, and their luminescence was not immediately noticeable after treating them with FITC-labeled lectin on the same day after incubation following the protocol for labeling them. Letting the treated samples stand with FITC-concanavalin A was necessary for better fluorescence microscopy images and better light intensity, where no change in the morphology of the cells was observed after they were stained in the solution overnight. The results were documented with an Axiocam 202 mono digital camera (Carl Zeiss Microscopy GmbH, Jena, Germany) and ZEN 2.5 (blue edition) software (Carl Zeiss Microscopy GmbH, Jena, Germany) [
29].
2.9. Electrochemical Impedance Spectroscopy of Bilayer Lipid Membranes
We studied the effect of melittin on the electrical properties of bilayer lipid membranes (BLM) formed via the Montal–Mueller method [
30,
31] from egg-yolk L-α-phosphatidylcholine. Pentane CH
3(CH
2)
3CH
3 and hexane CH
3(CH
2)
4CH
3 of HPLC grade were used for dissolving the lipid. The Montal–Mueller chamber, model BC-20A, was provided by Eastern Scientific LLC (Rockville, MD, USA). Solvent-free bilayers were obtained suspended on the aperture with a diameter of 100 µm in a 0.025 mm thin Teflon membrane according to the corresponding protocol [
30] as described in detail in [
31]. The final control of the bilayer formation and its quality were verified electrically [
32]. This was proceeded to the fast Fourier transform electrochemical impedance spectroscopy (FFT-EIS) measurement immediately afterwards. A multisine perturbation signal was applied, characterized by a small amplitude ~10 mV in the frequency range of 1.5 Hz–50 kHz. The measurement procedure comprised the simultaneous acquisition of the perturbation and the respective response signals followed by their fast Fourier transformation (FFT) to the frequency domain. This approach allowed the impedance spectrum to be acquired in a couple of seconds and the stationarity to be simultaneously monitored [
33,
34], which is particularly appropriate for probing the impedance characteristics of lipid membranes.
The capacitance
and resistance
of the planar lipid membrane were deduced from the analysis of the acquired FFT-EIS data taking into account the equivalent circuit representing the Montal–Mueller BLM configuration as discussed in [
31]. The membrane specific capacitance
and resistance
were calculated from the measured capacitance and resistance of the bilayer and its 100 µm-circle surface area
8 × 10
−5 cm
2. The values reported below were calculated as the weighted average of at least 3 independent measurements averaged over 10 repetitions each.
2.10. Statistical Analysis
