
Flavivirus NS1 and Its Potential in Vaccine Development


Abstract
1. Introduction
2. Flavivirus Genome
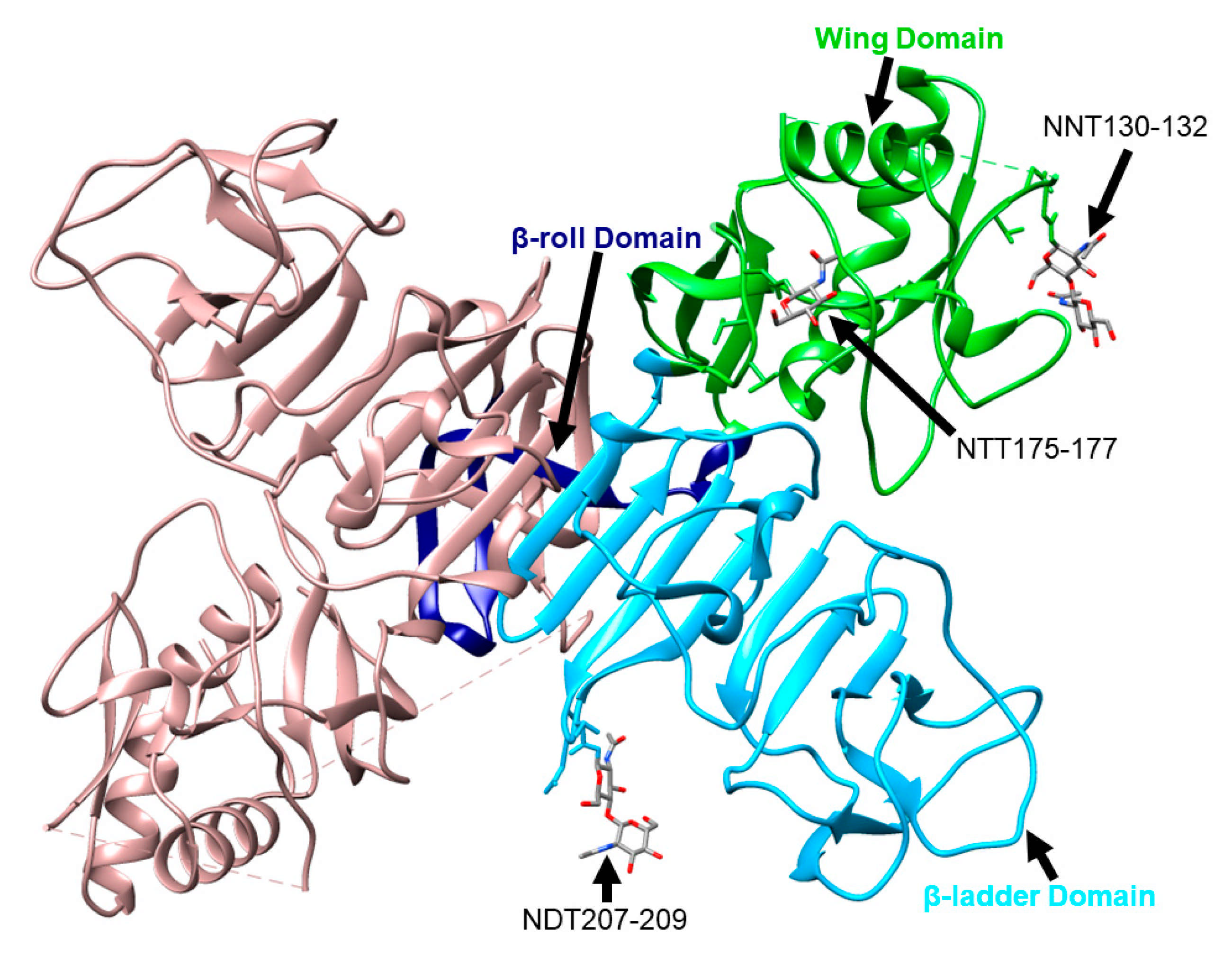
3. NS1 Structures
4. Cofactor in Viral Replication
5. Role of NS1 in Dengue Pathogenesis
6. Role of NS1 in Pathogenesis of Other Flaviviruses
7. Binding on Cell Surface
8. Anti-NS1 Antibodies and B Cell Epitopes
9. Complement and ADCC
10. Serology Using Complement Fixation and Neutralization Assays
11. Importance of Adding Carbohydrates
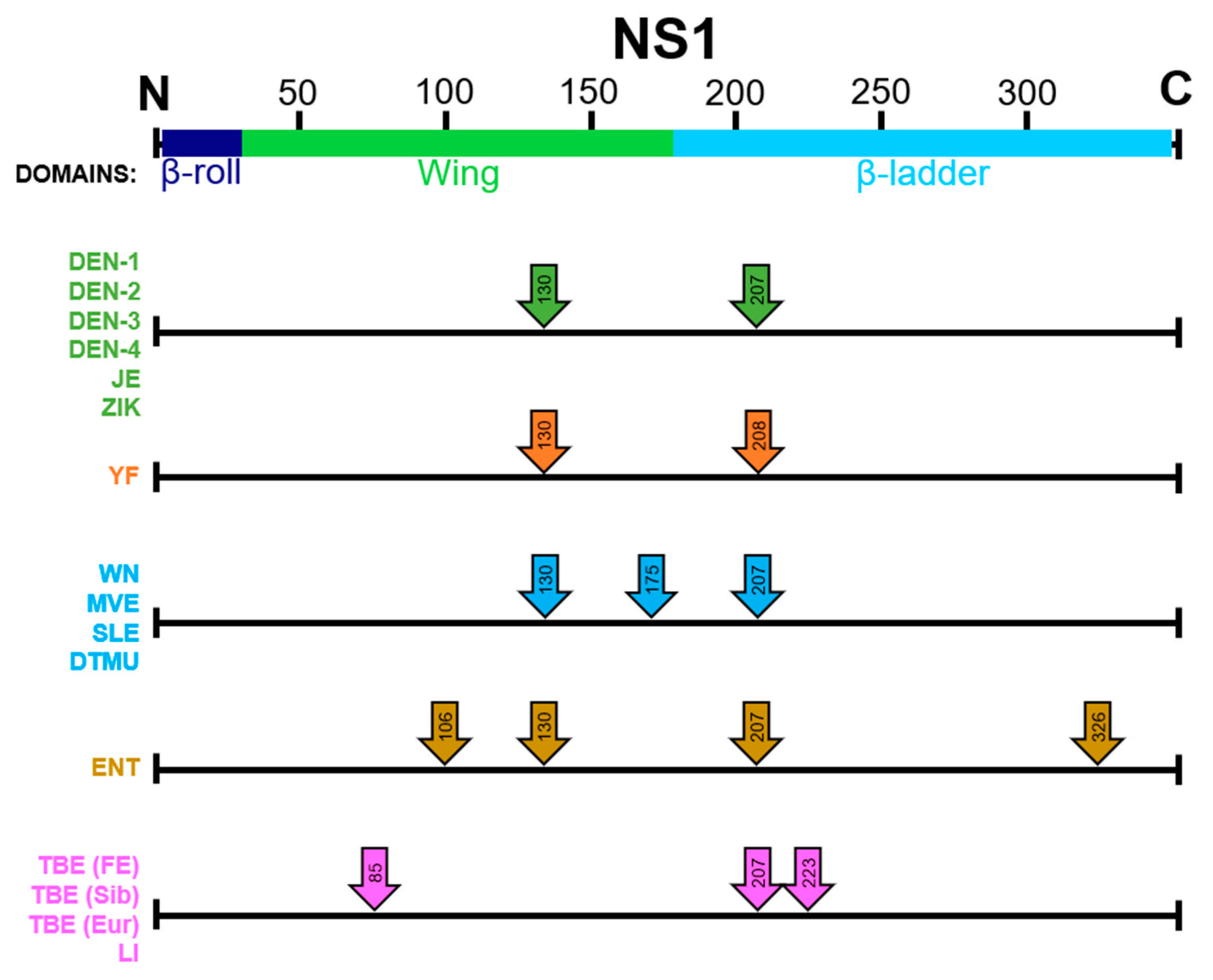
12. Effects of Removing Glycosylation Sites
13. NS1 as a Vaccine Immunogen
13.1. Subunit Vaccine Candidates
13.2. Vectored Vaccine Candidates
13.3. Live Attenuated Vaccines
13.4. Trans Complementation as a Vaccine Strategy
14. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kuno, G.; Chang, G.J.; Tsuchiya, K.R.; Karabatsos, N.; Cropp, C.B. Phylogeny of the Genus Flavivirus. J. Virol. 1998, 72, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Gubler, D.J. Dengue and Dengue Hemorrhagic Fever. Clin. Microbiol. Rev. 1998, 11, 480–496. [Google Scholar] [CrossRef] [PubMed]
- Barrett, A.D.T. Yellow Fever Live Attenuated Vaccine: A Very Successful Live Attenuated Vaccine but Still We Have Problems Controlling the Disease. Vaccine 2017, 35, 5951–5955. [Google Scholar] [CrossRef] [PubMed]
- The Lancet Infectious Diseases. The Dengue Vaccine Dilemma. Lancet Infect. Dis. 2018, 18, 123. [Google Scholar] [CrossRef]
- Li, X.; Ma, S.-J.; Liu, X.; Jiang, L.-N.; Zhou, J.-H.; Xiong, Y.-Q.; Ding, H.; Chen, Q. Immunogenicity and Safety of Currently Available Japanese Encephalitis Vaccines: A Systematic Review. Hum. Vaccines Immunother. 2015, 10, 3579–3593. [Google Scholar] [CrossRef]
- Lindenbach, B.D.; Rice, C.M. Molecular biology of flaviviruses. In Advances in Virus Research; Academic Press: Cambridge, MA, USA, 2003; Volume 59, pp. 23–61. [Google Scholar] [CrossRef]
- Muylaert, I.R.; Chambers, T.J.; Galler, R.; Rice, C.M. Mutagenesis of the N-Linked Glycosylation Sites of the Yellow Fever Virus NS1 Protein: Effects on Virus Replication and Mouse Neurovirulence. Virology 1996, 222, 159–168. [Google Scholar] [CrossRef]
- Whiteman, M.C.; Li, L.; Wicker, J.A.; Kinney, R.M.; Huang, C.; Beasley, D.W.C.; Chung, K.M.; Diamond, M.S.; Solomon, T.; Barrett, A.D.T. Development and Characterization of Non-Glycosylated E and NS1 Mutant Viruses as a Potential Candidate Vaccine for West Nile Virus. Vaccine 2010, 28, 1075–1083. [Google Scholar] [CrossRef]
- Kuno, G.; Chang, G.-J.J. Characterization of Sepik and Entebbe Bat Viruses Closely Related to Yellow Fever Virus. Am. J. Trop. Med. Hyg. 2006, 75, 1165–1170. [Google Scholar] [CrossRef]
- Mandl, C.W.; Heinz, F.X.; Stockl, E.; Kunz, C. Genome Sequence of Tick-Borne Encephalitis Virus (Western Subtype) and Comparative Analysis of Nonstructural Proteins with Other Flaviviruses. Virology 1989, 173, 291–301. [Google Scholar] [CrossRef]
- Blitvich, B.J.; Scanlon, D.; Shiell, B.J.; Mackenzie, J.S.; Pham, K.; Hall, R.A. Determination of the Intramolecular Disulfide Bond Arrangement and Biochemical Identification of the Glycosylation Sites of the Nonstructural Protein NS1 of Murray Valley Encephalitis Virus. J. Gen. Virol. 2001, 82, 2251–2256. [Google Scholar] [CrossRef]
