
What Constitutes Protective Immunity Following Yellow Fever Vaccination?


Abstract
1. Introduction
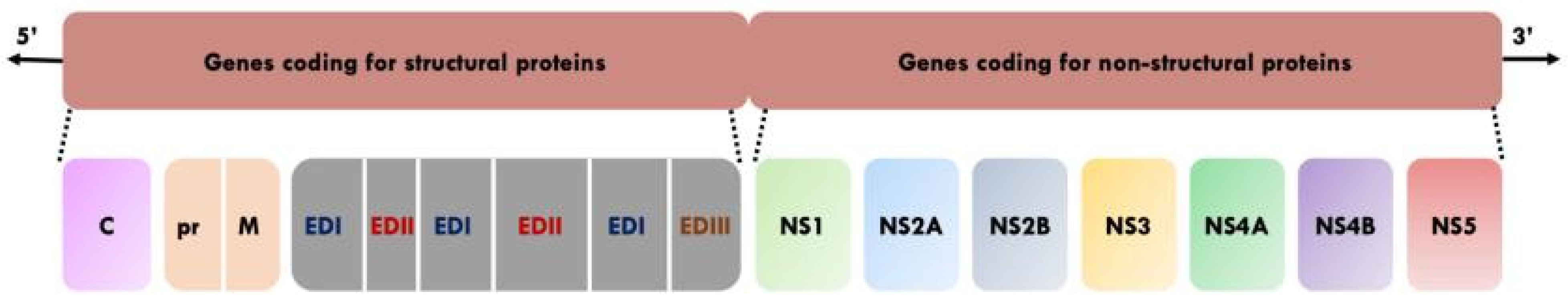
2. Molecular Biology of YF
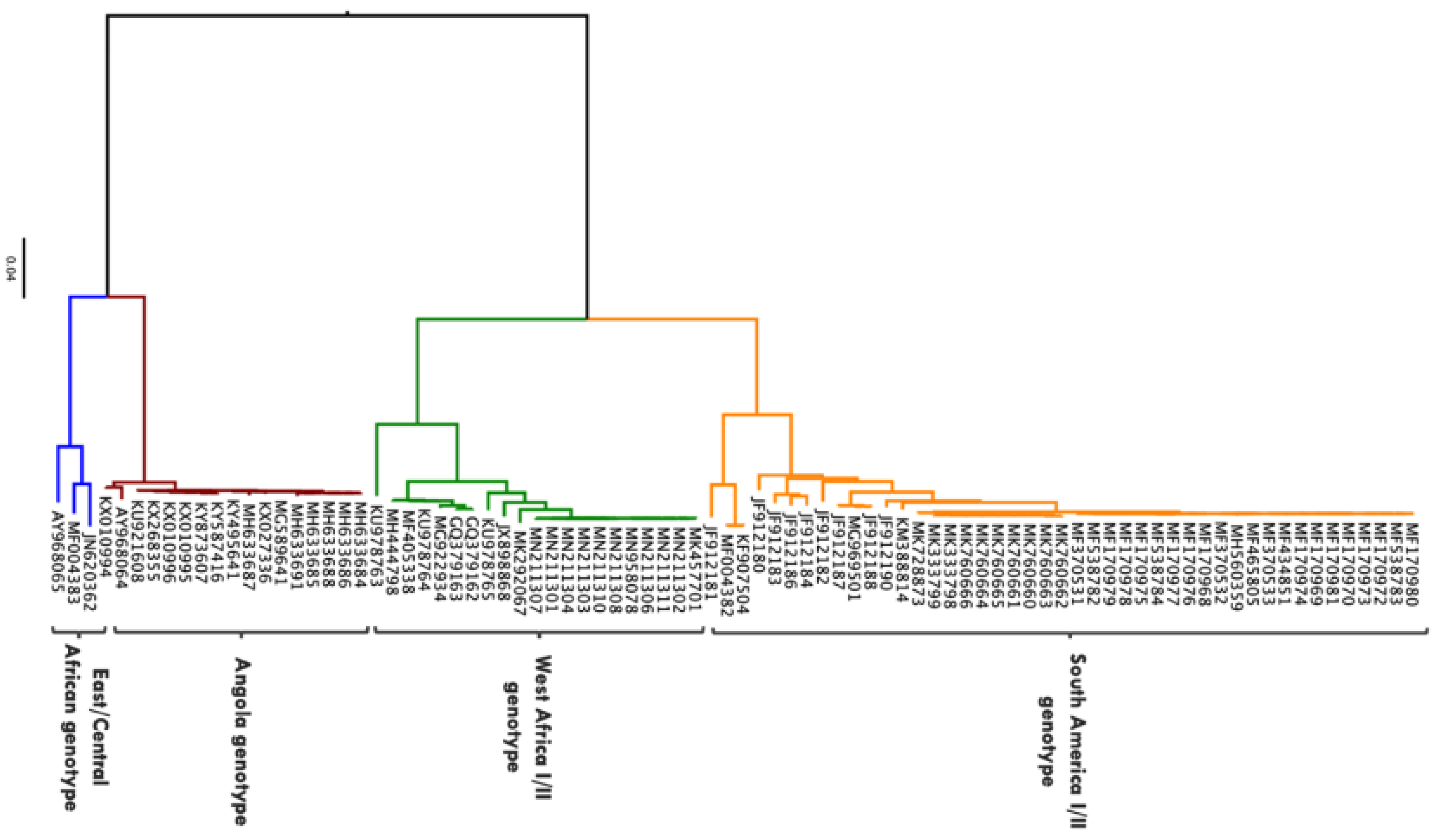
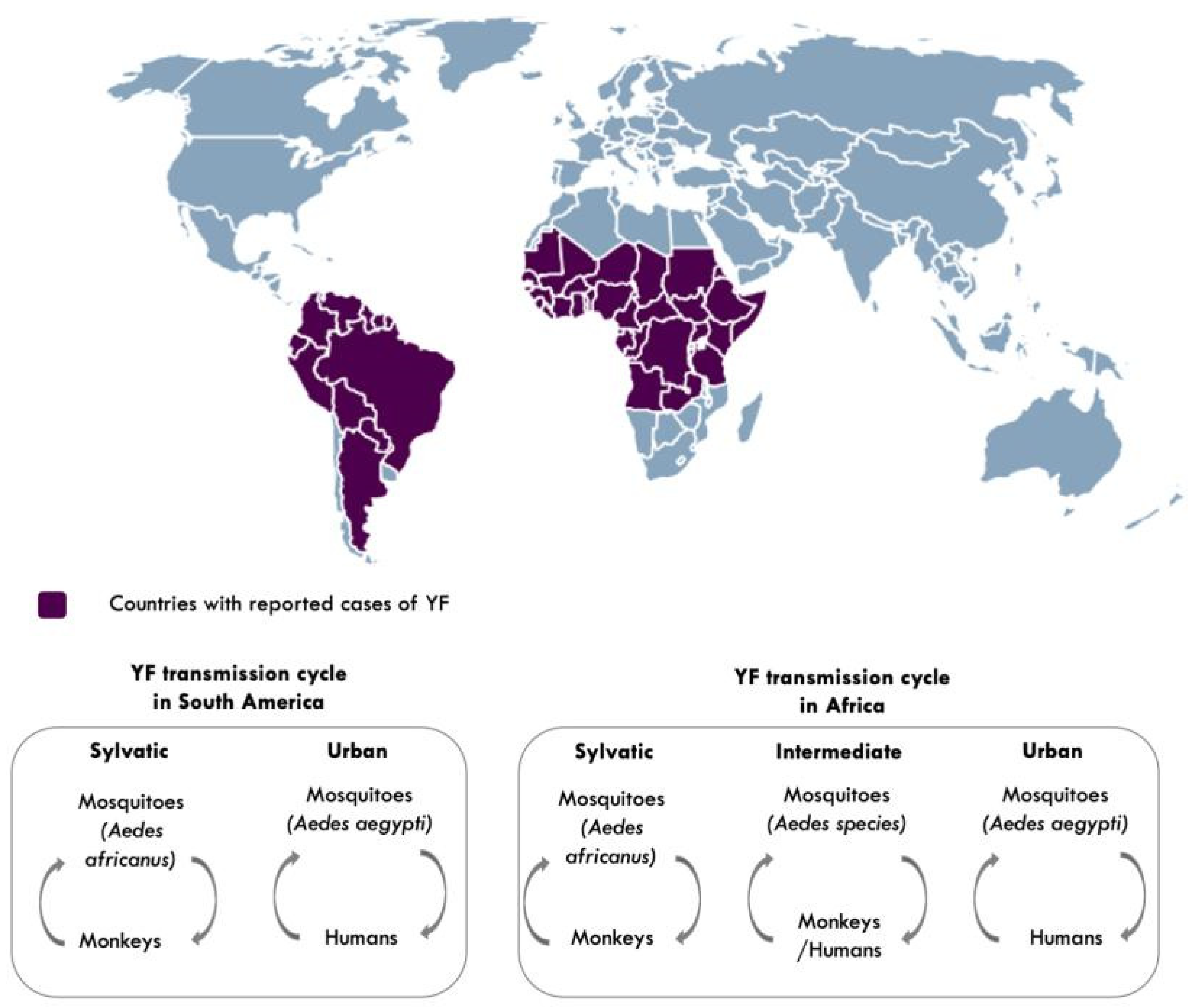
3. Diversity and Transmission of YFV
4. Clinical Presentation, Diagnosis and Treatment of YF
5. YF Prevention
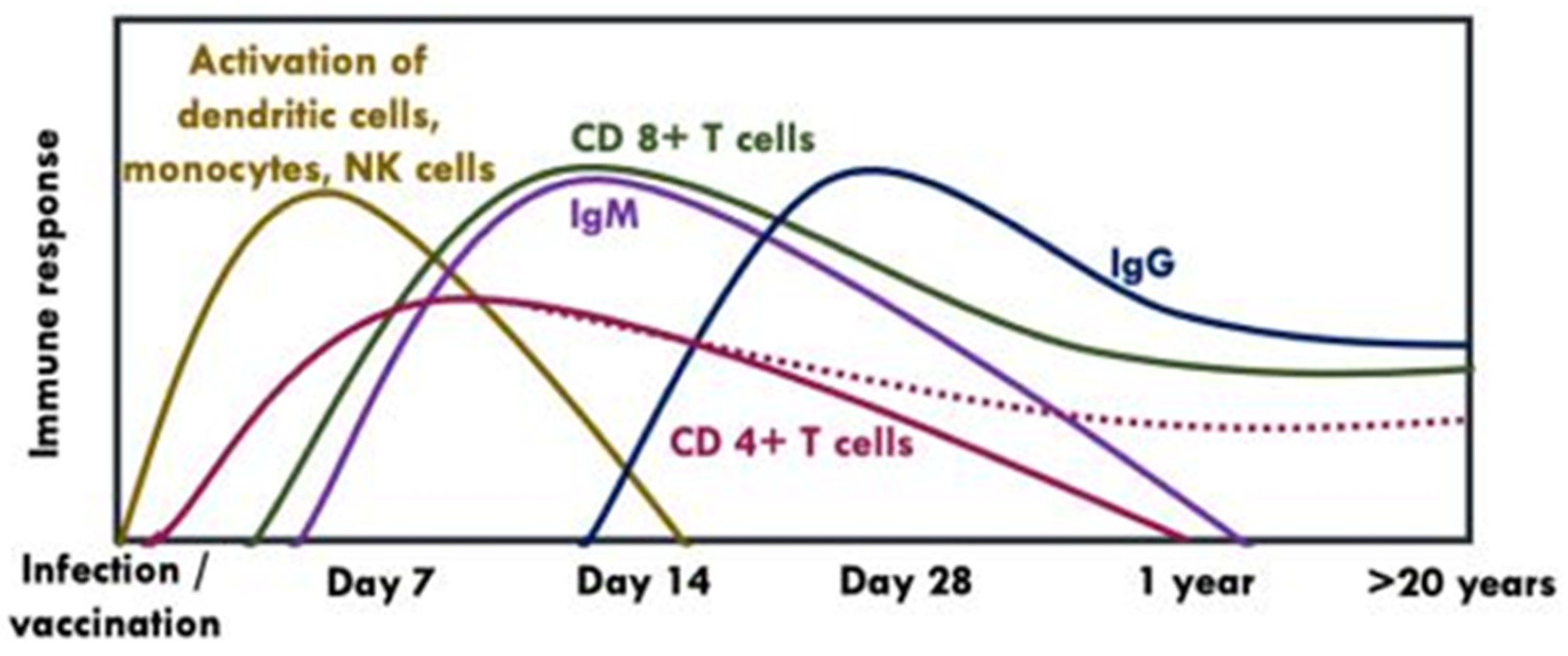
6. Quantity and Quality of YF Vaccine-Induced Immune Response
6.1. Cellular Immunity
6.2. Humoral Immunity
7. Research Gaps
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Staples, J.E.; Barrett, A.D.T.T.; Wilder-Smith, A.; Hombach, J. Review of data and knowledge gaps regarding yellow fever vaccine-induced immunity and duration of protection. NPJ Vaccines 2020, 5, 54. [Google Scholar] [CrossRef] [PubMed]
- Gardner, C.L.; Ryman, K.D. Yellow fever: A reemerging threat. Clin. Lab. Med. 2010, 30, 237–260. [Google Scholar] [CrossRef] [PubMed]
- World Health Organisation. Vaccines and vaccination against yellow fever: WHO Position Paper, June 2013—Recommendations. Vaccine 2015, 33, 76–77. [Google Scholar] [CrossRef]
- Chen, L.H.; Wilson, M.E. Yellow fever control: Current epidemiology and vaccination strategies. Trop. Dis. Travel Med. Vaccines 2020, 6, 1. [Google Scholar] [CrossRef]
- World Health Organisation. Eliminate Yellow Fever Epidemics (EYE) Strategy 2017–2026. Wkly. Epidemiol. Rec. 2017, 92, 193–204. Available online: https://www.who.int/initiatives/eye-strategy (accessed on 2 February 2021).