The electrokinetic data were averaged from triplicate measurements for every sample. The data are expressed as the mean ± SD. The significant means were determined by use of ANOVA. One-way analysis of variance was performed with Dunn’s Test following ranked-based ANOVA (Kruskal–Wallis One Way Analysis of Variance on Ranks) and the Student–Newman–Keuls method taking p < 0.05 as significant, p < 0.01 as highly significant and p < 0.001 as extremely significant and represented by an asterisk in the figures. Statistical analyses were also performed using Minitab v.17.
4. Discussion
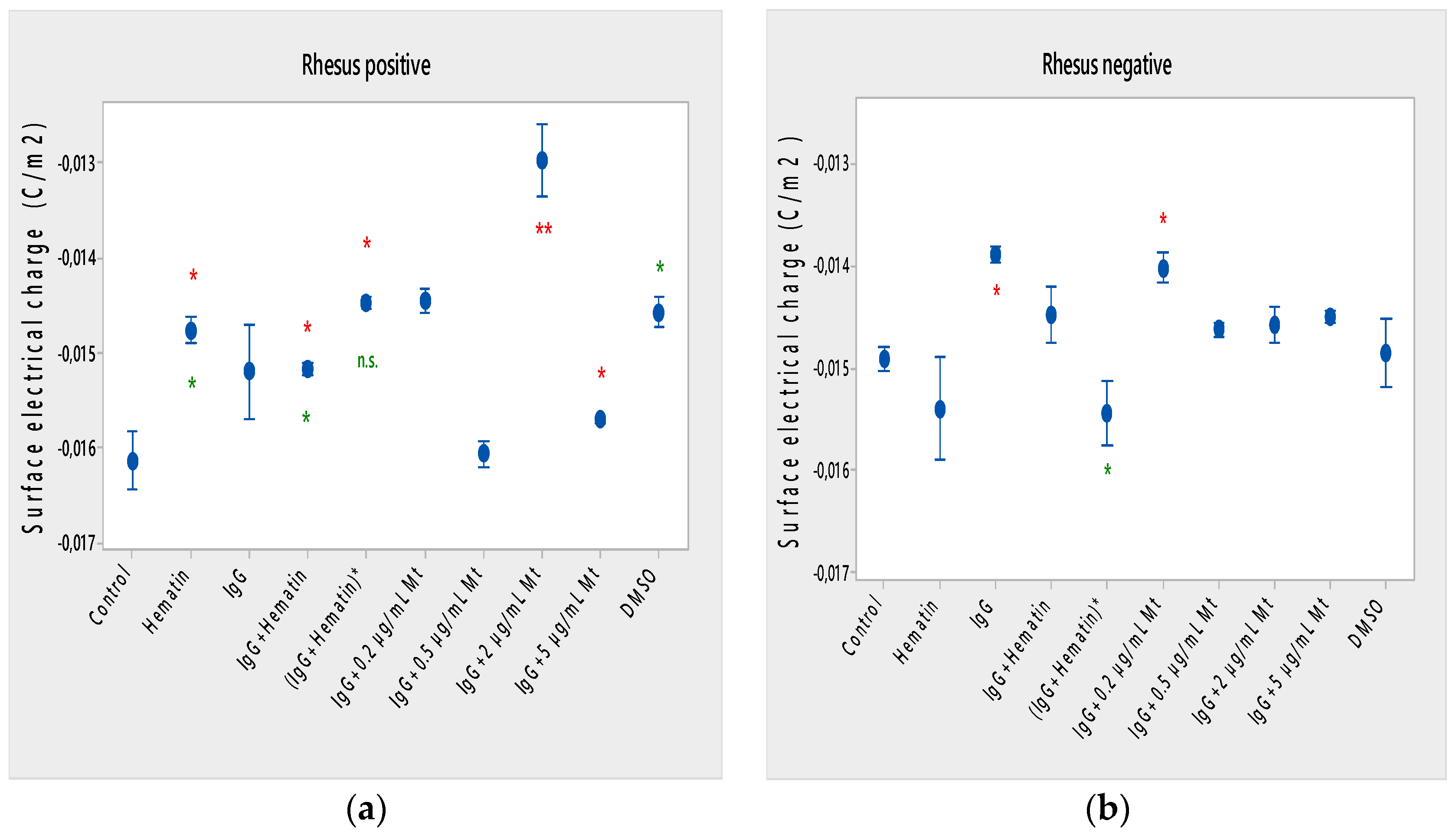
The open question of how the electrokinetic and other biophysical properties of the membranes behave under pooled IgG, hematin and melittin treatments in terms of dependence on the Rhesus factor of erythrocytes remains unclear, taking into account the effective charges of the biomacromolecules that they exhibit during their interaction with the cell. When the erythrocytes come into contact with the antibodies, they are fixed on the surface of the erythrocyte membranes. In this way, the surfaces of erythrocytes do not carry their own charge, but the charge of globulin molecules, which is much smaller. Therefore, the erythrocyte suspension when treated with pooled IgG is expected to be much more unstable, and aggregation occurs more easily. The results reported here indicate that pooled IgG causes a decrease in the surface electrical charge of erythrocytes with a Rhesus-negative factor, but does not affect erythrocytes with a Rhesus-positive factor. Pooled IgG can be considered a bipolar ion with charge homeostasis in physiological media (PBS, pH 7.4) with an effective Debye–Hückel–Henry charge between −3 and −9 [
42], the effective charge being calculated as Z
DHH = 7.7 ± 0.2 [
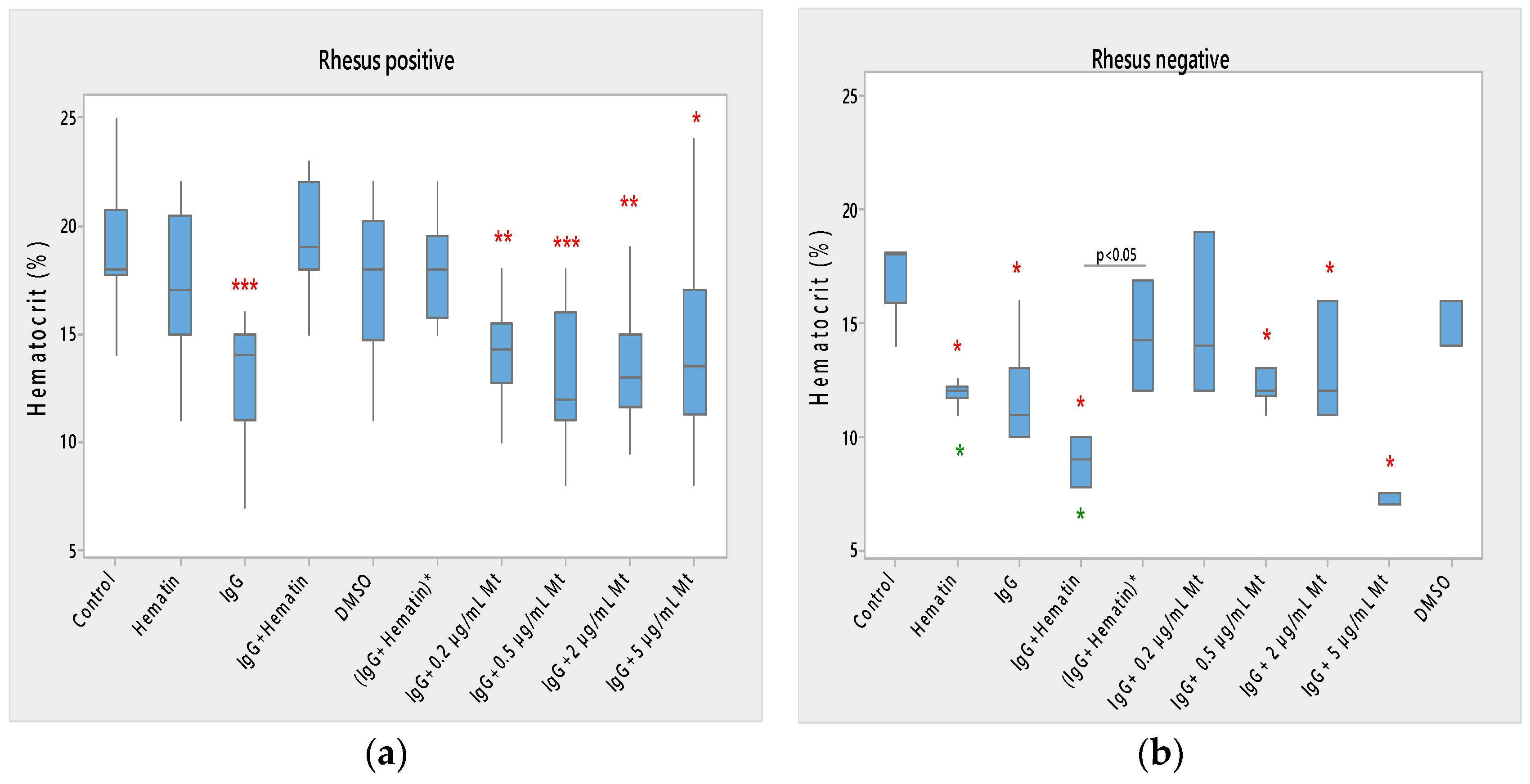
43], with the indication that salt and temperature dependence can be included in the calculation of the effective charge of pooled immunoglobulins. Changes in cell morphology in Rh-positive cells include the appearance of echinocytes, codocytes, stomatocytes and “ghost” cells. Pooled IgG (10 mg/mL) leads to a decrease in hematocrit and acid resistance of erythrocytes with a Rhesus-positive factor, but causes an increase in membrane transport, as well as the process of a strong release of hemoglobin in a medium of 0.3% NaCl. The action of pooled IgG on erythrocytes with a Rhesus-negative factor differs from that on erythrocytes with a Rhesus-positive factor, except in the case of lipid peroxidation of cells, where no change in secondary products of lipid peroxidation is registered. Pooled IgG causes the appearance mainly of codocytes, as well as of echinocytes, when examining the morphology of erythrocytes. In erythrocytes with a Rhesus-negative factor, pooled IgG causes a decrease in electrophoretic mobility, electrokinetic potential and surface electrical charge, hematocrit and acid-hemolytic stability of cells. Pooled IgG leads to an increase in membrane transport across erythrocyte membranes with a Rhesus-negative factor, as well as in the release of hemoglobin in a medium of 0.3% NaCl.
