
Optimization of Membrane Protein TmrA Purification Procedure Guided by Analytical Ultracentrifugation


Abstract
:1. Introduction
2. Materials and Methods
2.1. Expression and Purification of TmrA
2.1.1. Expression of TmrA in E. coli
2.1.2. Purification of TmrA in DDM
2.1.3. Detergent Exchange
2.2. Analytical Ultracentrifugation
2.3. Negative Staining Electron Microscope
3. Results
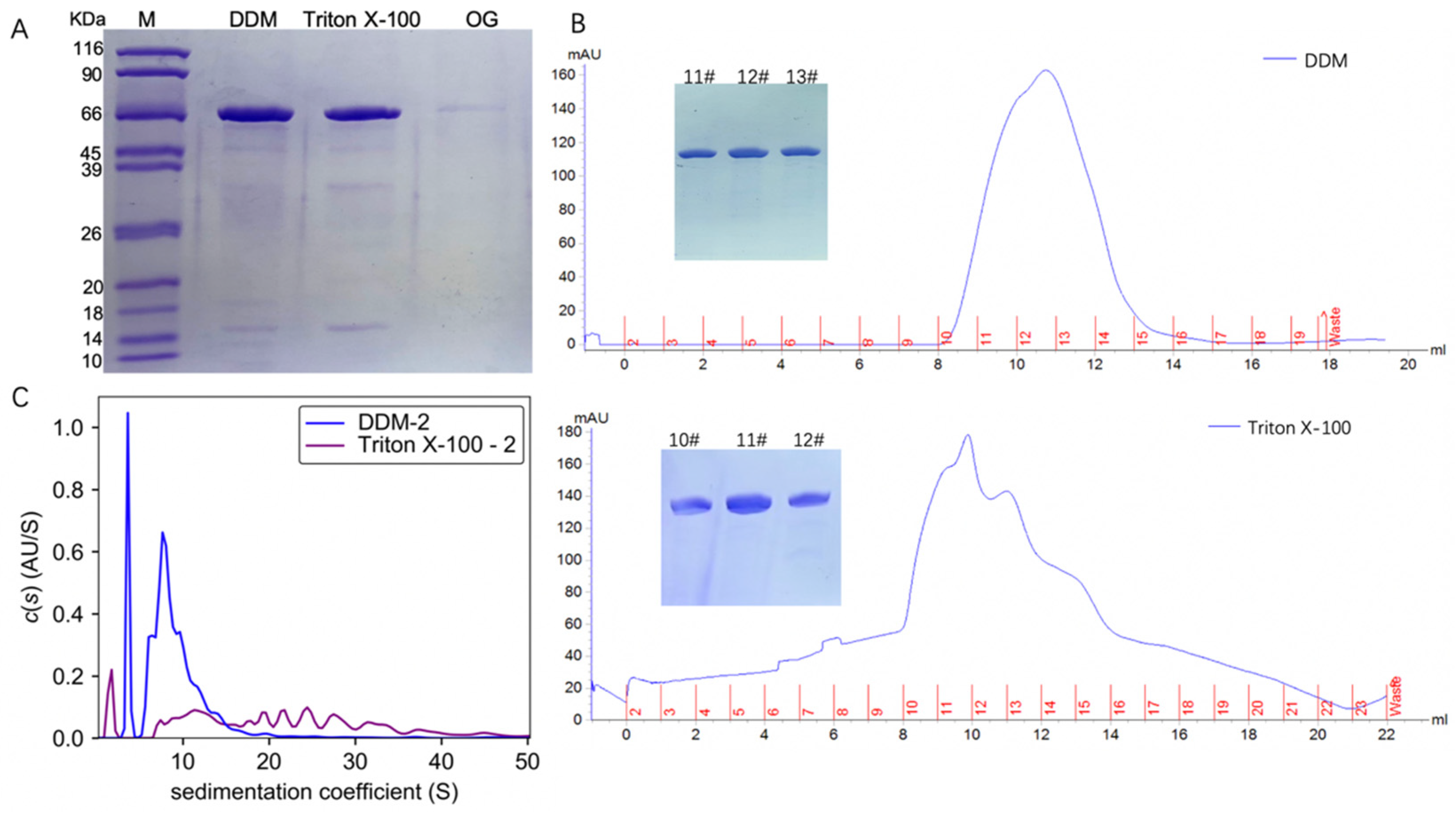
3.1. TmrA Possesses Different Aggregation Status in Distinct Detergents
3.2. High Concentration of DDM Improves the Homogeneity of TmrA
3.3. Detergent Supplementation Is a Cost-Effective Strategy and Does Not Compromise Protein Homogeneity or Aggregation State
3.4. Negative Stain Electron Microscope Defines Protein Aggregation States of TmrA Purified by the Optimized Workflow
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Jawhari, A. Editorial—Membrane protein tools for drug discovery. Methods 2020, 180, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Von Heijne, G. The membrane protein universe: What’s out there and why bother? J. Intern. Med. 2007, 261, 543–557. [Google Scholar] [CrossRef] [PubMed]
- Errey, J.C.; Fiez-Vandal, C. Production of membrane proteins in industry: The example of GPCRs. Protein Expr. Purif. 2020, 169, 105569. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.; Shin, K.; Patterson, R.E.; Liu, X.-Q.; Rainey, J.K. Current strategies for protein production and purification enabling membrane protein structural biology. Biochem. Cell Biol. 2016, 94, 507–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, S.-H.; Guidotti, G. Chapter 35 Purification of Membrane Proteins. Methods Enzymol. 2009, 463, 619–629. [Google Scholar] [CrossRef] [PubMed]
- Stock, C.; Hänelt, I. Membrane Protein Solubilization and Quality Control: An Example of a Primary Active Transporter. Methods Mol. Biol. 2020, 2127, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Ebel, C. Sedimentation velocity to characterize surfactants and solubilized membrane proteins. Methods 2011, 54, 56–66. [Google Scholar] [CrossRef]
- Kim, J.; Wu, S.; Tomasiak, T.M.; Mergel, C.; Winter, M.B.; Stiller, S.B.; Robles-Colmanares, Y.; Stroud, R.M.; Tampé, R.; Craik, C.S.; et al. Subnanometre-resolution electron cryomicroscopy structure of a heterodimeric ABC exporter. Nat. Cell Biol. 2015, 517, 396–400. [Google Scholar] [CrossRef] [Green Version]