- Gould, E.; Solomon, T. Pathogenic flaviviruses. Lancet 2008, 371, 500–509. [Google Scholar] [CrossRef]
- Rice, C.M.; Lenches, E.M.; Eddy, S.R.; Shin, S.J.; Sheets, R.L.; Strauss, J.H. Nucleotide sequence of yellow fever virus: Implications for flavivirus gene expression and evolution. Science 1985, 229, 726–733. [Google Scholar] [CrossRef]
- Davis, E.H.; Barrett, A.D.T. Structure-Function of the Yellow Fever Virus Envelope Protein: Analysis of Antibody Epitopes. Viral Immunol. 2020, 33, 12–21. [Google Scholar] [CrossRef]
- Fernandez-Garcia, M.D.; Mazzon, M.; Jacobs, M.; Amara, A. Pathogenesis of Flavivirus Infections: Using and Abusing the Host Cell. Cell Host Microbe 2009, 5, 318–328. [Google Scholar] [CrossRef]
- Bressanelli, S.; Stiasny, K.; Allison, S.L.; Stura, E.A.; Duquerroy, S.; Lescar, J.; Heinz, F.X.; Rey, F.A. Structure of a flavivirus envelope glycoprotein in its low-pH-induced membrane fusion conformation. EMBO J. 2004, 23, 728–738. [Google Scholar] [CrossRef]
- Brinton, M.A. The molecular biology of West Nile virus: A new invader of the Western hemisphere. Annu. Rev. Microbiol. 2002, 56, 371–402. [Google Scholar] [CrossRef] [PubMed]
- Campos, J.L.S.; Mongkolsapaya, J.; Screaton, G.R. The immune response against flaviviruses. Nat. Immunol. 2018, 19, 1189–1198. [Google Scholar] [CrossRef] [PubMed]
- Vratskikh, O.; Stiasny, K.; Zlatkovic, J.; Tsouchnikas, G.; Jarmer, J.; Karrer, U.; Roggendorf, M.; Roggendorf, H.; Allwinn, R.; Heinz, F.X. Dissection of Antibody Specificities Induced by Yellow Fever Vaccination. PLoS Pathog. 2013, 9, e1003458. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.R.; Wang, X.; Saron, W.A.A.; Gan, E.S.; Tan, H.C.; Mok, D.Z.L.; Zhang, S.L.-X.; Lee, Y.H.; Liang, C.; Wijaya, L.; et al. Cross-reactive antibodies enhance live attenuated virus infection for increased immunogenicity. Nat. Microbiol. 2016, 1, 1–10. [Google Scholar] [CrossRef]
- Wec, A.Z.; Haslwanter, D.; Abdiche, Y.N.; Shehata, L.; Pedreno-Lopez, N.; Moyer, C.L.; Bornholdt, Z.A.; Lilov, A.; Nett, J.H.; Jangra, R.K.; et al. Longitudinal dynamics of the human B cell response to the yellow fever 17D vaccine. Proc. Natl. Acad. Sci. USA 2020, 117, 6675–6685. [Google Scholar] [CrossRef]
- Beasley, D.W.C.; McAuley, A.J.; Bente, D.A. Yellow fever virus: Genetic and phenotypic diversity and implications for detection, prevention and therapy. Antivir. Res. 2015, 115, 48–70. [Google Scholar] [CrossRef]
- Sall, A.A.; Faye, O.; Diallo, M.; Firth, C.; Kitchen, A.; Holmes, E.C. Yellow Fever Virus Exhibits Slower Evolutionary Dynamics than Dengue Virus. J. Virol. 2010, 84, 765–772. [Google Scholar] [CrossRef] [PubMed]
- Klitting, R.; Fischer, C.; Drexler, J.F.; Gould, E.A.; Roiz, D.; Paupy, C.; De Lamballerie, X. What does the future hold for yellow fever virus? (II). Genes 2018, 9, 425. [Google Scholar] [CrossRef] [PubMed]
- Nunes, M.R.T.; Palacios, G.; Cardoso, J.F.; Martins, L.C.; Sousa, E.C.; de Lima, C.P.S.; Medeiros, D.B.A.; Savji, N.; Desai, A.; Rodrigues, S.G.; et al. Genomic and Phylogenetic Characterization of Brazilian Yellow Fever Virus Strains. J. Virol. 2012, 86, 13263–13271. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Shearer, F.M.; Moyes, C.L.; Pigott, D.M.; Brady, O.J.; Marinho, F.; Deshpande, A.; Longbottom, J.; Browne, A.J.; Kraemer, M.U.G.; O’Reilly, K.M.; et al. Global yellow fever vaccination coverage from 1970 to 2016: An adjusted retrospective analysis. Lancet Infect Dis. 2017, 17, 1109–1126. [Google Scholar] [CrossRef]
- World Health Organisation. Yellow Fever Laboratory Diagnostic Testing in Africa Interim Guidance. 2016. Available online: https://apps.who.int/iris/bitstream/handle/10665/246226/WHO-OHE-YF-LAB-16.1-eng.pdf?sequence=1 (accessed on 25 May 2021).