Heme acts as a signaling molecule, which suggests the possibility of dynamic and rapid mobilization from different carrier and scavenger proteins [
44,
45]. The heme-induced activation of the blood coagulation system is suggested as a mechanism for the initiation of thrombotic events under hemolytic conditions [
46]. Several reports describe the determination of heme concentrations in different conditions, which range from ~20 to >350 µM as occurring, e.g., in heme-driven pathologies [
47]. In our studies, the concentration of 80 µM hematin was used, at which there is weak release of hemoglobin from the cells, but the acid-hemolytic stability is significantly reduced. We believe that the presence of hematin (80 µM) and pooled IgG (10 mg/mL) in the erythrocyte suspension creates a situation where the antibody should appear as a cation, because of the acidification of the environment due to the hematin pKa (strongest acidic), which is 3.68, and pKa (strongest basic), which is 4.96. The physiological charge of hematin is (−2) [
48]. Therefore, the action of the preincubated sample of pooled IgG with hematin and of the pooled IgG in the presence of hematin will have a strong effect in decreasing the surface charge of Rh-positive erythrocytes, but will not affect the surface electrical charge of erythrocytes with the Rh-negative factor. Obviously, the preincubation of pooled IgG with hematin at 37 °C, as well as the action of pooled IgG and hematin on erythrocytes with a Rhesus-negative factor, could eliminate changes in the surface electrical charge of the cell in our experimental setup. That is the reason the changes caused by hematin need to be investigated.
In the present study, using erythrocytes with Rhesus-positive and Rhesus-negative factors, the action of pooled IgG (10 mg/mL), hematin (80 µM) and melittin (0.2 and 0.5 µg/mL; 2 and 5 µg/mL) on erythrocyte membranes was studied in detail.
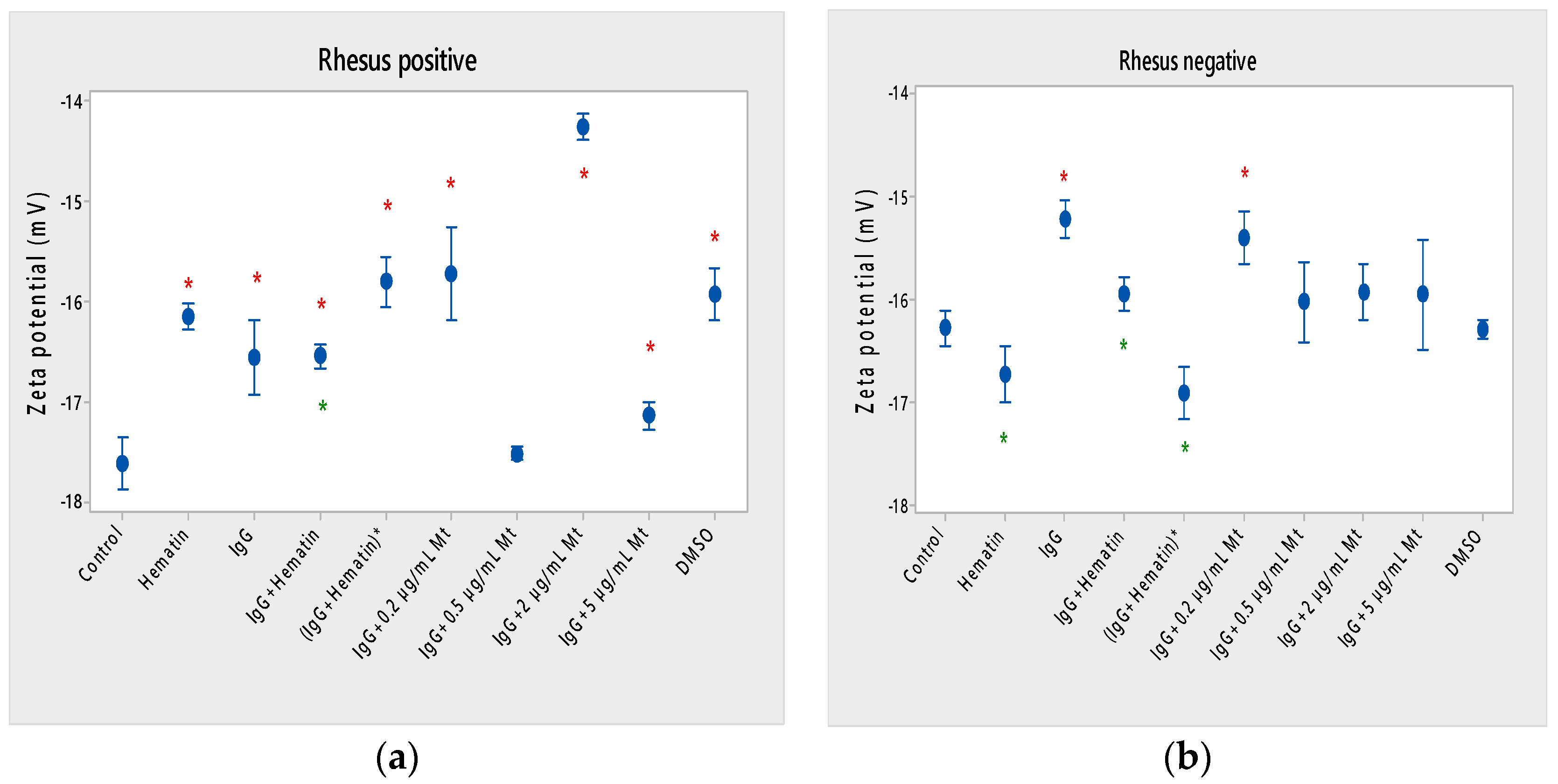
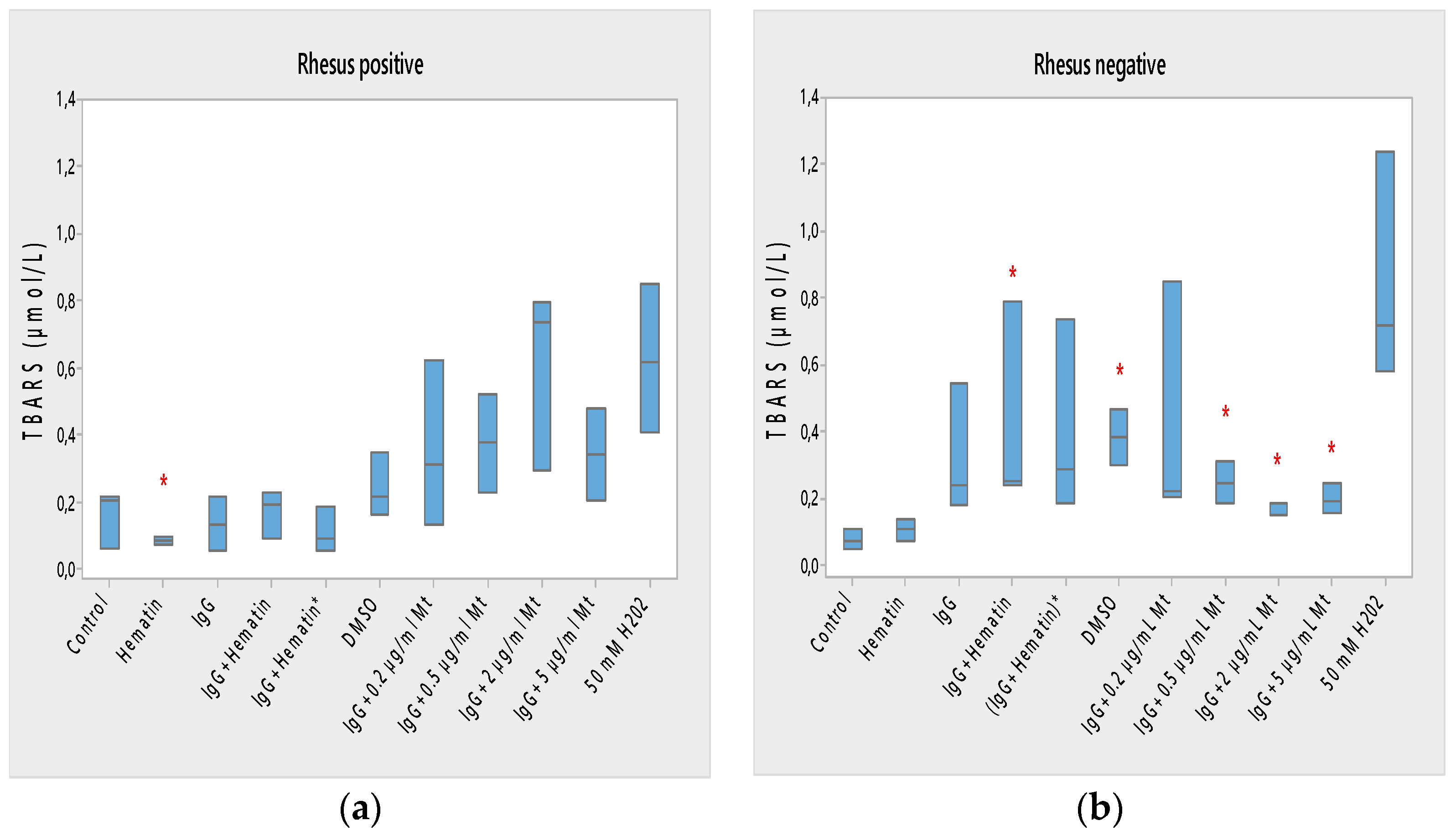
