
Vaccine Design and Vaccination Strategies against Rickettsiae

Abstract
1. Introduction
2. Adaptive Immunity Is Essential for Defense against Rickettsial Infections
3. Immunopathology in Rickettsial Infections
4. Vaccination against Rickettsiae with Whole-Cell Antigen (WCA)
5. Immunogenic Determinants and Vaccine Candidates
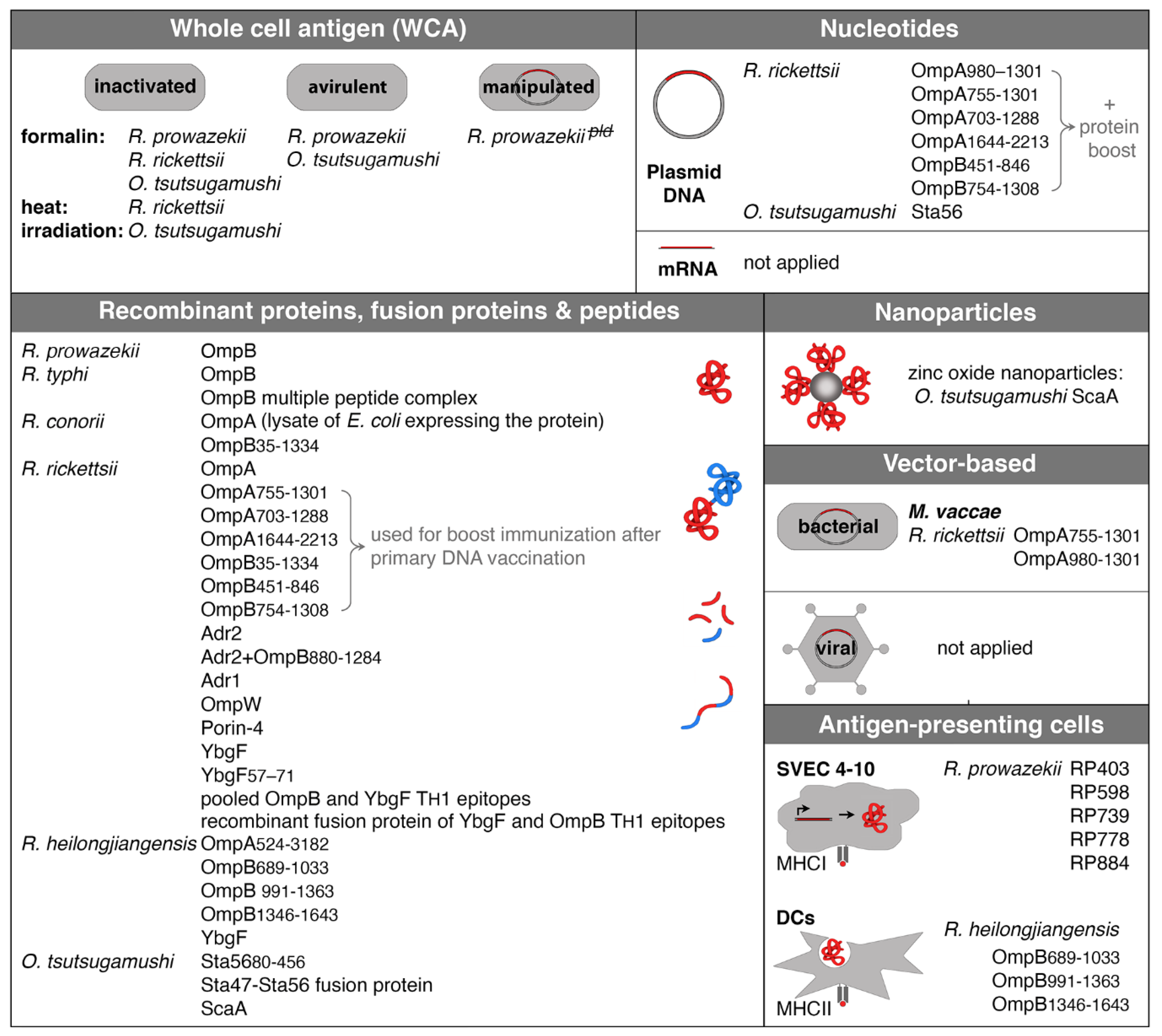
6. Experimental Approaches of Vaccination against Rickettsiae
6.1. Immunization with Recombinant Proteins and Peptides
6.2. Immunization with Antigen-Coupled Nanoparticles
6.3. Immunization with Nucleotides
6.4. Vector-Based Immunization: Adenoviral Vectors
6.5. Vaccination with Genetically Modified Bacterial Vectors
6.6. Immunization with Antigen-Expressing Cells or Antigen-Pulsed APCs
7. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Izzard, L.; Fuller, A.; Blacksell, S.D.; Paris, D.H.; Richards, A.L.; Aukkanit, N.; Nguyen, C.; Jiang, J.; Fenwick, S.; Day, N.P.; et al. Isolation of a novel Orientia species (O. chuto sp. nov.) from a patient infected in Dubai. J. Clin. Microbiol. 2010, 48, 4404–4409. [Google Scholar] [CrossRef]
- Abarca, K.; Martinez-Valdebenito, C.; Angulo, J.; Jiang, J.; Farris, C.M.; Richards, A.L.; Acosta-Jamett, G.; Weitzel, T. Molecular Description of a Novel Orientia Species Causing Scrub Typhus in Chile. Emerg. Infect. Dis. 2020, 26, 2148–2156. [Google Scholar] [CrossRef] [PubMed]
- Abdad, M.Y.; Abou Abdallah, R.; Fournier, P.E.; Stenos, J.; Vasoo, S. A Concise Review of the Epidemiology and Diagnostics of Rickettsioses: Rickettsia and Orientia spp. J. Clin. Microbiol. 2018, 56, e01728-17. [Google Scholar] [CrossRef] [PubMed]
- Mansueto, P.; Vitale, G.; Cascio, A.; Seidita, A.; Pepe, I.; Carroccio, A.; di Rosa, S.; Rini, G.B.; Cillari, E.; Walker, D.H. New insight into immunity and immunopathology of Rickettsial diseases. Clin. Dev. Immunol. 2012, 2012, 967852. [Google Scholar] [CrossRef] [PubMed]
- Sahni, S.K.; Rydkina, E. Host-cell interactions with pathogenic Rickettsia species. Future Microbiol. 2009, 4, 323–339. [Google Scholar] [CrossRef] [PubMed]
- Silverman, D.J.; Bond, S.B. Infection of human vascular endothelial cells by Rickettsia rickettsii. J. Infect. Dis. 1984, 149, 201–206. [Google Scholar] [CrossRef]
- Hackstadt, T. The biology of rickettsiae. Infect. Agents Dis. 1996, 5, 127–143. [Google Scholar]
- Kim, M.J.; Kim, M.K.; Kang, J.S. Involvement of lipid rafts in the budding-like exit of Orientia tsutsugamushi. Microb. Pathog. 2013, 63, 37–43. [Google Scholar] [CrossRef]
- Radulovic, S.; Price, P.W.; Beier, M.S.; Gaywee, J.; Macaluso, J.A.; Azad, A. Rickettsia-macrophage interactions: Host cell responses to Rickettsia akari and Rickettsia typhi. Infect. Immun. 2002, 70, 2576–2582. [Google Scholar] [CrossRef]
- Curto, P.; Riley, S.P.; Simoes, I.; Martinez, J.J. Macrophages Infected by a Pathogen and a Non-pathogen Spotted Fever Group Rickettsia Reveal Differential Reprogramming Signatures Early in Infection. Front. Cell. Infect. Microbiol. 2019, 9, 97. [Google Scholar] [CrossRef]
- Drevets, D.A.; Leenen, P.J.; Greenfield, R.A. Invasion of the central nervous system by intracellular bacteria. Clin. Microbiol. Rev. 2004, 17, 323–347. [Google Scholar] [CrossRef]
- Osterloh, A.; Papp, S.; Moderzynski, K.; Kuehl, S.; Richardt, U.; Fleischer, B. Persisting Rickettsia typhi Causes Fatal Central Nervous System Inflammation. Infect. Immun. 2016, 84, 1615–1632. [Google Scholar] [CrossRef] [PubMed]
- Regan, J.J.; Traeger, M.S.; Humpherys, D.; Mahoney, D.L.; Martinez, M.; Emerson, G.L.; Tack, D.M.; Geissler, A.; Yasmin, S.; Lawson, R.; et al. Risk factors for fatal outcome from rocky mountain spotted Fever in a highly endemic area-Arizona, 2002–2011. Clin. Infect. Dis. 2015, 60, 1659–1666. [Google Scholar] [CrossRef]
- Kuloglu, F. Rickettsial infections. Dis. Mol. Med. 2013, 1, 39–45. [Google Scholar] [CrossRef]
- Raoult, D.; Woodward, T.; Dumler, J.S. The history of epidemic typhus. Infect. Dis. Clin. N. Am. 2004, 18, 127–140. [Google Scholar] [CrossRef]
- Li, Z.; Xin, H.; Sun, J.; Lai, S.; Zeng, L.; Zheng, C.; Ray, S.E.; Weaver, N.D.; Wang, L.; Yu, J.; et al. Epidemiologic Changes of Scrub Typhus in China, 1952–2016. Emerg. Infect. Dis. 2020, 26, 1091–1101. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Murine typhus—Hawaii, 2002. MMWR Morb. Mortal. Wkly. Rep. 2003, 52, 1224–1226. [Google Scholar]
- Centers for Disease Control and Prevention. Outbreak of Rickettsia typhi infection—Austin, Texas, 2008. MMWR Morb. Mortal. Wkly. Rep. 2009, 58, 1267–1270. [Google Scholar]
- Adjemian, J.; Parks, S.; McElroy, K.; Campbell, J.; Eremeeva, M.E.; Nicholson, W.L.; McQuiston, J.; Taylor, J. Murine typhus in Austin, Texas, USA, 2008. Emerg. Infect. Dis. 2010, 16, 412–417. [Google Scholar] [CrossRef]
- Parola, P.; Miller, R.S.; McDaniel, P.; Telford, S.R., 3rd; Rolain, J.M.; Wongsrichanalai, C.; Raoult, D. Emerging rickettsioses of the Thai-Myanmar border. Emerg. Infect. Dis. 2003, 9, 592–595. [Google Scholar] [CrossRef]
- Suputtamongkol, Y.; Suttinont, C.; Niwatayakul, K.; Hoontrakul, S.; Limpaiboon, R.; Chierakul, W.; Losuwanaluk, K.; Saisongkork, W. Epidemiology and clinical aspects of rickettsioses in Thailand. Ann. N. Y. Acad. Sci. 2009, 1166, 172–179. [Google Scholar] [CrossRef]
- Phongmany, S.; Rolain, J.M.; Phetsouvanh, R.; Blacksell, S.D.; Soukkhaseum, V.; Rasachack, B.; Phiasakha, K.; Soukkhaseum, S.; Frichithavong, K.; Chu, V.; et al. Rickettsial infections and fever, Vientiane, Laos. Emerg. Infect. Dis. 2006, 12, 256–262. [Google Scholar] [CrossRef]
- Maude, R.R.; Maude, R.J.; Ghose, A.; Amin, M.R.; Islam, M.B.; Ali, M.; Bari, M.S.; Majumder, M.I.; Tanganuchitcharnchai, A.; Dondorp, A.M.; et al. Serosurveillance of Orientia tsutsugamushi and Rickettsia typhi in Bangladesh. Am. J. Trop. Med. Hyg. 2014, 91, 580–583. [Google Scholar] [CrossRef]
- Dupont, H.T.; Brouqui, P.; Faugere, B.; Raoult, D. Prevalence of antibodies to Coxiella burnetti, Rickettsia conorii, and Rickettsia typhi in seven African countries. Clin. Infect. Dis. 1995, 21, 1126–1133. [Google Scholar] [CrossRef]
- Acuna-Soto, R.; Calderon-Romero, L.; Romero-Lopez, D.; Bravo-Lindoro, A. Murine typhus in Mexico City. Trans. R. Soc. Trop. Med. Hyg. 2000, 94, 45. [Google Scholar] [CrossRef]