- Barrett, A.D. Yellow fever vaccines. Biologicals 1997, 25, 17–25. [Google Scholar] [CrossRef]
- Fernandez-Garcia, M.D.; Meertens, L.; Chazal, M.; Hafirassou, M.L.; Dejarnac, O.; Zamborlini, A.; Despres, P.; Sauvonnet, N.; Arenzana-Seisdedos, F.; Jouvenet, N.; et al. Vaccine and Wild-Type Strains of Yellow Fever Virus Engage Distinct Entry Mechanisms and Differentially Stimulate Antiviral Immune Responses. MBio 2016, 7, e01956-15. [Google Scholar] [CrossRef] [PubMed]
- Casey, R.M.; Harris, J.B.; Ahuka-Mundeke, S.; Dixon, M.G.; Kizito, G.M.; Nsele, P.M.; Umutesi, G.; Laven, J.; Kosoy, O.; Paluku, G.; et al. Immunogenicity of Fractional-Dose Vaccine during a Yellow Fever Outbreak—Final Report. N. Engl. J. Med. 2019, 381, 444–454. [Google Scholar] [CrossRef]
- Roukens, A.H.; Vossen, A.C.; Bredenbeek, P.J.; van Dissel, J.T.; Visser, L.G. Intradermally Administered Yellow Fever Vaccine at Reduced Dose Induces a Protective Immune Response: A Randomized Controlled Non-Inferiority Trial. PLoS ONE 2008, 3, e1993. [Google Scholar] [CrossRef] [PubMed]
- Juan-Giner, A.; Kimathi, D.; Grantz, K.H.; Hamaluba, M.; Kazooba, P.; Njuguna, P.; Fall, G.; Dia, M.; Bob, N.S.; Monath, T.P.; et al. Immunogenicity and safety of fractional doses of yellow fever vaccines: A randomised, double-blind, non-inferiority trial. Lancet 2021, 397, 119–127. [Google Scholar] [CrossRef]
- PATH. Yellow Fever Vaccination: The Potential of Dose-Sparing to Increase Vaccine Supply and Availability. Available online: https://www.path.org/resources/yellow-fever-vaccination-the-potential-of-dose-sparing-to-increase-vaccine-supply-and-availability/ (accessed on 2 February 2021).
- Wieten, R.W.; Jonker, E.F.F.; Van Leeuwen, E.M.M.; Remmerswaal, E.; Berge, I.J.M.T.; De Visser, A.W.; Van Genderen, P.J.J.; Goorhuis, A.; Visser, L.G.; Grobusch, M.P.; et al. A Single 17D Yellow Fever Vaccination Provides Lifelong Immunity; Characterization of Yellow-Fever-Specific Neutralizing Antibody and T-Cell Responses after Vaccination. PLoS ONE 2016, 11, e0149871. [Google Scholar] [CrossRef] [PubMed]
- Campi-Azevedo, A.C.; Peruhype-Magalhāes, V.; Coelho-Dos-Reis, J.G.; Antonelli, L.R.; Costa-Pereira, C.; Speziali, E.; Reis, L.R.; Lemos, J.A.; Ribeiro, J.G.L.; Camacho, L.A.B.; et al. 17DD Yellow Fever Revaccination and Heightened Long-Term Immunity in Populations of Disease-Endemic Areas, Brazil. Emerg. Infect. Dis. 2019, 25, 1511–1521. [Google Scholar] [CrossRef]
- Domingo, C.; Fraissinet, J.; Ansah, P.O.; Kelly, C.; Bhat, N.; Sow, S.O.; Mejía, J.E. Long-term immunity against yellow fever in children vaccinated during infancy: A longitudinal cohort study. Lancet Infect. Dis. 2019, 19, 1363–1370. [Google Scholar] [CrossRef]
- de Noronha, T.G.; Maia, M.D.L.D.S.; Ribeiro, J.G.L.; Lemos, J.A.C.; de Lima, S.M.B.; Martins-Filho, O.A.; Campi-Azevedo, A.C.; Freire, M.D.S.; Martins, R.D.M.; Camacho, L.A.B. Duration of post-vaccination humoral immunity against yellow fever in children. Vaccine 2019, 37, 7147–7154. [Google Scholar] [CrossRef]
- Chowdhury, P.R.; Meier, C.; Laraway, H.; Tang, Y.; Hodgson, A.; Sow, S.O.; Enwere, G.C.; Plikaytis, B.D.; Kulkarni, P.S.; Preziosi, M.-P.; et al. Immunogenicity of Yellow Fever Vaccine Coadministered With MenAfriVac in Healthy Infants in Ghana and Mali. Clin. Infect. Dis. 2015, 61, S586–S593. [Google Scholar] [CrossRef] [PubMed]
- López, P.; Lanata, C.F.; Zambrano, B.; Cortés, M.; Andrade, T.; Amemiya, I.; Terrones, C.; Gil, A.I.; Verastegui, H.; Marquez, V.; et al. Immunogenicity and Safety of Yellow Fever Vaccine (Stamaril) When Administered Concomitantly With a Tetravalent Dengue Vaccine Candidate in Healthy Toddlers at 12–13 Months of Age in Colombia and Peru. Pediatr. Infect. Dis. J. 2016, 35, 1140–1147. [Google Scholar] [CrossRef] [PubMed]