Hematin (80 µM) decreases the lipid peroxidation in the erythrocyte membrane, the electrokinetic properties of erythrocytes with a Rh-positive factor and the acid-hemolytic stability of the cell. Therefore, hematin eliminates the increased amount of radical forms from erythrocytes after treatment with DMSO, in which hematin is dissolved in our studies. Hematin preserves the tendency to decrease the electrophoretic mobility, zeta potential and surface electrical charge of Rh-positive erythrocytes after treatment with DMSO. The direction of action of hematin or DMSO in the treatment of erythrocyte membranes with a Rhesus-positive factor is also similar. In this case, the hematocrit does not change, the release of hemoglobin from erythrocytes is slightly increased, the membrane transport through the membrane is strongly increased and the acid-hemolytic stability of the cells is significantly reduced. Erythrocytes with a Rhesus-positive factor treated with hematin are characterized by the appearance of “ghost” cells, echinocytes and codocytes. In the case of erythrocytes (Rhesus-negative factor), codocytes and spherocytes are observed after treatment with 80 μM hematin. This is dominated by “ghost” cells with a loss of hemoglobin. The ghost cells indicate coagulative necrosis where there is cell death but retainment of cellular architecture. Spherocytes are characteristic of hemolytic anemia. A reduction in free radical forms (TBARS) of erythrocytes with a Rhesus-positive factor after exposure to hematin is observed due to a decreased concentration of malondialdehyde in cells.
The combined effect of pooled IgG and hematin, as well as the preincubation of the individual components, shows its different effect on cell morphology and biophysical parameters of erythrocytes with Rhesus-positive and those with Rhesus-negative factors. More codocytes, echinocytes and pyropoikilocytes are observed in the erythrocyte sample with a Rhesus-positive factor. Most of the codocytes are resistant to osmotic lysis compared to those not treated with pooled IgG and hematin control cells. Confirmation for this is obtained from the results of the release of hemoglobin on erythrocytes with a Rhesus-positive factor, as Hb-release is strongly reduced compared to control values, untreated cells. This effect is related to the increase in the ratio between the surface area and the volume of the cells and the decreased osmotic resistance of the erythrocytes with a Rhesus-positive factor. Treatment with pooled IgG and hematin causes the appearance of stomatocytes, except for codocytes in erythrocytes with the Rh-negative factor. Stomatocytosis are due to alterations in permeability, leading to an increased volume. “IgG+hematin” and preincubated (IgG+Hematin)* results in changes in lipid peroxidation, hematocrit and hemoglobin release from erythrocytes with a Rhesus-negative factor. The sample containing “IgG+hematin” significantly increased TBARS products from lipid peroxidation, in contrast to the absence of such in the pre-incubated form (IgG+Hematin)* on