- Crooks, A.J.; Lee, J.M.; Easterbrook, L.M.; Timofeev, A.V.; Stephenson, J.R. The NS1 Protein of Tick-Borne Encephalitis Virus Forms Multimeric Species upon Secretion from the Host Cell. J. Gen. Virol. 1994, 75, 3453–3460. [Google Scholar] [CrossRef]
- Pryor, M.J.; Wright, P.J. The Effects of Site-Directed Mutagenesis on the Dimerization and Secretion of the NS1 Protein Specified by Dengue Virus. Virology 1993, 194, 769–780. [Google Scholar] [CrossRef]
- Lai, Y.-C.; Chuang, Y.-C.; Liu, C.-C.; Ho, T.-S.; Lin, Y.-S.; Anderson, R.; Yeh, T.-M. Antibodies Against Modified NS1 Wing Domain Peptide Protect Against Dengue Virus Infection. Sci. Rep. 2017, 7, 6975. [Google Scholar] [CrossRef]
- Akey, D.L.; Brown, W.C.; Dutta, S.; Konwerski, J.; Jose, J.; Jurkiw, T.J.; DelProposto, J.; Ogata, C.M.; Skiniotis, G.; Kuhn, R.J.; et al. Flavivirus NS1 Crystal Structures Reveal a Surface for Membrane Association and Regions of Interaction with the Immune System. Science 2014, 343, 881–885. [Google Scholar] [CrossRef]
- Hertz, T.; Beatty, P.R.; MacMillen, Z.; Killingbeck, S.S.; Wang, C.; Harris, E. Antibody Epitopes Identified in Critical Regions of Dengue Virus Nonstructural 1 Protein in Mouse Vaccination and Natural Human Infections. J. Immunol. 2017, 198, 4025–4035. [Google Scholar] [CrossRef]
- Winkler, G.; Randolph, V.B.; Cleaves, G.R.; Ryan, T.E.; Stollar, V. Evidence That the Mature Form of the Flavivirus Nonstructural Protein NS1 Is a Dimer. Virology 1988, 162, 187–196. [Google Scholar] [CrossRef]
- Flamand, M.; Megret, F.; Mathieu, M.; Lepault, J.; Rey, F.A.; Deubel, V. Dengue Virus Type 1 Nonstructural Glycoprotein NS1 Is Secreted from Mammalian Cells as a Soluble Hexamer in a Glycosylation-Dependent Fashion. J. Virol. 1999, 73, 6104–6110. [Google Scholar] [CrossRef]
- Gutsche, I.; Coulibaly, F.; Voss, J.E.; Salmon, J.; d’Alayer, J.; Ermonval, M.; Larquet, E.; Charneau, P.; Krey, T.; Mégret, F.; et al. Secreted Dengue Virus Nonstructural Protein NS1 Is an Atypical Barrel-Shaped High-Density Lipoprotein. Proc. Natl. Acad. Sci. USA 2011, 108, 8003–8008. [Google Scholar] [CrossRef]
- Mackenzie, J.M.; Jones, M.K.; Young, P.R. Immunolocalization of the Dengue Virus Nonstructural Glycoprotein NS1 Suggests a Role in Viral RNA Replication. Virology 1996, 220, 232–240. [Google Scholar] [CrossRef]
- Lindenbach, B.D.; Rice, C.M. Genetic Interaction of Flavivirus Nonstructural Proteins NS1 and NS4A as a Determinant of Replicase Function. J. Virol. 1999, 73, 4611–4621. [Google Scholar] [CrossRef]
- Tan, M.J.A.; Brown, N.G.; Chan, K.W.K.; Jin, J.Y.; Zu Kong, S.Y.; Vasudevan, S.G. Mutations in the Cytoplasmic Domain of Dengue Virus NS4A Affect Virus Fitness and Interactions with Other Non-Structural Proteins. J. Gen. Virol. 2020. [Google Scholar] [CrossRef]
- Youn, S.; Li, T.; McCune, B.T.; Edeling, M.A.; Fremont, D.H.; Cristea, I.M.; Diamond, M.S. Evidence for a Genetic and Physical Interaction between Nonstructural Proteins NS1 and NS4B That Modulates Replication of West Nile Virus. J. Virol. 2012, 86, 7360–7371. [Google Scholar] [CrossRef]
- Fan, J.; Liu, Y.; Yuan, Z. Critical Role of Dengue Virus NS1 Protein in Viral Replication. Virol. Sin. 2014, 29, 162–169. [Google Scholar] [CrossRef]
- Wallis, T.P.; Huang, C.-Y.; Nimkar, S.B.; Young, P.R.; Gorman, J.J. Determination of the Disulfide Bond Arrangement of Dengue Virus NS1 Protein. J. Biol. Chem. 2004, 279, 20729–20741. [Google Scholar] [CrossRef]