- Dittrich, S.; Rattanavong, S.; Lee, S.J.; Panyanivong, P.; Craig, S.B.; Tulsiani, S.M.; Blacksell, S.D.; Dance, D.A.; Dubot-Peres, A.; Sengduangphachanh, A.; et al. Orientia, rickettsia, and leptospira pathogens as causes of CNS infections in Laos: A prospective study. Lancet. Glob. Health 2015, 3, e104–e112. [Google Scholar] [CrossRef]
- Koliou, M.; Psaroulaki, A.; Georgiou, C.; Ioannou, I.; Tselentis, Y.; Gikas, A. Murine typhus in Cyprus: 21 paediatric cases. Eur. J. Clin. Microbiol. Infect. Dis. 2007, 26, 491–493. [Google Scholar] [CrossRef] [PubMed]
- Psaroulaki, A.; Christou, C.; Chochlakis, D.; Tsiligianni, I.; Sandalakis, V.; Georgalis, L.; Ioannou, I.; Giorgalas, G.; Tselentis, Y. Murine typhus in Cyprus: A 9-year survey. Trans. R. Soc. Trop. Med. Hyg. 2012, 106, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Chaliotis, G.; Kritsotakis, E.I.; Psaroulaki, A.; Tselentis, Y.; Gikas, A. Murine typhus in central Greece: Epidemiological, clinical, laboratory, and therapeutic-response features of 90 cases. Int. J. Infect. Dis. 2012, 16, e591–e596. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lledo, L.; Gegundez, M.I.; Medina, J.; Gonzalez, J.V.; Alamo, R.; Saz, J.V. Epidemiological study of Rickettsia typhi infection in two provinces of the north of Spain: Analysis of sera from the general population and sheep. Vector Borne Zoonotic Dis. 2005, 5, 157–161. [Google Scholar] [CrossRef]
- Bernabeu-Wittel, M.; del Toro, M.D.; Nogueras, M.M.; Muniain, M.A.; Cardenosa, N.; Marquez, F.J.; Segura, F.; Pachon, J. Seroepidemiological study of Rickettsia felis, Rickettsia typhi, and Rickettsia conorii infection among the population of southern Spain. Eur. J. Clin. Microbiol. Infect. Dis. 2006, 25, 375–381. [Google Scholar] [CrossRef]
- Nogueras, M.M.; Cardenosa, N.; Sanfeliu, I.; Munoz, T.; Font, B.; Segura, F. Serological evidence of infection with Rickettsia typhi and Rickettsia felis among the human population of Catalonia, in the northeast of Spain. Am. J. Trop. Med. Hyg. 2006, 74, 123–126. [Google Scholar] [CrossRef]
- Espinosa, N.; Canas, E.; Bernabeu-Wittel, M.; Martin, A.; Viciana, P.; Pachon, J. The changing etiology of fever of intermediate duration. Enferm. Infecc. Y Microbiol. Clin. 2010, 28, 416–420. [Google Scholar] [CrossRef]
- Andre, E.; Correia, R.; Castro, P.; Neto, M.; Rola, J.; Bacelar, F.; Oliveira, I.; Velosa, I.; Feio, A.; Filipe, A. Murine typhus in Portugal. Acta Med. Port. 1998, 11, 81–85. [Google Scholar] [PubMed]
- Bacellar, F.; Lencastre, I.; Filipe, A.R. Is murine typhus re-emerging in Portugal? Eurosurveillance 1998, 3, 18–20. [Google Scholar] [CrossRef]
- Badiaga, S.; Benkouiten, S.; Hajji, H.; Raoult, D.; Brouqui, P. Murine typhus in the homeless. Comp. Immunol. Microbiol. Infect. Dis. 2012, 35, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Dhimal, M.; Dumre, S.P.; Sharma, G.N.; Khanal, P.; Ranabhat, K.; Shah, L.P.; Lal, B.K.; Jha, R.; Upadhyaya, B.P.; Acharya, B.; et al. An outbreak investigation of scrub typhus in Nepal: Confirmation of local transmission. BMC Infect. Dis. 2021, 21, 193. [Google Scholar] [CrossRef] [PubMed]
- Bise, G.; Coninx, R. Epidemic typhus in a prison in Burundi. Trans. R. Soc. Trop. Med. Hyg. 1997, 91, 133–134. [Google Scholar] [CrossRef]
- Raoult, D.; Roux, V.; Ndihokubwayo, J.B.; Bise, G.; Baudon, D.; Marte, G.; Birtles, R. Jail fever (epidemic typhus) outbreak in Burundi. Emerg. Infect. Dis. 1997, 3, 357–360. [Google Scholar] [CrossRef]
- Raoult, D.; Ndihokubwayo, J.B.; Tissot-Dupont, H.; Roux, V.; Faugere, B.; Abegbinni, R.; Birtles, R.J. Outbreak of epidemic typhus associated with trench fever in Burundi. Lancet 1998, 352, 353–358. [Google Scholar] [CrossRef]
- Tarasevich, I.; Rydkina, E.; Raoult, D. Outbreak of epidemic typhus in Russia. Lancet 1998, 352, 1151. [Google Scholar] [CrossRef]
- Lutwick, L.I. Brill-Zinsser disease. Lancet 2001, 357, 1198–1200. [Google Scholar] [CrossRef]
- Parker, R.T.; Menon, P.G.; Merideth, A.M.; Snyder, M.J.; Woodward, T.E. Persistence of Rickettsia rickettsii in a patient recovered from Rocky Mountain spotted fever. J. Immunol. 1954, 73, 383–386. [Google Scholar]
- Hove, M.G.; Walker, D.H. Persistence of rickettsiae in the partially viable gangrenous margins of amputated extremities 5 to 7 weeks after onset of Rocky Mountain spotted fever. Arch. Pathol. Lab. Med. 1995, 119, 429–431. [Google Scholar]
- Chung, M.H.; Lee, J.S.; Baek, J.H.; Kim, M.; Kang, J.S. Persistence of Orientia tsutsugamushi in humans. J. Korean Med. Sci. 2012, 27, 231–235. [Google Scholar] [CrossRef]
- Papp, S.; Moderzynski, K.; Rauch, J.; Heine, L.; Kuehl, S.; Richardt, U.; Mueller, H.; Fleischer, B.; Osterloh, A. Liver Necrosis and Lethal Systemic Inflammation in a Murine Model of Rickettsia typhi Infection: Role of Neutrophils, Macrophages and NK Cells. PLoS Negl. Trop. Dis. 2016, 10, e0004935. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.M.; Whitworth, T.; Olano, J.P.; Popov, V.L.; Walker, D.H. Fc-dependent polyclonal antibodies and antibodies to outer membrane proteins A and B, but not to lipopolysaccharide, protect SCID mice against fatal Rickettsia conorii infection. Infect. Immun. 2004, 72, 2222–2228. [Google Scholar] [CrossRef]
- Walker, D.H.; Olano, J.P.; Feng, H.M. Critical role of cytotoxic T lymphocytes in immune clearance of rickettsial infection. Infect. Immun. 2001, 69, 1841–1846. [Google Scholar] [CrossRef] [PubMed]
- Moderzynski, K.; Heine, L.; Rauch, J.; Papp, S.; Kuehl, S.; Richardt, U.; Fleischer, B.; Osterloh, A. Cytotoxic effector functions of T cells are not required for protective immunity against fatal Rickettsia typhi infection in a murine model of infection: Role of TH1 and TH17 cytokines in protection and pathology. PLoS Negl. Trop. Dis. 2017, 11, e0005404. [Google Scholar] [CrossRef]
- Moderzynski, K.; Papp, S.; Rauch, J.; Heine, L.; Kuehl, S.; Richardt, U.; Fleischer, B.; Osterloh, A. CD4+ T Cells Are as Protective as CD8+ T Cells against Rickettsia typhi Infection by Activating Macrophage Bactericidal Activity. PLoS Negl. Trop. Dis. 2016, 10, e0005089. [Google Scholar] [CrossRef]
- Hauptmann, M.; Kolbaum, J.; Lilla, S.; Wozniak, D.; Gharaibeh, M.; Fleischer, B.; Keller, C.A. Protective and Pathogenic Roles of CD8+ T Lymphocytes in Murine Orientia tsutsugamushi Infection. PLoS Negl. Trop. Dis. 2016, 10, e0004991. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Mendell, N.L.; Liang, Y.; Shelite, T.R.; Goez-Rivillas, Y.; Soong, L.; Bouyer, D.H.; Walker, D.H. CD8+ T cells provide immune protection against murine disseminated endotheliotropic Orientia tsutsugamushi infection. PLoS Negl. Trop. Dis. 2017, 11, e0005763. [Google Scholar] [CrossRef]