erythrocytes with a Rhesus-negative factor. “IgG and hematin” preparations lead to a strong decrease in the hematocrit of erythrocytes with a Rhesus-negative factor, in contrast to the effect of preincubated (IgG+Hematin)*, where no changes in the volume percentage of the red blood cells in the blood with a Rhesus-negative factor are observed. Rhesus-negative factor erythrocytes treated with “IgG+hematin” cause an increase in Hb release from cells, and those treated with pre-incubated (IgG+hematin)* are not characterized by changes in hemoglobin release from Rhesus-negative factor erythrocytes. It is important to note that erythrocytes with a Rhesus-positive factor are characterized by a lack of changes in lipid peroxidation, hematocrit and the release of hemoglobin from the cell, as well as an increase in membrane transport after incubation with “IgG+hematin” and pre-incubated (IgG+hematin)*. A significant decrease in the electrokinetic parameters of the erythrocytes with a Rhesus-positive factor and of the acid resistance is observed after the treatment of cells with “IgG+hematin” and of the pre-incubated sample of (IgG+hematin)*. The results show that compared to that in the control containing DMSO in the erythrocyte suspension, there is an increase in the zeta potential of Rh-negative erythrocytes in the presence of hematin and no statistically significant changes in the electrokinetic potential of Rh-positive erythrocytes. Treatment with preincubated (IgG and hematin)* in erythrocytes with Rh-positive factor causes the appearance of codocytes, stomatocytes and echinocytes, unlike the presence of many “ghost” cells, codocytes and spherocytes in cells with a Rhesus-negative factor.
Hematin and pooled IgG, as well as the pre-incubated preparation of (IgG+hematin)* mark a slight increase in the negative zeta potential values (* p < 0.05) of Rh-negative erythrocytes, which we believe is due to the ability of hematin to influence the conformational changes of the molecule of the pooled IgG and convert it into an IgG/hematin complex capable of causing the exposure of additional negatively charged groups on the surface of Rh-negative erythrocytes. The zeta potential changes are significant, and hematin causes an increase in the negative electrokinetic potential of the membrane when only the hematin molecule is applied in comparison to that of the DMSO-control. It can be seen that the zeta potential of Rh-negative erythrocytes treated with pooled IgG and hematin is slightly decreased (* p < 0.05) compared to that of the DMSO-treated control, but the pre-incubated sample of both components show an increase in electrokinetic potential of the erythrocyte membrane with a Rhesus-negative factor.