- Hall, R.A.; Khromykh, A.A.; Mackenzie, J.M.; Scherret, J.H.; Khromykh, T.I.; Mackenzie, J.S. Loss of Dimerisation of the Nonstructural Protein NS1 of Kunjin Virus Delays Viral Replication and Reduces Virulence in Mice, but Still Allows Secretion of NS1. Virology 1999, 264, 66–75. [Google Scholar] [CrossRef]
- Clark, D.C.; Lobigs, M.; Lee, E.; Howard, M.J.; Clark, K.; Blitvich, B.J.; Hall, R.A. In Situ Reactions of Monoclonal Antibodies with a Viable Mutant of Murray Valley Encephalitis Virus Reveal an Absence of Dimeric NS1 Protein. J. Gen. Virol. 2007, 88, 1175–1183. [Google Scholar] [CrossRef]
- Khromykh, A.A.; Sedlak, P.L.; Westaway, E.G. Cis- and Trans-Acting Elements in Flavivirus RNA Replication. J. Virol. 2000, 74, 3253–3263. [Google Scholar] [CrossRef]
- Wang, C.; Puerta-Guardo, H.; Biering, S.B.; Glasner, D.R.; Tran, E.B.; Patana, M.; Gomberg, T.A.; Malvar, C.; Lo, N.T.N.; Espinosa, D.A.; et al. Endocytosis of Flavivirus NS1 Is Required for NS1-Mediated Endothelial Hyperpermeability and Is Abolished by a Single N-Glycosylation Site Mutation. PLoS Pathog. 2019, 15, e1007938. [Google Scholar] [CrossRef]
- Lin, C.-F.; Lei, H.-Y.; Shiau, A.-L.; Liu, C.-C.; Liu, H.-S.; Yeh, T.-M.; Chen, S.-H.; Lin, Y.-S. Antibodies from Dengue Patient Sera Cross-React with Endothelial Cells and Induce Damage. J. Med. Virol. 2003, 69, 82–90. [Google Scholar] [CrossRef]
- Chen, H.-R.; Chuang, Y.-C.; Lin, Y.-S.; Liu, H.-S.; Liu, C.-C.; Perng, G.-C.; Yeh, T.-M. Dengue Virus Nonstructural Protein 1 Induces Vascular Leakage through Macrophage Migration Inhibitory Factor and Autophagy. PLoS Negl. Trop. Dis. 2016, 10, e0004828. [Google Scholar] [CrossRef]
- Soe, H.J.; Manikam, R.; Raju, C.S.; Khan, M.A.; Sekaran, S.D. Correlation of Host Inflammatory Cytokines and Immune-Related Metabolites, but Not Viral NS1 Protein, with Disease Severity of Dengue Virus Infection. PLoS ONE 2020, 15, e0237141. [Google Scholar] [CrossRef]
- Puerta-Guardo, H.; Glasner, D.R.; Espinosa, D.A.; Biering, S.B.; Patana, M.; Ratnasiri, K.; Wang, C.; Beatty, P.R.; Harris, E. Flavivirus NS1 Triggers Tissue-Specific Vascular Endothelial Dysfunction Reflecting Disease Tropism. Cell Rep. 2019, 26, 1598–1613.e8. [Google Scholar] [CrossRef] [PubMed]
- Chao, C.-H.; Wu, W.-C.; Lai, Y.-C.; Tsai, P.-J.; Perng, G.-C.; Lin, Y.-S.; Yeh, T.-M. Dengue Virus Nonstructural Protein 1 Activates Platelets via Toll-like Receptor 4, Leading to Thrombocytopenia and Hemorrhage. PLOS Pathog. 2019, 15, e1007625. [Google Scholar] [CrossRef] [PubMed]
- Modhiran, N.; Watterson, D.; Blumenthal, A.; Baxter, A.G.; Young, P.R.; Stacey, K.J. Dengue Virus NS1 Protein Activates Immune Cells via TLR4 but Not TLR2 or TLR6. Immunol. Cell Biol. 2017, 95, 491–495. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.-Y.; Cheng, M.-H.; Yu, C.-Y.; Lin, Y.-S.; Yeh, T.-M.; Chen, C.-L.; Chen, C.-C.; Wan, S.-W.; Chang, C.-P. Dengue Nonstructural Protein 1 Maintains Autophagy through Retarding Caspase-Mediated Cleavage of Beclin-1. Int. J. Mol. Sci. 2020, 21, 9702. [Google Scholar] [CrossRef] [PubMed]