- Feng, H.; Popov, V.L.; Yuoh, G.; Walker, D.H. Role of T lymphocyte subsets in immunity to spotted fever group Rickettsiae. J. Immunol. 1997, 158, 5314–5320. [Google Scholar]
- Walker, D.H.; Popov, V.L.; Feng, H.M. Establishment of a novel endothelial target mouse model of a typhus group rickettsiosis: Evidence for critical roles for gamma interferon and CD8 T lymphocytes. Lab. Investig. A J. Tech. Methods Pathol. 2000, 80, 1361–1372. [Google Scholar] [CrossRef][Green Version]
- Feng, H.M.; Walker, D.H. Mechanisms of intracellular killing of Rickettsia conorii in infected human endothelial cells, hepatocytes, and macrophages. Infect. Immun. 2000, 68, 6729–6736. [Google Scholar] [CrossRef]
- Turco, J.; Winkler, H.H. Gamma-interferon-induced inhibition of the growth of Rickettsia prowazekii in fibroblasts cannot be explained by the degradation of tryptophan or other amino acids. Infect. Immun. 1986, 53, 38–46. [Google Scholar] [CrossRef]
- Feng, H.M.; Popov, V.L.; Walker, D.H. Depletion of gamma interferon and tumor necrosis factor alpha in mice with Rickettsia conorii-infected endothelium: Impairment of rickettsicidal nitric oxide production resulting in fatal, overwhelming rickettsial disease. Infect. Immun. 1994, 62, 1952–1960. [Google Scholar] [CrossRef] [PubMed]
- Dumler, J.S.; Taylor, J.P.; Walker, D.H. Clinical and laboratory features of murine typhus in south Texas, 1980 through 1987. JAMA 1991, 266, 1365–1370. [Google Scholar] [CrossRef] [PubMed]
- Fournier, P.E.; Jensenius, M.; Laferl, H.; Vene, S.; Raoult, D. Kinetics of antibody responses in Rickettsia africae and Rickettsia conorii infections. Clin. Diagn. Lab. Immunol. 2002, 9, 324–328. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Tantibhedhyangkul, W.; Prachason, T.; Waywa, D.; El Filali, A.; Ghigo, E.; Thongnoppakhun, W.; Raoult, D.; Suputtamongkol, Y.; Capo, C.; Limwongse, C.; et al. Orientia tsutsugamushi stimulates an original gene expression program in monocytes: Relationship with gene expression in patients with scrub typhus. PLoS Negl. Trop. Dis. 2011, 5, e1028. [Google Scholar] [CrossRef]
- Koo, J.E.; Hong, H.J.; Dearth, A.; Kobayashi, K.S.; Koh, Y.S. Intracellular invasion of Orientia tsutsugamushi activates inflammasome in asc-dependent manner. PLoS ONE 2012, 7, e39042. [Google Scholar] [CrossRef]
- Tantibhedhyangkul, W.; Ben Amara, A.; Textoris, J.; Gorvel, L.; Ghigo, E.; Capo, C.; Mege, J.L. Orientia tsutsugamushi, the causative agent of scrub typhus, induces an inflammatory program in human macrophages. Microb. Pathog. 2013, 55, 55–63. [Google Scholar] [CrossRef]
- Ogawa, M.; Satoh, M.; Kataoka, M.; Ando, S.; Saijo, M. Nitric oxide enhanced the growth of an obligated intracellular bacterium Orientia tsutsugamushi in murine macrophages. Microb. Pathog. 2017, 107, 335–340. [Google Scholar] [CrossRef] [PubMed]
- Gharaibeh, M.; Hagedorn, M.; Lilla, S.; Hauptmann, M.; Heine, H.; Fleischer, B.; Keller, C. Toll-Like Receptor 2 Recognizes Orientia tsutsugamushi and Increases Susceptibility to Murine Experimental Scrub Typhus. Infect. Immun. 2016, 84, 3379–3387. [Google Scholar] [CrossRef] [PubMed]
- Peter, J.V.; Griffith, M.F.; Prakash, J.A.; Chrispal, A.; Pichamuthu, K.; Varghese, G.M. Anti-nuclear antibody expression in severe scrub typhus infection: Preliminary observations. J. Glob. Infect. Dis. 2014, 6, 195–196. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.J.; Lee, M.S.; Ki, M.; Ihm, C.; Kim, D.; Kim, Y.; Yoo, S.M. Does IL-17 play a role in hepatic dysfunction of scrub typhus patients? Vector Borne Zoonotic Dis. 2010, 10, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Weigl, R.L. Die Methoden der aktiven Fleckfieberimmunisierung. Bull. Int. Acad. Pol. Sci. Et Lett. 1930, 7, 25–62. [Google Scholar]
- Weigl, R. Immunization against typhus fever in Poland during World War II. Tex. Rep. Biol. Med. 1947, 5, 177–179. [Google Scholar]
- Walker, D.H. The realities of biodefense vaccines against Rickettsia. Vaccine 2009, 27 (Suppl. 4), D52–D55. [Google Scholar] [CrossRef]
- Zinsser, H.; Castaneda, M.R. Studies on Typhus Fever: Vii. Active Immunization against Mexican Typhus Fever with Dead Virus. J. Exp. Med. 1931, 53, 493–497. [Google Scholar] [CrossRef][Green Version]
- Veintemillas, F. Vaccination against typhus fever with the Zinsser-Castaneda Vaccine. J. Immunol. 1939, 36, 339–348. [Google Scholar]
- Spencer, R.R.; Parker, R.R. Rocky Mountain spotted fever: Vaccination of monkeys and man. Public Health Rep. 1925, 40, 2159–2167. [Google Scholar] [CrossRef]
- Ecke, R.S.; Gilliam, A.G.; Snyder, J.C.; Yeomans, A.; Zarafonetis, C.J.; Murray, E.S. The effect of Cox-type vaccine on louse-borne typhus fever; an account of 61 cases of naturally occurring typhus fever in patients who had previously received one or more injections of Cox-type vaccine. Am. J. Trop. Med. Hyg. 1945, 25, 447–462. [Google Scholar] [CrossRef] [PubMed]
- DuPont, H.L.; Hornick, R.B.; Dawkins, A.T.; Heiner, G.G.; Fabrikant, I.B.; Wisseman, C.L., Jr.; Woodward, T.E. Rocky Mountain spotted fever: A comparative study of the active immunity induced by inactivated and viable pathogenic Rickettsia rickettsii. J. Infect. Dis. 1973, 128, 340–344. [Google Scholar] [CrossRef] [PubMed]
- Kenyon, R.H.; Pedersen, C.E., Jr. Preparation of Rocky Mountain spotted fever vaccine suitable for human immunization. J. Clin. Microbiol. 1975, 1, 500–503. [Google Scholar] [CrossRef] [PubMed]
- Kenyon, R.H.; Sammons, L.S.; Pedersen, C.E., Jr. Comparison of three rocky mountain spotted fever vaccines. J. Clin. Microbiol. 1975, 2, 300–304. [Google Scholar] [CrossRef]
- Gonder, J.C.; Kenyon, R.H.; Pedersen, C.E., Jr. Evaluation of a killed Rocky Mountain spotted fever vaccine in cynomolgus monkeys. J. Clin. Microbiol. 1979, 10, 719–723. [Google Scholar] [CrossRef]
- Maugh, T.H., 2nd. Rickettsiae: A new vaccine for Rocky Mountain spotted fever. Science 1978, 201, 604. [Google Scholar] [CrossRef]
- Clements, M.L.; Wisseman, C.L., Jr.; Woodward, T.E.; Fiset, P.; Dumler, J.S.; McNamee, W.; Black, R.E.; Rooney, J.; Hughes, T.P.; Levine, M.M. Reactogenicity, immunogenicity, and efficacy of a chick embryo cell-derived vaccine for Rocky Mountain spotted fever. J. Infect. Dis. 1983, 148, 922–930. [Google Scholar] [CrossRef]