The kinetics of membrane transport of erythrocytes with the Rh-positive or Rh-negative factor are characterized by an increase in ΔpH changes up to 20 s and the subsequent influx of protons into the interior of erythrocytes from 20–600 s of measurement.
The main action of melittin on erythrocyte membranes is associated with its hemolytic activity [
49,
50,
51,
52]. In the present work, the action of melittin on human erythrocytes as a melittin-induced membrane permeability agent with a monomer species [
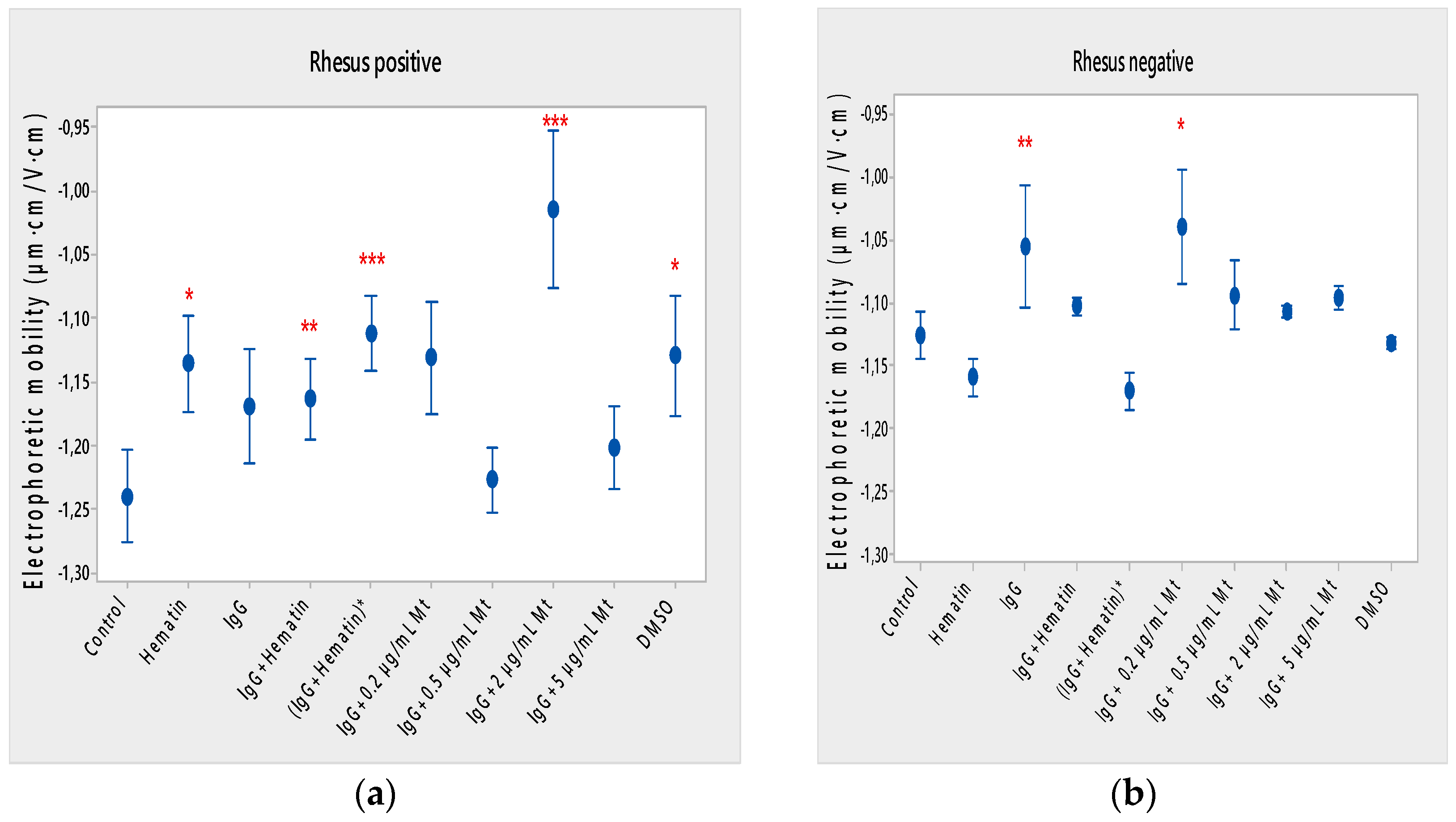
53] is investigated, where the hemolytic action of low concentrations of melittin on cells (Rhesus-positive factor, previous investigations) is established. The emphasis in the present research involving disruptive monomeric melittin-membrane properties has been shifted to the influence of pooled IgG in the presence of fixed concentrations of melittin, where a decrease in the zeta potential of cells under the action of different concentrations of melittin is found.