- Suputtamongkol, Y.; Avirutnan, P.; Mairiang, D.; Angkasekwinai, N.; Niwattayakul, K.; Yamasmith, E.; Saleh-arong, F.A.; Songjaeng, A.; Prommool, T.; Tangthawornchaikul, N.; et al. Ivermectin Accelerates Circulating Nonstructural Protein 1 (NS1) Clearance in Adult Dengue Patients: A Combined Phase 2/3 Randomized Double-Blinded Placebo Controlled Trial. Clin. Infect. Dis. 2021, 72, e586–e593. [Google Scholar] [CrossRef] [PubMed]
- Chuang, Y.-C.; Lei, H.-Y.; Lin, Y.-S.; Liu, H.-S.; Wu, H.-L.; Yeh, T.-M. Dengue Virus-Induced Autoantibodies Bind to Plasminogen and Enhance Its Activation. J. Immunol. 2011, 187, 6483–6490. [Google Scholar] [CrossRef]
- Chuang, Y.-C.; Lin, Y.-S.; Liu, H.-S.; Yeh, T.-M. Molecular Mimicry between Dengue Virus and Coagulation Factors Induces Antibodies To Inhibit Thrombin Activity and Enhance Fibrinolysis. J. Virol. 2014, 88, 13759–13768. [Google Scholar] [CrossRef]
- Reyes-Sandoval, A.; Ludert, J.E. The Dual Role of the Antibody Response Against the Flavivirus Non-Structural Protein 1 (NS1) in Protection and Immuno-Pathogenesis. Front. Immunol. 2019, 10, 1651. [Google Scholar] [CrossRef]
- Lee, P.X.; Ting, D.H.R.; Boey, C.P.H.; Tan, E.T.X.; Chia, J.Z.H.; Idris, F.; Oo, Y.; Ong, L.C.; Chua, Y.L.; Hapuarachchi, C.; et al. Relative Contribution of Nonstructural Protein 1 in Dengue Pathogenesis. J. Exp. Med. 2020, 217, e20191548. [Google Scholar] [CrossRef]
- Puerta-Guardo, H.; Tabata, T.; Petitt, M.; Dimitrova, M.; Glasner, D.R.; Pereira, L.; Harris, E. Zika Virus Nonstructural Protein 1 Disrupts Glycosaminoglycans and Causes Permeability in Developing Human Placentas. J. Infect. Dis. 2020, 221, 313–324. [Google Scholar] [CrossRef]
- Xia, H.; Luo, H.; Shan, C.; Muruato, A.E.; Nunes, B.T.D.; Medeiros, D.B.A.; Zou, J.; Xie, X.; Giraldo, M.I.; Vasconcelos, P.F.C.; et al. An Evolutionary NS1 Mutation Enhances Zika Virus Evasion of Host Interferon Induction. Nat. Commun. 2018, 9, 414. [Google Scholar] [CrossRef]
- Wilson, J.R.; de Sessions, P.F.; Leon, M.A.; Scholle, F. West Nile Virus Nonstructural Protein 1 Inhibits TLR3 Signal Transduction. J. Virol. 2008, 82, 8262. [Google Scholar] [CrossRef]
- Xu, X.; Song, H.; Qi, J.; Liu, Y.; Wang, H.; Su, C.; Shi, Y.; Gao, G.F. Contribution of Intertwined Loop to Membrane Association Revealed by Zika Virus Full-length NS1 Structure. EMBO J. 2016, 35, 2170–2178. [Google Scholar] [CrossRef]
- Akey, D.L.; Brown, W.C.; Jose, J.; Kuhn, R.J.; Smith, J.L. Structure- Guided Insights on the Role of NS1 in Flavivirus Infection. Bioessays 2015, 37, 489–494. [Google Scholar] [CrossRef]
- Noisakran, S.; Dechtawewat, T.; Avirutnan, P.; Kinoshita, T.; Siripanyaphinyo, U.; Puttikhunt, C.; Kasinrerk, W.; Malasit, P.; Sittisombut, N. Association of Dengue Virus NS1 Protein with Lipid Rafts. J. Gen. Virol. 2008, 89, 2492–2500. [Google Scholar] [CrossRef]
- Youn, S.; Cho, H.; Fremont, D.H.; Diamond, M.S. A Short N-Terminal Peptide Motif on Flavivirus Nonstructural Protein NS1 Modulates Cellular Targeting and Immune Recognition. J. Virol. 2010, 84, 9516–9532. [Google Scholar] [CrossRef]
- Fan, W.; Mason, P.W. Membrane Association and Secretion of the Japanese Encephalitis Virus NS1 Protein from Cells Expressing NS1 CDNA. Virology 1990, 177, 470–476. [Google Scholar] [CrossRef]
- Schlesinger, J.J.; Brandriss, M.W.; Walsh, E.E. Protection against 17D Yellow Fever Encephalitis in Mice by Passive Transfer of Monoclonal Antibodies to the Nonstructural Glycoprotein Gp48 and by Active Immunization with Gp48. J. Immunol. 1985, 135, 2805–2809. [Google Scholar]