- Buckland, F.E.; Dudgeon, A. Scrubtyphus vaccine; large-scale production. Lancet 1945, 2, 734–737. [Google Scholar] [CrossRef]
- Card, W.I.; Walker, J.M. Scrub-typhus vaccine; field trial in South-east Asia. Lancet 1947, 1, 481–483. [Google Scholar] [CrossRef]
- Berge, T.O.; Gauld, R.L.; Kitaoka, M. A field trial of a vaccine prepared from the Volner strain of Rickettsia tsutsugamushi. Am. J. Hyg. 1949, 50, 337–342. [Google Scholar] [CrossRef]
- Bailey, C.A.; Diercks, F.H.; Proffitt, J.E. Preparation of a serological antigen and a vaccine for experimental tsutsugamushi disease. J. Immunol. 1948, 60, 431–441. [Google Scholar]
- Rights, F.L.; Smadel, J.E. Studies on scrib typhus; tsutsugamushi disease; heterogenicity of strains of R. tsutsugamushi as demonstrated by cross-vaccination studies. J. Exp. Med. 1948, 87, 339–351. [Google Scholar] [CrossRef]
- Choi, Y.; Kim, K.S.; Kim, T.Y.; Cheong, H.S.; Ahn, B.Y. Long-term egg-yolk adaptation of the Orientia tsutsugamushi for preparation of a formalinized immunogen. Vaccine 2006, 24, 1438–1445. [Google Scholar] [CrossRef] [PubMed]
- Eisenberg, G.H., Jr.; Osterman, J.V. Gamma-irradiated scrub typhus immunogens: Broad-spectrum immunity with combinations of rickettsial strains. Infect. Immun. 1979, 26, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Eisenberg, G.H., Jr.; Osterman, J.V. Gamma-irradiated scrub typhus immunogens: Development and duration of immunity. Infect. Immun. 1978, 22, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Eisenberg, G.H., Jr.; Osterman, J.V. Experimental scrub typhus immunogens: Gamma-irradiated and formalinized rickettsiae. Infect. Immun. 1977, 15, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Alhassan, A.; Liu, H.; McGill, J.; Cerezo, A.; Jakkula, L.; Nair, A.D.S.; Winkley, E.; Olson, S.; Marlow, D.; Sahni, A.; et al. Rickettsia rickettsii Whole-Cell Antigens Offer Protection against Rocky Mountain Spotted Fever in the Canine Host. Infect. Immun. 2019, 87, e00628-18. [Google Scholar] [CrossRef]
- Kawamura, R.; Kasahar, S.; Toyama, T.; Nishinarita, F.; Tsubaki, S. On the prevention of tsutsugamushi. Results of preventive inoculations for people in the endemic region, and laboratory tests with the Pescadores strain. Trop. Dis. Bull. 1940, 37, 269–270. [Google Scholar]
- Fox, J.P.; Jordan, M.E.; Gelfand, H.M. Immunization of man against epidemic typhus by infection with avirulent Rickettsia prowazeki strain E. IV. Persistence of immunity and a note as to differing complement-fixation antigen requirements in post-infection and post-vaccination sera. J. Immunol. 1957, 79, 348–354. [Google Scholar] [PubMed]
- Wisseman, C.L., Jr. Concepts of louse-borne typhus control in developing countries: The use of the living attenuated E strain typhus vaccine in epidemic and endemic situations. Adv. Exp. Med. Biol. 1972, 31, 97–130. [Google Scholar]
- Ching, W.M.; Wang, H.; Davis, J.; Dasch, G.A. Amino acid analysis and multiple methylation of lysine residues in the surface protein antigen of Rickettsia prowazekii. In Techniques in Protein Chemistry IV; Angeletti, R.H., Ed.; Academic Press, Inc.: San Diego, CA, USA, 1993; pp. 307–314. [Google Scholar]
- Zhang, J.Z.; Hao, J.F.; Walker, D.H.; Yu, X.J. A mutation inactivating the methyltransferase gene in avirulent Madrid E strain of Rickettsia prowazekii reverted to wild type in the virulent revertant strain Evir. Vaccine 2006, 24, 2317–2323. [Google Scholar] [CrossRef]
- Driskell, L.O.; Yu, X.J.; Zhang, L.; Liu, Y.; Popov, V.L.; Walker, D.H.; Tucker, A.M.; Wood, D.O. Directed mutagenesis of the Rickettsia prowazekii pld gene encoding phospholipase D. Infect. Immun. 2009, 77, 3244–3248. [Google Scholar] [CrossRef]
- Whitworth, T.; Popov, V.L.; Yu, X.J.; Walker, D.H.; Bouyer, D.H. Expression of the Rickettsia prowazekii pld or tlyC gene in Salmonella enterica serovar Typhimurium mediates phagosomal escape. Infect. Immun. 2005, 73, 6668–6673. [Google Scholar] [CrossRef] [PubMed]
- Noriea, N.F.; Clark, T.R.; Hackstadt, T. Targeted knockout of the Rickettsia rickettsii OmpA surface antigen does not diminish virulence in a mammalian model system. MBio 2015, 6, e00323-15. [Google Scholar] [CrossRef] [PubMed]
- Teysseire, N.; Raoult, D. Comparison of Western immunoblotting and microimmunofluorescence for diagnosis of Mediterranean spotted fever. J. Clin. Microbiol. 1992, 30, 455–460. [Google Scholar] [CrossRef] [PubMed]
- Anacker, R.L.; List, R.H.; Mann, R.E.; Hayes, S.F.; Thomas, L.A. Characterization of monoclonal antibodies protecting mice against Rickettsia rickettsii. J. Infect. Dis. 1985, 151, 1052–1060. [Google Scholar] [CrossRef]
- Anacker, R.L.; McDonald, G.A.; List, R.H.; Mann, R.E. Neutralizing activity of monoclonal antibodies to heat-sensitive and heat-resistant epitopes of Rickettsia rickettsii surface proteins. Infect. Immun. 1987, 55, 825–827. [Google Scholar] [CrossRef]
- Lange, J.V.; Walker, D.H. Production and characterization of monoclonal antibodies to Rickettsia rickettsii. Infect. Immun. 1984, 46, 289–294. [Google Scholar] [CrossRef]
- Chan, Y.G.; Riley, S.P.; Chen, E.; Martinez, J.J. Molecular basis of immunity to rickettsial infection conferred through outer membrane protein B. Infect. Immun. 2011, 79, 2303–2313. [Google Scholar] [CrossRef]
- Feng, H.M.; Whitworth, T.; Popov, V.; Walker, D.H. Effect of antibody on the rickettsia-host cell interaction. Infect. Immun. 2004, 72, 3524–3530. [Google Scholar] [CrossRef]
- Li, H.; Walker, D.H. rOmpA is a critical protein for the adhesion of Rickettsia rickettsii to host cells. Microb. Pathog. 1998, 24, 289–298. [Google Scholar] [CrossRef]
- Qi, Y.; Xiong, X.; Wang, X.; Duan, C.; Jia, Y.; Jiao, J.; Gong, W.; Wen, B. Proteome analysis and serological characterization of surface-exposed proteins of Rickettsia heilongjiangensis. PLoS ONE 2013, 8, e70440. [Google Scholar] [CrossRef]
- Rauch, J.; Barton, J.; Kwiatkowski, M.; Wunderlich, M.; Steffen, P.; Moderzynski, K.; Papp, S.; Höhn, K.; Schwanke, H.; Witt, S.; et al. GroEL is an immunodominant surface-exposed antigen of Rickettsia typhi. PLoS ONE 2021, 16, e0253084. [Google Scholar] [CrossRef]
- Park, H.; Lee, J.H.; Gouin, E.; Cossart, P.; Izard, T. The rickettsia surface cell antigen 4 applies mimicry to bind to and activate vinculin. J. Biol. Chem. 2011, 286, 35096–35103. [Google Scholar] [CrossRef]