- Ravaud, S.; Cao, M.-A.D.; Jidenko, M.; Ebel, C.; Le Maire, M.; Jault, J.-M.; Di Pietro, A.; Haser, R.; Aghajari, N. The ABC transporter BmrA from Bacillus subtilis is a functional dimer when in a detergent-solubilized state. Biochem. J. 2006, 395, 345–353. [Google Scholar] [CrossRef] [Green Version]
- Doshi, R.; Ali, A.; Shi, W.; Freeman, E.V.; Fagg, L.A.; van Veen, H.W. Molecular Disruption of the Power Stroke in the ATP-binding Cassette Transport Protein MsbA. J. Biol. Chem. 2013, 288, 6801–6813. [Google Scholar] [CrossRef] [Green Version]
- Barth, K.; Hank, S.; Spindler, P.E.; Prisner, T.F.; Tampé, R.; Joseph, B. Conformational Coupling and trans-Inhibition in the Human Antigen Transporter Ortholog TmrAB Resolved with Dipolar EPR Spectroscopy. J. Am. Chem. Soc. 2018, 140, 4527–4533. [Google Scholar] [CrossRef]
- Nöll, A.; Thomas, C.; Herbring, V.; Zollmann, T.; Barth, K.; Mehdipour, A.R.; Tomasiak, T.M.; Brüchert, S.; Joseph, B.; Abele, R.; et al. Crystal structure and mechanistic basis of a functional homolog of the antigen transporter TAP. Proc. Natl. Acad. Sci. USA 2017, 114, E438–E447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, H.; Lakatos-Karoly, A.; Vogel, R.; Nöll, A.; Tampé, R.; Glaubitz, C. Coupled ATPase-adenylate kinase activity in ABC transporters. Nat. Commun. 2016, 7, 13864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zutz, A.; Hoffmann, J.; Hellmich, U.; Glaubitz, C.; Ludwig, B.; Brutschy, B.; Tampé, R. Asymmetric ATP Hydrolysis Cycle of the Heterodimeric Multidrug ABC Transport Complex TmrAB from Thermus thermophilus. J. Biol. Chem. 2011, 286, 7104–7115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millan, C.R.; Francis, M.; Khandelwal, N.K.; Thompson, V.F.; Thaker, T.M.; Tomasiak, T.M. A Conserved Motif in Intracellular Loop 1 Stabilizes the Outward-Facing Conformation of TmrAB. J. Mol. Biol. 2021, 433, 166834. [Google Scholar] [CrossRef] [PubMed]
- Velamakanni, S.; Yao, Y.; Gutmann, D.A.P.; Van Veen, H.W. Multidrug Transport by the ABC Transporter Sav1866 fromStaphylococcus aureus†. Biochemistry 2008, 47, 9300–9308. [Google Scholar] [CrossRef] [PubMed]
- Stefan, E.; Hofmann, S.; Tampé, R. A single power stroke by ATP binding drives substrate translocation in a heterodimeric ABC transporter. eLife 2020, 9, 9. [Google Scholar] [CrossRef] [PubMed]