- Gould, E.A.; Buckley, A.; Barrett, A.D.; Cammack, N. Neutralizing (54K) and Non-Neutralizing (54K and 48K) Monoclonal Antibodies against Structural and Non-Structural Yellow Fever Virus Proteins Confer Immunity in Mice. J. Gen. Virol. 1986, 67 Pt 3, 591–595. [Google Scholar] [CrossRef]
- Hall, R.A.; Brand, T.N.; Lobigs, M.; Sangster, M.Y.; Howard, M.J.; Mackenzie, J.S. Protective Immune Responses to the E and NS1 Proteins of Murray Valley Encephalitis Virus in Hybrids of Flavivirus-Resistant Mice. J. Gen. Virol. 1996, 77 Pt 6, 1287–1294. [Google Scholar] [CrossRef]
- Hall, R.A.; Kay, B.H.; Burgess, G.W.; Clancy, P.; Fanning, I.D. Epitope Analysis of the Envelope and Non-Structural Glycoproteins of Murray Valley Encephalitis Virus. J. Gen. Virol. 1990, 71 Pt 12, 2923–2930. [Google Scholar] [CrossRef]
- Chen, C.-L.; Lin, C.-F.; Wan, S.-W.; Wei, L.-S.; Chen, M.-C.; Yeh, T.-M.; Liu, H.-S.; Anderson, R.; Lin, Y.-S. Anti–Dengue Virus Nonstructural Protein 1 Antibodies Cause NO-Mediated Endothelial Cell Apoptosis via Ceramide-Regulated Glycogen Synthase Kinase-3β and NF-ΚB Activation. J. Immunol. 2013, 191, 1744–1752. [Google Scholar] [CrossRef]
- Lin, C.-F.; Chiu, S.-C.; Hsiao, Y.-L.; Wan, S.-W.; Lei, H.-Y.; Shiau, A.-L.; Liu, H.-S.; Yeh, T.-M.; Chen, S.-H.; Liu, C.-C.; et al. Expression of Cytokine, Chemokine, and Adhesion Molecules during Endothelial Cell Activation Induced by Antibodies against Dengue Virus Nonstructural Protein 1. J. Immunol. 2005, 174, 395–403. [Google Scholar] [CrossRef]
- Biering, S.B.; Akey, D.L.; Wong, M.P.; Brown, W.C.; Lo, N.T.N.; Puerta-Guardo, H.; Tramontini Gomes de Sousa, F.; Wang, C.; Konwerski, J.R.; Espinosa, D.A.; et al. Structural Basis for Antibody Inhibition of Flavivirus NS1-Triggered Endothelial Dysfunction. Science 2021, 371, 194–200. [Google Scholar] [CrossRef]
- Chung, K.M.; Thompson, B.S.; Fremont, D.H.; Diamond, M.S. Antibody Recognition of Cell Surface-Associated NS1 Triggers Fc-γ Receptor-Mediated Phagocytosis and Clearance of West Nile Virus-Infected Cells. J. Virol. 2007, 81, 9551–9555. [Google Scholar] [CrossRef]
- Chung, K.M.; Nybakken, G.E.; Thompson, B.S.; Engle, M.J.; Marri, A.; Fremont, D.H.; Diamond, M.S. Antibodies against West Nile Virus Nonstructural Protein NS1 Prevent Lethal Infection through Fc γ Receptor-Dependent and -Independent Mechanisms. J. Virol. 2006, 80, 1340–1351. [Google Scholar] [CrossRef]
- Yu, L.; Liu, X.; Ye, X.; Su, W.; Zhang, X.; Deng, W.; Luo, J.; Xiang, M.; Guo, W.; Zhang, S.; et al. Monoclonal Antibodies against Zika Virus NS1 Protein Confer Protection via Fcγ Receptor-Dependent and -Independent Pathways. mBio 2021, 12, e03179-20. [Google Scholar] [CrossRef]
- Wessel, A.W.; Kose, N.; Bombardi, R.G.; Roy, V.; Chantima, W.; Mongkolsapaya, J.; Edeling, M.A.; Nelson, C.A.; Bosch, I.; Alter, G.; et al. Antibodies Targeting Epitopes on the Cell-Surface Form of NS1 Protect against Zika Virus Infection during Pregnancy. Nat. Commun. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Modhiran, N.; Song, H.; Liu, L.; Bletchly, C.; Brillault, L.; Amarilla, A.A.; Xu, X.; Qi, J.; Chai, Y.; Cheung, S.T.M.; et al. A Broadly Protective Antibody That Targets the Flavivirus NS1 Protein. Science 2021, 371, 190–194. [Google Scholar] [CrossRef]