- Fish, A.I.; Riley, S.P.; Singh, B.; Riesbeck, K.; Martinez, J.J. The Rickettsia conorii Adr1 Interacts with the C-Terminus of Human Vitronectin in a Salt-Sensitive Manner. Front. Cell. Infect. Microbiol. 2017, 7, 61. [Google Scholar] [CrossRef] [PubMed]
- Riley, S.P.; Patterson, J.L.; Nava, S.; Martinez, J.J. Pathogenic Rickettsia species acquire vitronectin from human serum to promote resistance to complement-mediated killing. Cell. Microbiol. 2014, 16, 849–861. [Google Scholar] [CrossRef][Green Version]
- Garza, D.A.; Riley, S.P.; Martinez, J.J. Expression of Rickettsia Adr2 protein in E. coli is sufficient to promote resistance to complement-mediated killing, but not adherence to mammalian cells. PLoS ONE 2017, 12, e0179544. [Google Scholar] [CrossRef] [PubMed]
- Gong, W.; Xiong, X.; Qi, Y.; Jiao, J.; Duan, C.; Wen, B. Identification of novel surface-exposed proteins of Rickettsia rickettsii by affinity purification and proteomics. PLoS ONE 2014, 9, e100253. [Google Scholar] [CrossRef] [PubMed]
- Renesto, P.; Azza, S.; Dolla, A.; Fourquet, P.; Vestris, G.; Gorvel, J.P.; Raoult, D. Proteome analysis of Rickettsia conorii by two-dimensional gel electrophoresis coupled with mass spectrometry. FEMS Microbiol. Lett. 2005, 245, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Osterloh, A. The neglected challenge: Vaccination against rickettsiae. PLoS Negl. Trop. Dis. 2020, 14, e0008704. [Google Scholar] [CrossRef]
- He, Y.; Racz, R.; Sayers, S.; Lin, Y.; Todd, T.; Hur, J.; Li, X.; Patel, M.; Zhao, B.; Chung, M.; et al. Updates on the web-based VIOLIN vaccine database and analysis system. Nucleic Acids Res. 2014, 42, D1124–D1132. [Google Scholar] [CrossRef]
- Doytchinova, I.A.; Flower, D.R. VaxiJen: A server for prediction of protective antigens, tumour antigens and subunit vaccines. BMC Bioinform. 2007, 8, 4. [Google Scholar] [CrossRef]
- Magnan, C.N.; Zeller, M.; Kayala, M.A.; Vigil, A.; Randall, A.; Felgner, P.L.; Baldi, P. High-throughput prediction of protein antigenicity using protein microarray data. Bioinformatics 2010, 26, 2936–2943. [Google Scholar] [CrossRef]
- Saha, S.; Raghava, G.P. Prediction of continuous B-cell epitopes in an antigen using recurrent neural network. Proteins 2006, 65, 40–48. [Google Scholar] [CrossRef]
- Schlessinger, A.; Ofran, Y.; Yachdav, G.; Rost, B. Epitome: Database of structure-inferred antigenic epitopes. Nucleic Acids Res. 2006, 34, D777–D780. [Google Scholar] [CrossRef]
- Nielsen, M.; Lundegaard, C.; Worning, P.; Lauemoller, S.L.; Lamberth, K.; Buus, S.; Brunak, S.; Lund, O. Reliable prediction of T-cell epitopes using neural networks with novel sequence representations. Protein Sci. 2003, 12, 1007–1017. [Google Scholar] [CrossRef]
- Nielsen, M.; Lundegaard, C.; Worning, P.; Hvid, C.S.; Lamberth, K.; Buus, S.; Brunak, S.; Lund, O. Improved prediction of MHC class I and class II epitopes using a novel Gibbs sampling approach. Bioinformatics 2004, 20, 1388–1397. [Google Scholar] [CrossRef]
- Lin, H.H.; Ray, S.; Tongchusak, S.; Reinherz, E.L.; Brusic, V. Evaluation of MHC class I peptide binding prediction servers: Applications for vaccine research. BMC Immunol. 2008, 9, 8. [Google Scholar] [CrossRef]
- Rammensee, H.; Bachmann, J.; Emmerich, N.P.; Bachor, O.A.; Stevanovic, S. SYFPEITHI: Database for MHC ligands and peptide motifs. Immunogenetics 1999, 50, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Reche, P.A.; Reinherz, E.L. Prediction of peptide-MHC binding using profiles. Methods Mol. Biol. 2007, 409, 185–200. [Google Scholar] [CrossRef]
- Zhang, G.L.; Srinivasan, K.N.; Veeramani, A.; August, J.T.; Brusic, V. PREDBALB/c: A system for the prediction of peptide binding to H2d molecules, a haplotype of the BALB/c mouse. Nucleic Acids Res. 2005, 33, W180–W183. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zhang, G.L.; Petrovsky, N.; Kwoh, C.K.; August, J.T.; Brusic, V. PRED(TAP): A system for prediction of peptide binding to the human transporter associated with antigen processing. Immunome Res. 2006, 2, 3. [Google Scholar] [CrossRef][Green Version]
- Guan, P.; Doytchinova, I.A.; Zygouri, C.; Flower, D.R. MHCPred: Bringing a quantitative dimension to the online prediction of MHC binding. Appl. Bioinform. 2003, 2, 63–66. [Google Scholar]
- Imai, K.; Asakawa, N.; Tsuji, T.; Akazawa, F.; Ino, A.; Sonoyama, M.; Mitaku, S. SOSUI-GramN: High performance prediction for sub-cellular localization of proteins in gram-negative bacteria. Bioinformation 2008, 2, 417–421. [Google Scholar] [CrossRef]
- Gardy, J.L.; Spencer, C.; Wang, K.; Ester, M.; Tusnady, G.E.; Simon, I.; Hua, S.; deFays, K.; Lambert, C.; Nakai, K.; et al. PSORT-B: Improving protein subcellular localization prediction for Gram-negative bacteria. Nucleic Acids Res. 2003, 31, 3613–3617. [Google Scholar] [CrossRef] [PubMed]
- Almagro Armenteros, J.J.; Tsirigos, K.D.; Sonderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef] [PubMed]
- Bendtsen, J.D.; Jensen, L.J.; Blom, N.; Von Heijne, G.; Brunak, S. Feature-based prediction of non-classical and leaderless protein secretion. Protein Eng. Des. Sel. PEDS 2004, 17, 349–356. [Google Scholar] [CrossRef]
- Gazi, M.; Caro-Gomez, E.; Goez, Y.; Cespedes, M.A.; Hidalgo, M.; Correa, P.; Valbuena, G. Discovery of a protective Rickettsia prowazekii antigen recognized by CD8+ T cells, RP884, using an in vivo screening platform. PLoS ONE 2013, 8, e76253. [Google Scholar] [CrossRef]
- Caro-Gomez, E.; Gazi, M.; Goez, Y.; Valbuena, G. Discovery of novel cross-protective Rickettsia prowazekii T-cell antigens using a combined reverse vaccinology and in vivo screening approach. Vaccine 2014, 32, 4968–4976. [Google Scholar] [CrossRef]
- Dzul-Rosado, K.; Balam-Romero, J.; Valencia-Pacheco, G.; Lugo-Caballero, C.; Arias-Leon, J.; Peniche-Lara, G.; Zavala-Castro, J. Immunogenicity of OmpA and OmpB antigens from Rickettsia rickettsii on mononuclear cells from Rickettsia positive Mexican patients. J. Vector Borne Dis. 2017, 54, 317–327. [Google Scholar] [CrossRef]
- Sumner, J.W.; Sims, K.G.; Jones, D.C.; Anderson, B.E. Protection of guinea-pigs from experimental Rocky Mountain spotted fever by immunization with baculovirus-expressed Rickettsia rickettsii rOmpA protein. Vaccine 1995, 13, 29–35. [Google Scholar] [CrossRef]
- Jiao, Y.; Wen, B.; Chen, M.; Niu, D.; Zhang, J.; Qiu, L. Analysis of immunoprotectivity of the recombinant OmpA of Rickettsia heilongjiangensis. Ann. N. Y. Acad. Sci. 2005, 1063, 261–265. [Google Scholar] [CrossRef] [PubMed]