- Infed, N.; Hanekop, N.; Driessen, A.J.; Smits, S.H.; Schmitt, L. Influence of detergents on the activity of the ABC transporter LmrA. Biochim. et Biophys. Acta (BBA)-Biomembr. 2011, 1808, 2313–2321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bechara, C.; Nöll, A.; Morgner, N.; Degiacomi, M.T.; Tampé, R.; Robinson, C. A subset of annular lipids is linked to the flippase activity of an ABC transporter. Nat. Chem. 2015, 7, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Bartoschik, T.; Gupta, A.; Kern, B.; Hitchcock, A.; Adams, N.B.; Tschammer, N. Quantifying the Interaction of Phosphite with ABC Transporters: MicroScale Thermophoresis and a Novel His-Tag Labeling Approach. Biophys. Membr. Proteins: Methods Protoc. 2020, 2168, 51–62. [Google Scholar]
- Barth, K.; Rudolph, M.; Diederichs, T.; Prisner, T.F.; Tampé, R.; Joseph, B. Thermodynamic Basis for Conformational Coupling in an ATP-Binding Cassette Exporter. J. Phys. Chem. Lett. 2020, 11, 7946–7953. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Andya, J.D.; Shire, S.J. A critical review of analytical ultracentrifugation and field flow fractionation methods for measuring protein aggregation. AAPS J. 2006, 8, E580–E589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arthur, K.K.; Kendrick, B.S.; Gabrielson, J.P. Guidance to Achieve Accurate Aggregate Quantitation in Biopharmaceuticals by SV-AUC. Methods Enzymol. 2015, 562, 477–500. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Yadav, S.; Andya, J.; Demeule, B.; Shire, S.J. Analytical Ultracentrifugation and Its Role in Development and Research of Therapeutical Proteins. Methods Enzymol. 2015, 562, 441–476. [Google Scholar] [CrossRef]
- Salvay, A.G.; Santamaria, M.; le Maire, M.; Ebel, C. Analytical Ultracentrifugation Sedimentation Velocity for the Characterization of Detergent-Solubilized Membrane Proteins Ca++-ATPase and ExbB. J. Biol. Phys. 2007, 33, 399–419. [Google Scholar] [CrossRef] [Green Version]
- Guillet, P.; Mahler, F.; Garnier, K.; Boussambe, G.N.M.; Igonet, S.; Vargas, C.; Ebel, C.; Soulié, M.; Keller, S.; Jawhari, A.; et al. Hydrogenated Diglucose Detergents for Membrane-Protein Extraction and Stabilization. Langmuir 2019, 35, 4287–4295. [Google Scholar] [CrossRef] [PubMed]
- Wen-Dan, C.; Yang, X.; Cui-Yan, Z.; Ya-Fei, L.; Xiao-Xia, Y.; Rui-Xuan, Z. Wen-Qi, L. Study of membrane protein TmrAB by Analytical Ultracentrifugation. Prog. Biochem. Biophys. 2018, 45, 1047–1053. [Google Scholar]
- Zhou, R.; Yang, G.; Shi, Y. Dominant negative effect of the loss-of-function gamma-secretase mutants on the wild-type enzyme through heterooligomerization. Proc. Natl. Acad. Sci. USA 2017, 114, 12731–12736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rames, M.; Yu, Y.; Ren, G. Optimized Negative Staining: A High-throughput Protocol for Examining Small and Asymmetric Protein Structure by Electron Microscopy. J. Vis. Exp. 2014, 2014, e51087. [Google Scholar] [CrossRef] [PubMed]