- Pulendran, B. Learning immunology from the yellow fever vaccine: Innate immunity to systems vaccinology. Nat. Rev. Immunol. 2009, 9, 741–747. [Google Scholar] [CrossRef] [PubMed]
- Watson, A.M.; Klimstra, W.B.T. Cell-Mediated Immunity towards Yellow Fever Virus and Useful Animal Models. Viruses 2017, 9, 77. [Google Scholar] [CrossRef]
- Salerno-Gonçalves, R.; Sztein, M.B. Cell-mediated immunity and the challenges for vaccine development. Trends Microbiol. 2006, 14, 536–542. [Google Scholar] [CrossRef]
- Bovay, A.; Marraco, S.A.F.; Speiser, D.E. Yellow fever virus vaccination: An emblematic model to elucidate robust human immune responses. Hum. Vaccines Immunother. 2021, 1–11. [Google Scholar] [CrossRef]
- James, E.A.; LaFond, R.E.; Gates, T.J.; Mai, D.T.; Malhotra, U.; Kwok, W.W. Yellow Fever Vaccination Elicits Broad Functional CD4 + T Cell Responses That Recognize Structural and Nonstructural Proteins. J. Virol. 2013, 87, 12794–12804. [Google Scholar] [CrossRef]
- Akondy, R.; Monson, N.D.; Miller, J.D.; Edupuganti, S.; Teuwen, D.; Wu, H.; Quyyumi, F.; Garg, S.; Altman, J.D.; Del Rio, C.; et al. The Yellow Fever Virus Vaccine Induces a Broad and Polyfunctional Human Memory CD8+ T Cell Response. J. Immunol. 2009, 183, 7919–7930. [Google Scholar] [CrossRef]
- Martins, M.A.; Silva, M.L.; Marciano, A.P.V.; Peruhype-Magalhães, V.; Eloi-Santos, S.M.; Ribeiro, J.G.L.; Correa-Oliveira, R.; Homma, A.; Kroon, E.G.; Teixeira-Carvalho, A.; et al. Activation/modulation of adaptive immunity emerges simultaneously after 17DD yellow fever first-time vaccination: Is this the key to prevent severe adverse reactions following immunization? Clin. Exp. Immunol. 2007, 148, 90–100. [Google Scholar] [CrossRef]
- Bovay, A.; Nassiri, S.; Hajjami, H.M.; Mondéjar, P.M.; Akondy, R.S.; Ahmed, R.; Lawson, B.; Speiser, D.E.; Marraco, S.A.F. Minimal immune response to booster vaccination against Yellow Fever associated with pre-existing antibodies. Vaccine 2020, 38, 2172–2182. [Google Scholar] [CrossRef]
- Blom, K.; Braun, M.; Ivarsson, M.A.; Gonzalez, V.D.; Falconer, K.; Moll, M.; Ljunggren, H.-G.; Michaëlsson, J.; Sandberg, J.K. Temporal Dynamics of the Primary Human T Cell Response to Yellow Fever Virus 17D As It Matures from an Effector- to a Memory-Type Response. J. Immunol. 2013, 190, 2150–2158. [Google Scholar] [CrossRef] [PubMed]
- Marraco, S.A.F.; Soneson, C.; Cagnon, L.; Gannon, P.O.; Allard, M.; Maillard, S.A.; Montandon, N.; Rufer, N.; Waldvogel, S.; Delorenzi, M.; et al. Long-lasting stem cell–like memory CD8+ T cells with a naïve-like profile upon yellow fever vaccination. Sci. Transl. Med. 2015, 7, 282ra48. [Google Scholar] [CrossRef] [PubMed]
- Da Costa-Rocha, I.A.; Campi-Azevedo, A.C.; Peruhype-Magalhães, V.; Coelho-Dos-Reis, J.G.; Fradico, J.R.B.; Souza-Lopes, T.; Reis, L.R.; Freire, L.C.; Costa-Pereira, C.; Mambrini, J.V.D.M.; et al. Duration of Humoral and Cellular Immunity 8 Years After Administration of Reduced Doses of the 17DD-Yellow Fever Vaccine. Front. Immunol. 2019, 10, 1211. [Google Scholar] [CrossRef] [PubMed]
- Campi-Azevedo, A.C.; Reis, L.R.; Peruhype-Magalhães, V.; Coelho-Dos-Reis, J.G.; Antonelli, L.R.; Fonseca, C.T.; Costa-Pereira, C.; Souza-Fagundes, E.M.; Da Costa-Rocha, I.A.; Mambrini, J.V.D.M.; et al. Short-Lived Immunity After 17DD Yellow Fever Single Dose Indicates That Booster Vaccination May Be Required to Guarantee Protective Immunity in Children. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef]
- Gibney, K.B.; Kosoy, O.I.; Fischer, M.; Edupuganti, S.; Lanciotti, R.S.; DeLorey, M.J.; Staples, J.E.; Panella, A.J.; Mulligan, M.J. Detection of Anti-Yellow Fever Virus Immunoglobulin M Antibodies at 3–4 Years Following Yellow Fever Vaccination. Am. J. Trop. Med. Hyg. 2012, 87, 1112–1115. [Google Scholar] [CrossRef]