- Edeling, M.A.; Diamond, M.S.; Fremont, D.H. Structural Basis of Flavivirus NS1 Assembly and Antibody Recognition. Proc. Natl. Acad. Sci. USA 2014, 111, 4285–4290. [Google Scholar] [CrossRef] [PubMed]
- Poonsiri, T.; Wright, G.S.A.; Diamond, M.S.; Turtle, L.; Solomon, T.; Antonyuk, S.V. Structural Study of the C-Terminal Domain of Nonstructural Protein 1 from Japanese Encephalitis Virus. J. Virol. 2018, 92, e01868-17. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; Pei, C.; Liu, Z.; Yang, K.; Li, Q.; Chen, H.; Cao, S.; Song, Y. Identification of a Protective Epitope in Japanese Encephalitis Virus NS1 Protein. Antiviral Res. 2020, 182, 104930. [Google Scholar] [CrossRef] [PubMed]
- Sun, E.-C.; Ma, J.-N.; Liu, N.-H.; Yang, T.; Zhao, J.; Geng, H.-W.; Wang, L.-F.; Qin, Y.-L.; Bu, Z.-G.; Yang, Y.-H.; et al. Identification of Two Linear B-Cell Epitopes from West Nile Virus NS1 by Screening a Phage-Displayed Random Peptide Library. BMC Microbiol. 2011, 11, 160. [Google Scholar] [CrossRef]
- Wang, B.; Hua, R.-H.; Tian, Z.-J.; Chen, N.-S.; Zhao, F.-R.; Liu, T.-Q.; Wang, Y.-F.; Tong, G.-Z. Identification of a Virus-Specific and Conserved B-Cell Epitope on NS1 Protein of Japanese Encephalitis Virus. Virus Res. 2009, 141, 90–95. [Google Scholar] [CrossRef]
- Hua, R.-H.; Liu, L.-K.; Huo, H.; Li, Y.-N.; Guo, L.-P.; Wang, X.-L.; Qin, C.-F.; Bu, Z.-G. Comprehensive Mapping of a Novel NS1 Epitope Conserved in Flaviviruses within the Japanese Encephalitis Virus Serocomplex. Virus Res. 2014, 185, 103–109. [Google Scholar] [CrossRef]
- Rocha, L.B.; Alves, R.P.D.S.; Caetano, B.A.; Pereira, L.R.; Mitsunari, T.; Amorim, J.H.; Polatto, J.M.; Botosso, V.F.; Gallina, N.M.F.; Palacios, R.; et al. Epitope Sequences in Dengue Virus NS1 Protein Identified by Monoclonal Antibodies. Antibodies 2017, 6, 14. [Google Scholar] [CrossRef]
- Omokoko, M.D.; Pambudi, S.; Phanthanawiboon, S.; Masrinoul, P.; Setthapramote, C.; Sasaki, T.; Kuhara, M.; Ramasoota, P.; Yamashita, A.; Hirai, I.; et al. A Highly Conserved Region Between Amino Acids 221 and 266 of Dengue Virus Non-Structural Protein 1 Is a Major Epitope Region in Infected Patients. Am. J. Trop. Med. Hyg. 2014, 91, 146–155. [Google Scholar] [CrossRef]
- Wu, H.-C.; Huang, Y.-L.; Chao, T.-T.; Jan, J.-T.; Huang, J.-L.; Chiang, H.-Y.; King, C.-C.; Shaio, M.-F. Identification of B-Cell Epitope of Dengue Virus Type 1 and Its Application in Diagnosis of Patients. J. Clin. Microbiol. 2001, 39, 977–982. [Google Scholar] [CrossRef]
- Tang, Y.-L.; Liu, I.-J.; Li, P.-C.; Chiu, C.-Y.; Lin, C.-Y.; Huang, C.-H.; Chen, Y.-H.; Fu, C.-Y.; Chao, D.-Y.; King, C.-C.; et al. Generation and Characterization of Antinonstructural Protein 1 Monoclonal Antibodies and Development of Diagnostics for Dengue Virus Serotype 2. Am. J. Trop. Med. Hyg. 2017, 97, 1049–1061. [Google Scholar] [CrossRef]
- Falconar, A.K.; Young, P.R.; Miles, M.A. Precise Location of Sequential Dengue Virus Subcomplex and Complex B Cell Epitopes on the Nonstructural-1 Glycoprotein. Arch. Virol. 1994, 137, 315–326. [Google Scholar] [CrossRef]
- Lee, H.-J.; Cho, Y.; Kang, H.J.; Choi, H.; Han, K.R.; Chong, C.K.; Kim, Y.B. Identification of Peptide Based B-Cell Epitopes in Zika Virus NS1. Biochem. Biophys. Res. Commun. 2018, 505, 1010–1014. [Google Scholar] [CrossRef]