- Vishwanath, S.; McDonald, G.A.; Watkins, N.G. A recombinant Rickettsia conorii vaccine protects guinea pigs from experimental boutonneuse fever and Rocky Mountain spotted fever. Infect. Immun. 1990, 58, 646–653. [Google Scholar] [CrossRef]
- Bourgeois, A.L.; Dasch, G.A. The species-specific surface protein antigen of Rickettsia typhi: Immunogenicity and protective efficacy in guinea pigs. In Rickettsiae and Rickettsial Diseases; Burgdorfer, W., Anacker, R.L., Eds.; Academic Press: New York, NY, USA, 1981; pp. 71–80. [Google Scholar]
- Ching, W.M.; Wang, H.; Jan, B.; Dasch, G.A. Identification and characterization of epitopes on the 120-kilodalton surface protein antigen of Rickettsia prowazekii with synthetic peptides. Infect. Immun. 1996, 64, 1413–1419. [Google Scholar] [CrossRef] [PubMed]
- Sears, K.T.; Ceraul, S.M.; Gillespie, J.J.; Allen, E.D., Jr.; Popov, V.L.; Ammerman, N.C.; Rahman, M.S.; Azad, A.F. Surface proteome analysis and characterization of surface cell antigen (Sca) or autotransporter family of Rickettsia typhi. PLoS Pathog. 2012, 8, e1002856. [Google Scholar] [CrossRef]
- Wang, P.; Xiong, X.; Jiao, J.; Yang, X.; Jiang, Y.; Wen, B.; Gong, W. Th1 epitope peptides induce protective immunity against Rickettsia rickettsii infection in C3H/HeN mice. Vaccine 2017, 35, 7204–7212. [Google Scholar] [CrossRef]
- Gong, W.; Qi, Y.; Xiong, X.; Jiao, J.; Duan, C.; Wen, B. Rickettsia rickettsii outer membrane protein YbgF induces protective immunity in C3H/HeN mice. Hum. Vaccines Immunother. 2015, 11, 642–649. [Google Scholar] [CrossRef]
- Qi, Y.; Xiong, X.; Duan, C.; Jiao, J.; Gong, W.; Wen, B. Recombinant protein YbgF induces protective immunity against Rickettsia heilongjiangensis infection in C3H/HeN mice. Vaccine 2013, 31, 5643–5650. [Google Scholar] [CrossRef]
- Gong, W.; Xiong, X.; Qi, Y.; Jiao, J.; Duan, C.; Wen, B. Surface protein Adr2 of Rickettsia rickettsii induced protective immunity against Rocky Mountain spotted fever in C3H/HeN mice. Vaccine 2014, 32, 2027–2033. [Google Scholar] [CrossRef]
- Gong, W.; Wang, P.; Xiong, X.; Jiao, J.; Yang, X.; Wen, B. Enhanced protection against Rickettsia rickettsii infection in C3H/HeN mice by immunization with a combination of a recombinant adhesin rAdr2 and a protein fragment rOmpB-4 derived from outer membrane protein B. Vaccine 2015, 33, 985–992. [Google Scholar] [CrossRef]
- Chen, W.J.; Niu, D.S.; Zhang, X.Y.; Chen, M.L.; Cui, H.; Wei, W.J.; Wen, B.H.; Chen, X.R. Recombinant 56-kilodalton major outer membrane protein antigen of Orientia tsutsugamushi Shanxi and its antigenicity. Infect. Immun. 2003, 71, 4772–4779. [Google Scholar] [CrossRef]
- Seong, S.Y.; Park, S.G.; Huh, M.S.; Jang, W.J.; Kim, H.R.; Han, T.H.; Choi, M.S.; Chang, W.H.; Kim, I.S. Mapping of antigenic determinant regions of the Bor56 protein of Orientia tsutsugamushi. Infect. Immun. 1997, 65, 5250–5256. [Google Scholar] [CrossRef]
- Ramaiah, A.; Koralur, M.C.; Dasch, G.A. Complexity of type-specific 56 kDa antigen CD4 T-cell epitopes of Orientia tsutsugamushi strains causing scrub typhus in India. PLoS ONE 2018, 13, e0196240. [Google Scholar] [CrossRef]
- Chen, H.W.; Zhang, Z.; Huber, E.; Mutumanje, E.; Chao, C.C.; Ching, W.M. Kinetics and magnitude of antibody responses against the conserved 47-kilodalton antigen and the variable 56-kilodalton antigen in scrub typhus patients. Clin. Vaccine Immunol. 2011, 18, 1021–1027. [Google Scholar] [CrossRef]
- Seong, S.Y.; Huh, M.S.; Jang, W.J.; Park, S.G.; Kim, J.G.; Woo, S.G.; Choi, M.S.; Kim, I.S.; Chang, W.H. Induction of homologous immune response to Rickettsia tsutsugamushi Boryong with a partial 56-kilodalton recombinant antigen fused with the maltose-binding protein MBP-Bor56. Infect. Immun. 1997, 65, 1541–1545. [Google Scholar] [CrossRef]
- Seong, S.Y.; Kim, H.R.; Huh, M.S.; Park, S.G.; Kang, J.S.; Han, T.H.; Choi, M.S.; Chang, W.H.; Kim, I.S. Induction of neutralizing antibody in mice by immunization with recombinant 56 kDa protein of Orientia tsutsugamushi. Vaccine 1997, 15, 1741–1747. [Google Scholar] [CrossRef]
- Choi, S.; Jeong, H.J.; Ju, Y.R.; Gill, B.; Hwang, K.J.; Lee, J. Protective immunity of 56-kDa type-specific antigen of Orientia tsutsugamushi causing scrub typhus. J. Microbiol. Biotechnol. 2014, 24, 1728–1735. [Google Scholar] [CrossRef]
- Kim, H.I.; Ha, N.Y.; Kim, G.; Min, C.K.; Kim, Y.; Yen, N.T.H.; Choi, M.S.; Cho, N.H. Immunization with a recombinant antigen composed of conserved blocks from TSA56 provides broad genotype protection against scrub typhus. Emerg. Microbes Infect. 2019, 8, 946–958. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Wen, B.; Wen, B.; Niu, D.; Chen, M.; Qiu, L. Induction of protective immunity against scrub typhus with a 56-kilodalton recombinant antigen fused with a 47-kilodalton antigen of Orientia tsutsugamushi Karp. Am. J. Trop. Med. Hyg. 2005, 72, 458–464. [Google Scholar] [CrossRef]
- Chattopadhyay, S.; Jiang, J.; Chan, T.C.; Manetz, T.S.; Chao, C.C.; Ching, W.M.; Richards, A.L. Scrub typhus vaccine candidate Kp r56 induces humoral and cellular immune responses in cynomolgus monkeys. Infect. Immun. 2005, 73, 5039–5047. [Google Scholar] [CrossRef]
- Hickman, C.J.; Stover, C.K.; Joseph, S.W.; Oaks, E.V. Molecular cloning and sequence analysis of a Rickettsia tsutsugamushi 22 kDa antigen containing B- and T-cell epitopes. Microb. Pathog. 1991, 11, 19–31. [Google Scholar] [CrossRef]
- Ha, N.Y.; Kim, Y.; Choi, J.H.; Choi, M.S.; Kim, I.S.; Kim, Y.S.; Cho, N.H. Detection of antibodies against Orientia tsutsugamushi Sca proteins in scrub typhus patients and genetic variation of sca genes of different strains. Clin. Vaccine Immunol. 2012, 19, 1442–1451. [Google Scholar] [CrossRef]
- Ha, N.Y.; Sharma, P.; Kim, G.; Kim, Y.; Min, C.K.; Choi, M.S.; Kim, I.S.; Cho, N.H. Immunization with an autotransporter protein of Orientia tsutsugamushi provides protective immunity against scrub typhus. PLoS Negl. Trop. Dis. 2015, 9, e0003585. [Google Scholar] [CrossRef]
- Ha, N.Y.; Shin, H.M.; Sharma, P.; Cho, H.A.; Min, C.K.; Kim, H.I.; Yen, N.T.; Kang, J.S.; Kim, I.S.; Choi, M.S.; et al. Generation of protective immunity against Orientia tsutsugamushi infection by immunization with a zinc oxide nanoparticle combined with ScaA antigen. J. Nanobiotechnol. 2016, 14, 76. [Google Scholar] [CrossRef]