- Collaborative Group for Studies on Yellow Fever Vaccines. Duration of post-vaccination immunity against yellow fever in adults. Vaccine 2014, 32, 4977–4984. [Google Scholar] [CrossRef]
- Miyaji, K.T.; Avelino-Silva, V.I.; Simões, M.; Freire, M.D.S.; De Medeiros, C.R.; Braga, P.E.; Neves, M.A.A.; Lopes, M.H.; Kallas, E.G.; Sartori, A.M.C. Prevalence and titers of yellow fever virus neutralizing antibodies in previously vaccinated adults. Rev. Inst. Med. Trop. S. Paulo 2017, 59. [Google Scholar] [CrossRef]
- Martins, R.D.M.; Maia, M.D.L.S.; De Lima, S.M.B.; De Noronha, T.G.; Xavier, J.R.; Camacho, L.A.B.; De Albuquerque, E.M.; Farias, R.H.G.; Castro, T.D.M.D.; Homma, A. Duration of post-vaccination immunity to yellow fever in volunteers eight years after a dose-response study. Vaccine 2018, 36, 4112–4117. [Google Scholar] [CrossRef]
- Lindsey, N.P.; Horiuchi, K.A.; Fulton, D.C.; Panella, A.J.; Kosoy, O.I.; Velez, J.O.; Krow-Lucal, E.R.; Fischer, M.; Staples, J.E. Persistence of yellow fever virus-specific neutralizing antibodies after vaccination among US travellers. J. Travel Med. 2018, 25, 1–6. [Google Scholar] [CrossRef]
- Roukens, A.H.; Van Halem, K.; De Visser, A.W.; Visser, L.G. Long-Term Protection After Fractional-Dose Yellow Fever Vaccination: Follow-up Study of a Randomized, Controlled, Noninferiority Trial. Ann.Intern. Med. 2018, 169, 761–765. [Google Scholar] [CrossRef]
- Idoko, O.T.; Mohammed, N.; Ansah, P.; Hodgson, A.; Tapia, M.D.; Sow, S.O.; Chowdhury, P.R.; Niedrig, M.; Saathoff, E.; Kampmann, B. Antibody responses to yellow fever vaccine in 9 to 11-month-old Malian and Ghanaian children. Expert Rev. Vaccines 2019, 18, 867–875. [Google Scholar] [CrossRef]
- Nnaji, C.A.; Shey, M.S.; Adetokunboh, O.O.; Wiysonge, C.S. Immunogenicity and safety of fractional dose yellow fever vaccination: A systematic review and meta-analysis. Vaccine 2020, 38, 1291–1301. [Google Scholar] [CrossRef]
- Chung, A.W.; Alter, G. Systems serology: Profiling vaccine induced humoral immunity against HIV. Retrovirology 2017, 14, 57. [Google Scholar] [CrossRef] [PubMed]
- Vanderven, H.A.; Jegaskanda, S.; Wines, B.D.; Hogarth, P.M.; Carmuglia, S.; Rockman, S.; Chung, A.W.; Kent, S.J. Antibody-Dependent Cellular Cytotoxicity Responses to Seasonal Influenza Vaccination in Older Adults. J. Infect. Dis. 2018, 217, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Erdman, D.D.; Heath, J.L.; Watson, J.C.; Markowitz, L.E.; Bellini, W.J. Immunoglobulin M antibody response to measles virus following primary and secondary vaccination and natural virus infection. J. Med. Virol. 1993, 41, 44–48. [Google Scholar] [CrossRef] [PubMed]
- Arvin, A.M.; Fink, K.; Schmid, M.A.; Cathcart, A.; Spreafico, R.; Havenar-Daughton, C.; Lanzavecchia, A.; Corti, D.; Virgin, H.W. A perspective on potential antibody-dependent enhancement of SARS-CoV-2. Nat. Cell Biol. 2020, 584, 353–363. [Google Scholar] [CrossRef]
- Mishra, N.; Boudewijns, R.; Schmid, M.; Marques, R.; Sharma, S.; Neyts, J.; Dallmeier, K. Achimeric Japanese encephalitis vaccine protects against lethal yellow fever virus infection without inducing neutralizing antibodies. mBio 2020, 11. [Google Scholar] [CrossRef]
- Jennewein, M.F.; Alter, G. The Immunoregulatory Roles of Antibody Glycosylation. Trends Immunol. 2017, 38, 358–372. [Google Scholar] [CrossRef] [PubMed]
- Irvine, E.B.; Alter, G. Understanding the role of antibody glycosylation through the lens of severe viral and bacterial diseases. Glycobiology 2020, 30, 241–253. [Google Scholar] [CrossRef]
- Karsten, C.; Mehta, N.; Shin, S.; Diefenbach, T.J.; Slein, M.D.; Karpinski, W.; Irvine, E.B.; Broge, T.; Suscovich, T.J.; Alter, G. A versatile high-throughput assay to characterize antibody-mediated neutrophil phagocytosis. J. Immunol. Methods 2019, 471, 46–56. [Google Scholar] [CrossRef]