- Bugrysheva, J.V.; Matveeva, V.A.; Dobrikova, E.Y.; Bykovskaya, N.V.; Korobova, S.A.; Bakhvalova, V.N.; Morozova, O.V. Tick-Borne Encephalitis Virus NS1 Glycoprotein during Acute and Persistent Infection of Cells. Virus Res. 2001, 76, 161–169. [Google Scholar] [CrossRef]
- Ti, J.; Li, Z.; Li, X.; Lu, Y.; Diao, Y.; Li, F. Identification of One B-Cell Epitope from NS1 Protein of Duck Tembusu Virus with Monoclonal Antibodies. PLoS ONE 2017, 12, e0181177. [Google Scholar] [CrossRef]
- Bailey, M.J.; Broecker, F.; Duehr, J.; Arumemi, F.; Krammer, F.; Palese, P.; Tan, G.S. Antibodies Elicited by an NS1-Based Vaccine Protect Mice against Zika Virus. mBio 2019, 10, e02861-18. [Google Scholar] [CrossRef]
- Avirutnan, P.; Fuchs, A.; Hauhart, R.E.; Somnuke, P.; Youn, S.; Diamond, M.S.; Atkinson, J.P. Antagonism of the Complement Component C4 by Flavivirus Nonstructural Protein NS1. J. Exp. Med. 2010, 207, 793–806. [Google Scholar] [CrossRef]
- Charles, A.; Janeway, J.; Travers, P.; Walport, M.; Shlomchik, M.J. The Complement System and Innate Immunity. In Immunobiology: The Immune System in Health and Disease, 5th ed.; Garland Science: New York, NY, USA, 2001. [Google Scholar]
- Bokisch, V.A.; Top, F.H.; Russell, P.K.; Dixon, F.J.; Müller-Eberhard, H.J. The Potential Pathogenic Role of Complement in Dengue Hemorrhagic Shock Syndrome. N. Engl. J. Med. 1973, 289, 996–1000. [Google Scholar] [CrossRef]
- Hodes, H.L.; Thomas, L.; Peck, J.L. Cause of an Outbreak of Encephalitis Established by Means of Complement-Fixation Tests. Proc. Soc. Exp. Biol. Med. 1945, 60, 220–225. [Google Scholar] [CrossRef]
- Hirota, J.; Shimizu, S.; Shibahara, T.; Isobe, T.; Yamada, M.; Tanimura, N. Development of Monoclonal Antibodies to West Nile Virus and Their Application in Immunohistochemistry. Clin. Vaccine Immunol. CVI 2012, 19, 1853–1858. [Google Scholar] [CrossRef]
- Falkler, W.A.; Diwan, A.R.; Halstead, S.B. Human Antibody to Dengue Soluble Complement-Fixing (SCF) Antigens. J. Immunol. 1973, 111, 1804–1809. [Google Scholar]
- Monath, T.P.; Craven, R.B.; Muth, D.J.; Trautt, C.J.; Calisher, C.H.; Fitzgerald, S.A. Limitations of the Complement-Fixation Test for Distinguishing Naturally Acquired from Vaccine-Induced Yellow Fever Infection in Flavivirus-Hyperendemic Areas. Am. J. Trop. Med. Hyg. 1980, 29, 624–634. [Google Scholar] [CrossRef]
- Idris, F.; Muharram, S.H.; Diah, S. Glycosylation of Dengue Virus Glycoproteins and Their Interactions with Carbohydrate Receptors: Possible Targets for Antiviral Therapy. Arch. Virol. 2016, 161, 1751–1760. [Google Scholar] [CrossRef]
- Pryor, M.J.; Wright, P.J. Glycosylation Mutants of Dengue Virus NS1 Protein. J. Gen. Virol. 1994, 75, 1183–1187. [Google Scholar] [CrossRef]
- Warfield, K.L.; Plummer, E.; Alonzi, D.S.; Wolfe, G.W.; Sampath, A.; Nguyen, T.; Butters, T.D.; Enterlein, S.G.; Stavale, E.J.; Shresta, S.; et al. A Novel Iminosugar UV-12 with Activity against the Diverse Viruses Influenza and Dengue (Novel Iminosugar Antiviral for Influenza and Dengue). Viruses 2015, 7, 2404–2427. [Google Scholar] [CrossRef]
- Perry, S.T.; Buck, M.D.; Plummer, E.M.; Penmasta, R.A.; Batra, H.; Stavale, E.J.; Warfield, K.L.; Dwek, R.A.; Butters, T.D.; Alonzi, D.S.; et al. An Iminosugar with Potent Inhibition of Dengue Virus Infection in Vivo. Antiviral Res. 2013, 98, 35–43. [Google Scholar] [CrossRef]