The effective charge of melittin in isotonic PBS medium, pH 7.4, is calculated as 4.24 [
54]. The combined action of melittin and pooled IgG makes the electrokinetic behavior of erythrocytes different in the presence of fixed concentrations of melittin, which is known to increase the electrokinetic potential of human erythrocytes, without the presence of pooled IgG in the erythrocyte suspension [
55]. Pooled IgG (10 mg/mL) and melittin caused an increase in lipid peroxidation of Rh-negative erythrocytes except for the sample containing (IgG + 0.2 µg/mL) melittin. The action of pooled IgG and fixed concentrations of melittin does not affect MDA-products in Rh-positive erythrocytes. The fixed concentration of 0.2 µg/mL melittin in the presence of 10 mg/mL pooled IgG leads to a decrease in electrophoretic mobility and a decrease in surface electrical charge due to a decrease in the zeta potential of erythrocytes with a Rhesus-positive factor, via a decrease in the acid-hemolytic stability of cells, an increase in membrane transport of the erythrocyte suspension. Data from fluorescent microscopy confirm the presence of echinocytes, spherocytes and ghost cells in erythrocytes with Rhesus-positive factor, as well as echinocytes in the effect on erythrocytes with a Rhesus-negative factor in the presence of pooled IgG and 0.2 μg/mL melittin. These forms of erythrocytes are characteristic of other concentrations of the effect of pooled IgG and melittin on erythrocytes with the Rh-positive factor. Pooled IgG in the presence of 2 µg/mL melittin induces an increased amount of radical forms of Rh-negative erythrocytes and leads to a decrease in hematocrit and acid resistance, but is associated with an increase in membrane transport across the membrane. However, with the doses of pooled IgG and 2 μg/mL melittin, ovalocytes are observed in cases of exposure to erythrocytes with the Rh-negative factor. The effects of “IgG + 2 µg/mL melittin” or “IgG + 5 µg/mL melittin” preparations on the biophysical parameters of the membrane decrease the surface charge of erythrocytes with a Rhesus-positive factor, but significantly reduce the hematocrit, acid-hemolytic stability of erythrocytes with both Rh factors. Rhesus-positive and Rhesus-negative erythrocytes, after treatment with “IgG + 2 µg/mL melittin” and “IgG + 5 µg/mL melittin”, are characterized by a strong increase in the membrane transport of protons, chloride and hydroxyl ions, which means that pooled IgG in the presence of 2 µg/mL melittin and 5 µg/mL melittin, respectively, increases H
+/Cl
− coexchange and H
+/OH
− transport, reducing the acid resistance of erythrocyte membranes and hematocrit. The additional TBARS products are not detected as a result of incubation with Rh-positive erythrocytes, but lipid peroxidation increases significantly in Rh-negative erythrocytes. When pooled IgG and 0.5 μg/mL melittin act on erythrocytes with a Rhesus-positive factor, stomatocytes are observed, as well as with a dose of impact of pooled IgG and 5 μg/mL melittin on erythrocytes with a Rhesus-negative factor. The different forms of erythrocytes after their impact with biomacromolecules will also lead to different biophysical actions on the membranes of cells with the Rh-positive or Rh-negative factor.
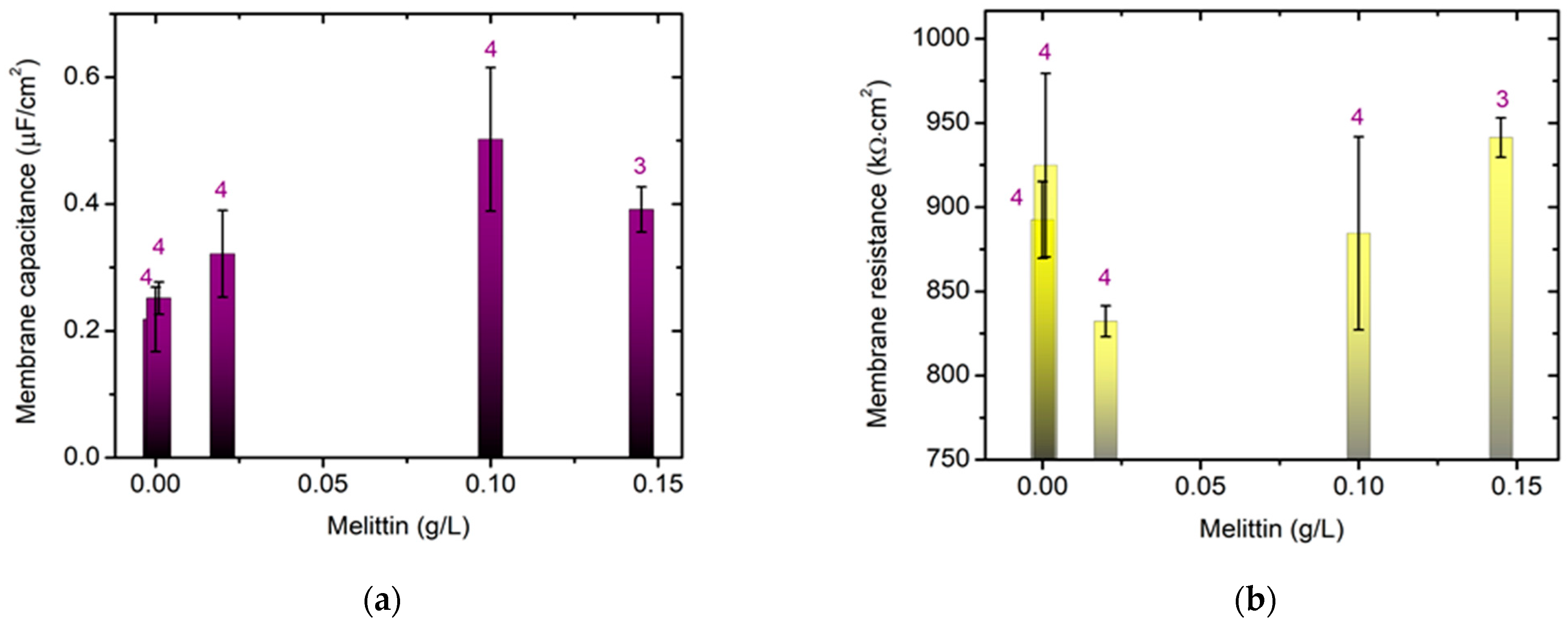
Based on our previous studies on the effect of melittin on the electrokinetic properties of biological membranes, we consider it particularly important to monitor the membrane capacitance of model lipid bilayer membranes in the presence of different concentrations of melittin. The electrochemical impedance study of model lipid bilayers in the presence of melittin yields the membrane electrical capacitance and resistance in low ionic strength media (I = 0.003 mol/L). As expected, at low concentrations, ~0.02 g/L monomeric melittin triggers pore formation [
56] resulting in lower specific resistance of the bilayers, as shown in
Figure 6b. Benachir and Lafleur [
57] investigated the mechanism of interactions of melittin with lipid vesicles and found that melittin leads to a complete and abrupt emptying of the contents of the vesicles. This mechanism obeys the “all or nothing” principle, where melittin acts rapidly within seconds to minutes [
57].