- Kheirollahpour, M.; Mehrabi, M.; Dounighi, N.M.; Mohammadi, M.; Masoudi, A. Nanoparticles and Vaccine Development. Pharm. Nanotechnol. 2020, 8, 6–21. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.H.; Youn, J.W.; Sung, Y.C. Cross-priming as a predominant mechanism for inducing CD8(+) T cell responses in gene gun DNA immunization. J. Immunol. 2001, 167, 5549–5557. [Google Scholar] [CrossRef] [PubMed]
- Kutzler, M.A.; Weiner, D.B. Developing DNA vaccines that call to dendritic cells. J. Clin. Investig. 2004, 114, 1241–1244. [Google Scholar] [CrossRef] [PubMed]
- Crocquet-Valdes, P.A.; Diaz-Montero, C.M.; Feng, H.M.; Li, H.; Barrett, A.D.; Walker, D.H. Immunization with a portion of rickettsial outer membrane protein A stimulates protective immunity against spotted fever rickettsiosis. Vaccine 2001, 20, 979–988. [Google Scholar] [CrossRef]
- Diaz-Montero, C.M.; Feng, H.M.; Crocquet-Valdes, P.A.; Walker, D.H. Identification of protective components of two major outer membrane proteins of spotted fever group Rickettsiae. Am. J. Trop. Med. Hyg. 2001, 65, 371–378. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ni, Y.S.; Chan, T.C.; Chao, C.C.; Richards, A.L.; Dasch, G.A.; Ching, W.M. Protection against scrub typhus by a plasmid vaccine encoding the 56-KD outer membrane protein antigen gene. Am. J. Trop. Med. Hyg. 2005, 73, 936–941. [Google Scholar] [CrossRef] [PubMed]
- Maruggi, G.; Zhang, C.; Li, J.; Ulmer, J.B.; Yu, D. mRNA as a Transformative Technology for Vaccine Development to Control Infectious Diseases. Mol. Ther. J. Am. Soc. Gene Ther. 2019, 27, 757–772. [Google Scholar] [CrossRef]
- Scheel, B.; Teufel, R.; Probst, J.; Carralot, J.P.; Geginat, J.; Radsak, M.; Jarrossay, D.; Wagner, H.; Jung, G.; Rammensee, H.G.; et al. Toll-like receptor-dependent activation of several human blood cell types by protamine-condensed mRNA. Eur. J. Immunol. 2005, 35, 1557–1566. [Google Scholar] [CrossRef] [PubMed]
- Kallen, K.J.; Heidenreich, R.; Schnee, M.; Petsch, B.; Schlake, T.; Thess, A.; Baumhof, P.; Scheel, B.; Koch, S.D.; Fotin-Mleczek, M. A novel, disruptive vaccination technology: Self-adjuvanted RNActive((R)) vaccines. Hum. Vaccines Immunother. 2013, 9, 2263–2276. [Google Scholar] [CrossRef]
- Rauch, S.; Lutz, J.; Kowalczyk, A.; Schlake, T.; Heidenreich, R. RNActive(R) Technology: Generation and Testing of Stable and Immunogenic mRNA Vaccines. Methods Mol. Biol. 2017, 1499, 89–107. [Google Scholar] [CrossRef]
- Chen, N.; Xia, P.; Li, S.; Zhang, T.; Wang, T.T.; Zhu, J. RNA sensors of the innate immune system and their detection of pathogens. IUBMB Life 2017, 69, 297–304. [Google Scholar] [CrossRef]
- Kowalczyk, A.; Doener, F.; Zanzinger, K.; Noth, J.; Baumhof, P.; Fotin-Mleczek, M.; Heidenreich, R. Self-adjuvanted mRNA vaccines induce local innate immune responses that lead to a potent and boostable adaptive immunity. Vaccine 2016, 34, 3882–3893. [Google Scholar] [CrossRef]
- Edwards, D.K.; Jasny, E.; Yoon, H.; Horscroft, N.; Schanen, B.; Geter, T.; Fotin-Mleczek, M.; Petsch, B.; Wittman, V. Adjuvant effects of a sequence-engineered mRNA vaccine: Translational profiling demonstrates similar human and murine innate response. J. Transl. Med. 2017, 15, 1. [Google Scholar] [CrossRef]
- Li, Z.; Diaz-Montero, C.M.; Valbuena, G.; Yu, X.J.; Olano, J.P.; Feng, H.M.; Walker, D.H. Identification of CD8 T-lymphocyte epitopes in OmpB of Rickettsia conorii. Infect. Immun. 2003, 71, 3920–3926. [Google Scholar] [CrossRef]
- Saade, F.; Petrovsky, N. Technologies for enhanced efficacy of DNA vaccines. Expert Rev. Vaccines 2012, 11, 189–209. [Google Scholar] [CrossRef]
- Harro, C.; Sun, X.; Stek, J.E.; Leavitt, R.Y.; Mehrotra, D.V.; Wang, F.; Bett, A.J.; Casimiro, D.R.; Shiver, J.W.; DiNubile, M.J.; et al. Safety and immunogenicity of the Merck adenovirus serotype 5 (MRKAd5) and MRKAd6 human immunodeficiency virus type 1 trigene vaccines alone and in combination in healthy adults. Clin. Vaccine Immunol. 2009, 16, 1285–1292. [Google Scholar] [CrossRef]
- Hill, A.V.; Reyes-Sandoval, A.; O’Hara, G.; Ewer, K.; Lawrie, A.; Goodman, A.; Nicosia, A.; Folgori, A.; Colloca, S.; Cortese, R.; et al. Prime-boost vectored malaria vaccines: Progress and prospects. Hum. Vaccin 2010, 6, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Radosevic, K.; Wieland, C.W.; Rodriguez, A.; Weverling, G.J.; Mintardjo, R.; Gillissen, G.; Vogels, R.; Skeiky, Y.A.; Hone, D.M.; Sadoff, J.C.; et al. Protective immune responses to a recombinant adenovirus type 35 tuberculosis vaccine in two mouse strains: CD4 and CD8 T-cell epitope mapping and role of gamma interferon. Infect. Immun. 2007, 75, 4105–4115. [Google Scholar] [CrossRef] [PubMed]
- McCoy, K.; Tatsis, N.; Korioth-Schmitz, B.; Lasaro, M.O.; Hensley, S.E.; Lin, S.W.; Li, Y.; Giles-Davis, W.; Cun, A.; Zhou, D.; et al. Effect of preexisting immunity to adenovirus human serotype 5 antigens on the immune responses of nonhuman primates to vaccine regimens based on human- or chimpanzee-derived adenovirus vectors. J. Virol. 2007, 81, 6594–6604. [Google Scholar] [CrossRef]
- Pichla-Gollon, S.L.; Lin, S.W.; Hensley, S.E.; Lasaro, M.O.; Herkenhoff-Haut, L.; Drinker, M.; Tatsis, N.; Gao, G.P.; Wilson, J.M.; Ertl, H.C.; et al. Effect of preexisting immunity on an adenovirus vaccine vector: In vitro neutralization assays fail to predict inhibition by antiviral antibody in vivo. J. Virol. 2009, 83, 5567–5573. [Google Scholar] [CrossRef]
- Xiang, Z.; Li, Y.; Cun, A.; Yang, W.; Ellenberg, S.; Switzer, W.M.; Kalish, M.L.; Ertl, H.C. Chimpanzee adenovirus antibodies in humans, sub-Saharan Africa. Emerg. Infect. Dis. 2006, 12, 1596–1599. [Google Scholar] [CrossRef] [PubMed]
- Gong, W.; Liang, Y.; Wu, X. The current status, challenges, and future developments of new tuberculosis vaccines. Hum. Vaccines Immunother. 2018, 14, 1697–1716. [Google Scholar] [CrossRef]
- Skinner, M.A.; Yuan, S.; Prestidge, R.; Chuk, D.; Watson, J.D.; Tan, P.L. Immunization with heat-killed Mycobacterium vaccae stimulates CD8+ cytotoxic T cells specific for macrophages infected with Mycobacterium tuberculosis. Infect. Immun. 1997, 65, 4525–4530. [Google Scholar] [CrossRef]
- Zhang, L.; Jiang, Y.; Cui, Z.; Yang, W.; Yue, L.; Ma, Y.; Shi, S.; Wang, C.; Wang, C.; Qian, A. Mycobacterium vaccae induces a strong Th1 response that subsequently declines in C57BL/6 mice. J. Vet. Sci. 2016, 17, 505–513. [Google Scholar] [CrossRef]