- Tay, M.Z.; Wiehe, K.; Pollara, J. Antibody-Dependent Cellular Phagocytosis in Antiviral Immune Responses. Front. Immunol. 2019, 10, 332. [Google Scholar] [CrossRef]
- Fischinger, S.; Fallon, J.K.; Michell, A.; Broge, T.; Suscovich, T.J.; Streeck, H.; Alter, G. A high-throughput, bead-based, antigen-specific assay to assess the ability of antibodies to induce complement activation. J. Immunol. Methods 2019, 473, 112630. [Google Scholar] [CrossRef] [PubMed]
- Román, V.R.G.; Murray, J.C.; Weiner, L.M. Antibody-Dependent Cellular Cytotoxicity (ADCC). In Antibody Fc: Linking Adaptive and Innate Immunity; Elsevier Inc. Academic Press: Cambridge, MA, USA, 2013; pp. 1–27. [Google Scholar]
- Bifani, A.M.; Ong, E.Z.; De Alwis, R. Vaccination and Therapeutics: Responding to the Changing Epidemiology of Yellow Fever. Curr. Treat. Options Infect. Dis. 2020, 12, 398–409. [Google Scholar] [CrossRef] [PubMed]
- Muyanja, E.; Ssemaganda, A.; Ngauv, P.; Cubas, R.; Perrin, H.; Srinivasan, D.; Canderan, G.; Lawson, B.; Kopycinski, J.; Graham, A.S.; et al. Immune activation alters cellular and humoral responses to yellow fever 17D vaccine. J. Clin. Investig. 2014, 124, 3147–3158. [Google Scholar] [CrossRef]
- Selva, K.J.; van de Sandt, C.E.; Lemke, M.M.; Lee, C.Y.; Shoffner, S.K.; Chua, B.Y.; Davis, S.K.; Nguyen, T.H.O.; Rowntree, L.C.; Hensen, L.; et al. Systems serology detects functionally distinct coronavirus antibody features in children and elderly. Nat. Commun. 2021, 12, 1–14. [Google Scholar] [CrossRef]
- Suscovich, T.J.; Fallon, J.K.; Das, J.; Demas, A.R.; Crain, J.; Linde, C.H.; Michell, A.; Natarajan, H.; Arevalo, C.; Broge, T.; et al. Mapping functional humoral correlates of protection against malaria challenge following RTS,S/AS01 vaccination. Sci. Transl. Med. 2020, 12, eabb4757. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody Effector Function | Antibody Dependent Cellular Phagocytosis (ADCP) | Antibody Dependent Cellular Cytotoxicity (ADCC) | Antibody Dependent Complement Deposition (ADCD) |
|---|---|---|---|
| Description | Following activation of Fc receptors, effector cells eliminate antibody-opsonised pathogens through phagocytosis and also activate adaptive immune responses by facilitating antigen presentation and/or secretion of inflammatory mediators | Following activation of Fc receptors, effector cells recognise and kill antibody-coated target cells through perforin/granzyme cell death pathway, FAS-L pathway and/or reactive oxygen species pathway | Antibody-antigen complexes activate complement proteins which, following a cascade of enzymatic reaction, result in the assembly of membrane attack complexes and the formation of pores on the surface of target cells or pathogens causing cell-lysis |
| Antibodies/Fc receptors commonly implicated in specific antibody effector functions | IgA (FcαRI); IgG-dependent (FcγR1, FcγRII, FcγRIIIa) | IgG-dependent (FcγR1, FcγRII, FcγRIIIa) | IgM, IgG1, IgG2, IgG3, IgG4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mokaya, J.; Kimathi, D.; Lambe, T.; Warimwe, G.M. What Constitutes Protective Immunity Following Yellow Fever Vaccination? Vaccines 2021, 9, 671. https://doi.org/10.3390/vaccines9060671
Mokaya J, Kimathi D, Lambe T, Warimwe GM. What Constitutes Protective Immunity Following Yellow Fever Vaccination? Vaccines. 2021; 9(6):671. https://doi.org/10.3390/vaccines9060671
Chicago/Turabian StyleMokaya, Jolynne, Derick Kimathi, Teresa Lambe, and George M. Warimwe. 2021. "What Constitutes Protective Immunity Following Yellow Fever Vaccination?" Vaccines 9, no. 6: 671. https://doi.org/10.3390/vaccines9060671
APA StyleMokaya, J., Kimathi, D., Lambe, T., & Warimwe, G. M. (2021). What Constitutes Protective Immunity Following Yellow Fever Vaccination? Vaccines, 9(6), 671. https://doi.org/10.3390/vaccines9060671