In the presence of higher peptide concentrations (~0.15 g/L) in the buffer medium [
4,
5,
58], melittin tetramers form, by means of hydrophobic bonds between the monomers [
4,
5]. The aggregation of melittin is expected at the higher melittin concentrations, ~0.15 g/L, in the bulk phase [
59] at which increased resistance is measured compared to that in untreated lipid bilayers (
Figure 6b). Lipid bilayer capacitance is higher in the presence of 0.10 g/L melittin compared to that in the control, which is likely related to bilayer thinning [
60] and alterations to the dielectric permittivity of melittin-treated membranes. Thus, the stabilization of the lipid phosphatidylcholine bilayer is characterized also by higher specific capacitance and resistance upon treatment with tetrameric melittin in the presence of 0.15 g/L melittin in the buffer medium and a higher membrane capacity compared to that of the control. The positively charged peptide interacts electrostatically with the membrane. An increase in the surface density of the negative charge of the membrane leads to a decrease in the lytic power of the membrane. The effective charge of melittin is about 2 [
61] and is significantly lower than its net electrical charge of 5–6 expected at neutral pH.
The interaction of melittin with the membrane is sensitive to the lipid composition of the bilayer. Lipids with negatively charged heads and cholesterol inhibit the lytic activity of melittin [
62,
63]. It has been suggested that the primary and secondary structure of a peptide determines its interaction with specific lipid moieties. Even when there is no lytic activity, melittin binds to the membrane. Therefore, the reduced activity of the peptide is not caused by reduced binding to the lipid bilayer.
As the concentration of melittin increases, the membrane capacitance increases, and at 0.15 g/L melittin in the egg–yolk lecithin system, a decrease in the electrical capacitance value is observed compared to that with 0.10 g/L melittin, which could be explained by the thinning of the lipid bilayer thickness observed with the higher concentrations of the cationic peptide [
60] and a subsequent decrease in the effect of melittin on membrane capacitance.
The obtained results might be useful in understanding the functional significance of therapeutic antibodies exposed to pro-oxidants, using the biophysical explanation of their action on human erythrocytes [
64]. The acquired knowledge can also be used as additional information at the membrane level in unraveling the role of therapeutic immunoglobulin G in its application in inflammatory diseases [
65].
5. Conclusions
The pooled IgG, hematin, melittin and combinations of these chemical agents change the morphology of erythrocytes from both the Rhesus-positive or Rhesus-negative factors. This is due to effects on the activity of the Band 3 protein, which carries out proton and anion transport through the erythrocyte membrane depending on the Rhesus factor of the erythrocytes. It is necessary to note the important role of the spectrin network in the shear elasticity of the red blood cells, which also exhibits a change in the bending rigidity [
65].
The electrokinetic properties of erythrocytes are affected, which are characterized by a reduction in negatively charged surface-exposed groups in the presence of pooled IgG and hematin, as well as its preincubated form, after exposure to erythrocytes with a Rhesus-positive factor. The oxidative status of erythrocytes with a Rhesus-negative factor is also affected, which demonstrates an increase in lipid peroxidation under the influence of pooled IgG and hematin, as well as under the influence of pooled IgG with fixed concentrations of melittin. It is noted that MDA products are affected only under the influence of 80 µM hematin after exposure to erythrocytes with the Rh-positive factor. It is necessary to note the influence of the studied biomacromolecules on the hematocrit of erythrocytes. The hematocrit of erythrocytes with a Rhesus-positive factor is affected only by pooled IgG and its combination with melittin, in contrast to the hematocrit of erythrocytes with a Rhesus-negative factor, where a reduction in the parameter is also observed with hematin, pooled IgG and hematin and pooled IgG and melittin. Thus, we prove that the noticed biomacromolecules in the applied concentrations of impact on the erythrocyte membrane lead to a change in the biophysical parameters of the erythrocytes depending on their Rhesus factor (Rhesus-positive or Rhesus-negative).
Pooled IgG and hematin, as well as the preincubated sample, strongly suppress the acid resistance of young erythrocytes with the Rh-positive factor, but reduce it to a lesser extent in old erythrocytes with an Rh-negative factor. When increasing melittin concentrations to 0.15 g/L, an increase in membrane capacitance of egg yolk PC lipid bilayer is observed, but with a decrease in the membrane resistance, which is associated with the tetrameric form of the peptide affecting the impedance of the model system.
The present studies add to our fundamental knowledge of the action of pooled immunoglobulin G and hematin, as well as pooled immunoglobulin G and melittin, on the human erythrocyte membrane.



 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