- Safar, H.A.; Mustafa, A.S.; Amoudy, H.A.; El-Hashim, A. The effect of adjuvants and delivery systems on Th1, Th2, Th17 and Treg cytokine responses in mice immunized with Mycobacterium tuberculosis-specific proteins. PLoS ONE 2020, 15, e0228381. [Google Scholar] [CrossRef] [PubMed]
- Meng, Y.; Xiong, X.; Qi, Y.; Duan, C.; Gong, W.; Jiao, J.; Wen, B. Protective immunity against Rickettsia heilongjiangensis in a C3H/HeN mouse model mediated by outer membrane protein B-pulsed dendritic cells. Sci. China Life Sci. 2015, 58, 287–296. [Google Scholar] [CrossRef] [PubMed][Green Version]


{kind=link}
| Genus | Group | Species | Disease | Distribution |
|---|---|---|---|---|
| Rickettsia | SFG | R. rickettsii | Rocky Mountain SF | North America (Midwest and Southeastern U.S.), Central and South America (Mexico, Panama, Costa Rica, Brazil, Argentina, Colombia) |
| R. conorii ssp. conorii | Mediterranean SF | Europe (Mediterranean Basin), North Africa (Tunisia, Algeria, Morocco), multiple sub-Saharan countries | ||
| R. conorii ssp. indica | Indian tick typhus | Middle East, India | ||
| R. conorii ssp. israelensis | Israeli SF | Israel, North Africa (Tunisia) | ||
| R. conorii ssp. israelensis | Astrakhan fever | Astrakhan region, France | ||
| R. conorii ssp. caspia | Astrakhan fever | Africa (Chad) | ||
| 364D (R. phillipi) | Unnamed rickettsiosis | U.S. (southern California) | ||
| R. honei | Flinder’s Island SF/Thailand SF | Australia, Tasmania, Thailand | ||
| R. helvetica | Tick-bite fever | Europe (Denmark, Austria, France, Italy), Asia (Laos) | ||
| R. japonica | Japanese SF | Japan, detected in ticks in South Korea and Northern Thailand | ||
| R. heilongjiangensis | Far-Eastern SF | Northern China, Russia far east, Japan, Eastern Asia | ||
| R. parkeri | Maculatum infection/American boutonneuse fever/Tidewater SF | North and South America | ||
| R. africae | African tick-bite fever | Sub-Saharan Africa, Caribbean, West Indies | ||
| R. sibirica | Siberian tick typhus/North Asian tick typhus | Russia, China, Mongolia | ||
| R. sibiria ssp. mongolotimonae | Tickborne lymphadenopathy (TIBOLA) | Southern Europe (France, Greece, Portugal, Spain), Asia, South Africa | ||
| R. massiliae | Mediterranean SF-like disease | Southern Europe, South America (Argentina) | ||
| R. monacensis | Tick-bite fever | Europe | ||
| R. slovaca | Tickborne lymphadenopathy (TIBOLA)/Dermacentor-borne necrosis and lymphadenopathy (DEBONEL)/ scalp eschar and neck lymphadenopathy after tick bite (SENLAT) | Europe (France, Slovakia, Italy, Germany, Hungary, Spain, Poland), Georgia, Russia | ||
| R. raoultii | Europe (France, Slovakia, Poland) | |||
| R. aeschlimannii | Tick-bite fever | North Africa (Tunisia, Morocco), South Africa | ||
| 364D (R. phillipi) | Unnamed rickettsiosis | Southern California | ||
| TG | R. prowazekii | Epidemic typhus | Worldwide, sporadic in Africa, Asia, Central and South America, Russia | |
| R. typhi | Endemic typhus/Murine typhus | Worldwide | ||
| transitional | R. felis | Cat-flea typhus | Probably worldwide | |
| R. akari | Rickettsialpox | Probably worldwide | ||
| R. australis | Queensland tick typhus | Australia, Tasmania | ||
| ancestral | R. bellii | |||
| R. canadensis | ||||
| Orientia | O. tsutsugamushi | Scrub typhus/tsutsugamushi fever | Asia, Northern Australia, serological evidence in sub-Saharan countries (Cameroon, Congo, Kenya) | |
| candidatus O. chuto | Scrub typhus/tsutsugamushi fever | Arabian Peninsula (Dubai), other areas in Middle East? | ||
| candidatus O. chiloensis | Scrub typhus/tsutsugamushi fever | Chile |
| Rickettsial | Localization | Function | Recognition by | ||
|---|---|---|---|---|---|
| Immunogens | B | CD4+ | CD8+ | ||
| Sca0 (OmpA) | OM | adhesion and invasion | √ | √ | |
| Sca1 | OM | adhesion and invasion | √ | ||
| Sca2 | OM | adhesion and invasion | √ | ||
| Sca3 | OM | adhesion and invasion | √ | ||
| Sca4 | C | binds and activates vinculin [107] | √ | ||
| Sca5 (OmpB) | OM | adhesion and invasion | √ | √ | √ |
| Adr1 | OM | adhesion and invasion, binds vitronectin, confers resistance to complement-mediated killing [108,109] | √ | ||
| Adr2 | OM | adhesion and invasion, binds vitronectin, confers resistance to complement-mediated killing [110] | √ | √ | √ |
| TolC | OM | adhesion and invasion of vascular endothelial cells [111] | √ | ||
| OmpW | OM | adhesion and invasion of vascular endothelial cells [111] | |||
| Porin-4 | IM/OM/EC | export of glycostructures (eg. LPS O-antigen) | √ | ||
| YbgF | OM/C | tol-pal system protein | √ | √ | √ |
| GroEL | C/OM } | 60 kDa heat shock protein, molecular chaperone; surface-exposed [105,106,112] | √ | ||
| PrsA | OM/C | Parvulin-like peptidyl-prolyl cis-trans isomerase (Parvulin-like PPIase), protein export protein | √ | ||
| RplY | C/OM | 50S ribosomal protein L25/general stress protein Ctc | √ | ||
| RpsB | C/OM | 30S ribosomal protein S2 | √ | ||
| SurA | C/OM | chaperone SurA, parvulin-like peptidyl-prolyl isomerase | √ | ||
| RP403 | C/OM | RecB family exonuclease | √ | ||
| RP598 | C/OM | transcription repair coupling factor | √ | ||
| RP739 | IM | ADP/ATP carrier protein (tlc5) | √ | ||
| RP778 | C/OM | DNA polymerase III a chain (dnaE) | √ | ||
| RP884 | C | ferrochelatase (hemE) | √ | ||
| Orientia immunogens | Localization | Function | B | CD4+ | CD8+ |
| Sta22 | OM | TSA47, transposase/DegP-like serin protease | √ | √ | |
| Sta47 | C/P | TSA56, multi-pass membrane protein | √ | ||
| Sta56 | OM | autotransporter protein | √ | √ | |
| ScaA | OM | autotransporter protein | √ | √ | √ |
| ScaC | OM | autotransporter protein | √ | ||
| ScaD | OM | autotransporter protein | √ | ||
| ScaE | C/OM | TSA47, transposase/DegP-like serin protease | √ | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Osterloh, A. Vaccine Design and Vaccination Strategies against Rickettsiae. Vaccines 2021, 9, 896. https://doi.org/10.3390/vaccines9080896
Osterloh A. Vaccine Design and Vaccination Strategies against Rickettsiae. Vaccines. 2021; 9(8):896. https://doi.org/10.3390/vaccines9080896
Chicago/Turabian StyleOsterloh, Anke. 2021. "Vaccine Design and Vaccination Strategies against Rickettsiae" Vaccines 9, no. 8: 896. https://doi.org/10.3390/vaccines9080896
APA StyleOsterloh, A. (2021). Vaccine Design and Vaccination Strategies against Rickettsiae. Vaccines, 9(8), 896. https://doi.org/10.3390/vaccines9080896