- Seddon, A.; Curnow, P.; Booth, P.J. Membrane proteins, lipids and detergents: Not just a soap opera. Biochim. et Biophys. Acta (BBA)-Biomembr. 2004, 1666, 105–117. [Google Scholar] [CrossRef] [Green Version]
- Lund, S.; Orlowski, S.; De Foresta, B.; Champeil, P.; Le Maire, M.; Møller, J.V. Detergent structure and associated lipid as determinants in the stabilization of solubilized Ca2+-ATPase from sarcoplasmic reticulum. J. Biol. Chem. 1989, 264, 4907–4915. [Google Scholar] [CrossRef]
- Le Roy, A.; Wang, K.; Schaack, B.; Schuck, P.; Breyton, C.; Ebel, C. AUC and Small-Angle Scattering for Membrane Proteins. Methods in Enzymology 2015, 562, 257–286. [Google Scholar] [CrossRef] [PubMed]
- Fleming, K.G. Determination of Membrane Protein Molecular Weight Using Sedimentation Equilibrium Analytical Ultracentrifugation. Curr. Protoc. Protein Sci. 2008, 53, 7–12. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak Data | Units | DDM-2 | DDM-6 | DDM-10 |
|---|---|---|---|---|
| MW 1 | KDa | 64.6 | 64.6 | 64.6 |
| Mp 2 | KDa | 52.3 | 66.6 | 64.2 |
| f/f0 | 1.26 | 1.37 | 1.335 | |
| delta D 3 | 157 ± 7 | 135 ± 6 | 117 ± 5 | |
| Abs/IF 4 | ||||
| s20,w | S | 6.4/6.1 | 6.2/5.9 | 6.0/5.7 |
| proportion of total | % | 22.7/18.4 | 61.3/26.8 | 56.7/10.8 |
| Stokes radius | nm | 3.39/4.41 | 3.50/5.21 | 3.48/4.75 |
| a/b (oblate) | 1.47/5.03 | 2.12/8.00 | 2.23/6.72 | |
| a/b (prolate) | 1.45/4.69 | 2.07/7.23 | 2.18/6.15 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, D.; Chu, W.; Sheng, X.; Li, W. Optimization of Membrane Protein TmrA Purification Procedure Guided by Analytical Ultracentrifugation. Membranes 2021, 11, 780. https://doi.org/10.3390/membranes11100780
Li D, Chu W, Sheng X, Li W. Optimization of Membrane Protein TmrA Purification Procedure Guided by Analytical Ultracentrifugation. Membranes. 2021; 11(10):780. https://doi.org/10.3390/membranes11100780
Chicago/Turabian StyleLi, Dongdong, Wendan Chu, Xinlei Sheng, and Wenqi Li. 2021. "Optimization of Membrane Protein TmrA Purification Procedure Guided by Analytical Ultracentrifugation" Membranes 11, no. 10: 780. https://doi.org/10.3390/membranes11100780
APA StyleLi, D., Chu, W., Sheng, X., & Li, W. (2021). Optimization of Membrane Protein TmrA Purification Procedure Guided by Analytical Ultracentrifugation. Membranes, 11(10), 780. https://doi.org/10.3390/membranes11100780