- Mohan, S.; McAtamney, S.; Jayakanthan, K.; Eskandari, R.; von Itzstein, M.; Pinto, B.M. Antiviral Activities of Sulfonium-Ion Glucosidase Inhibitors and 5-Thiomannosylamine Disaccharide Derivatives against Dengue Virus. Int. J. Antimicrob. Agents 2012, 40, 273–276. [Google Scholar] [CrossRef]
- Watanabe, S.; Rathore, A.P.S.; Sung, C.; Lu, F.; Khoo, Y.M.; Connolly, J.; Low, J.; Ooi, E.E.; Lee, H.S.; Vasudevan, S.G. Dose- and Schedule-Dependent Protective Efficacy of Celgosivir in a Lethal Mouse Model for Dengue Virus Infection Informs Dosing Regimen for a Proof of Concept Clinical Trial. Antiviral Res. 2012, 96, 32–35. [Google Scholar] [CrossRef]
- Chang, J.; Wang, L.; Ma, D.; Qu, X.; Guo, H.; Xu, X.; Mason, P.M.; Bourne, N.; Moriarty, R.; Gu, B.; et al. Novel Imino Sugar Derivatives Demonstrate Potent Antiviral Activity against Flaviviruses. Antimicrob. Agents Chemother. 2009, 53, 1501–1508. [Google Scholar] [CrossRef]
- Rathore, A.P.S.; Paradkar, P.N.; Watanabe, S.; Tan, K.H.; Sung, C.; Connolly, J.E.; Low, J.; Ooi, E.E.; Vasudevan, S.G. Celgosivir Treatment Misfolds Dengue Virus NS1 Protein, Induces Cellular pro-Survival Genes and Protects against Lethal Challenge Mouse Model. Antivir. Res. 2011, 92, 453–460. [Google Scholar] [CrossRef]
- Low, J.G.; Sung, C.; Wijaya, L.; Wei, Y.; Rathore, A.P.S.; Watanabe, S.; Tan, B.H.; Toh, L.; Chua, L.T.; Hou, Y.; et al. Efficacy and Safety of Celgosivir in Patients with Dengue Fever (CELADEN): A Phase 1b, Randomised, Double-Blind, Placebo-Controlled, Proof-of-Concept Trial. Lancet Infect. Dis. 2014, 14, 706–715. [Google Scholar] [CrossRef]
- Whitby, K.; Pierson, T.C.; Geiss, B.; Lane, K.; Engle, M.; Zhou, Y.; Doms, R.W.; Diamond, M.S. Castanospermine, a Potent Inhibitor of Dengue Virus Infection In Vitro and In Vivo. J. Virol. 2005, 79, 8698–8706. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.L.; Cherepanova, N.A.; Bozzacco, L.; MacDonald, M.R.; Gilmore, R.; Tai, A.W. Dengue Virus Hijacks a Noncanonical Oxidoreductase Function of a Cellular Oligosaccharyltransferase Complex. mBio 2017, 8, e00939-17. [Google Scholar] [CrossRef] [PubMed]
- Winkler, G.; Maxwell, S.E.; Ruemmler, C.; Stollar, V. Newly Synthesized Dengue-2 Virus Nonstructural Protein NS1 Is a Soluble Protein but Becomes Partially Hydrophobic and Membrane-Associated after Dimerization. Virology 1989, 171, 302–305. [Google Scholar] [CrossRef]
- Whiteman, M.C.; Popov, V.; Sherman, M.B.; Wen, J.; Barrett, A.D.T. Attenuated West Nile Virus Mutant NS1130-132QQA/175A/207A Exhibits Virus-Induced Ultrastructural Changes and Accumulation of Protein in the Endoplasmic Reticulum. J. Virol. 2015, 89, 1474–1478. [Google Scholar] [CrossRef]
- Somnuke, P.; Hauhart, R.E.; Atkinson, J.P.; Diamond, M.S.; Avirutnan, P. N-Linked Glycosylation of Dengue Virus NS1 Protein Modulates Secretion, Cell-Surface Expression, Hexamer Stability, and Interactions with Human Complement. Virology 2011, 413, 253–264. [Google Scholar] [CrossRef]
- Tajima, S.; Takasaki, T.; Kurane, I. Characterization of Asn130-to-Ala Mutant of Dengue Type 1 Virus NS1 Protein. Virus Genes 2008, 36, 323–329. [Google Scholar] [CrossRef]
- Pletnev, A.G.; Bray, M.; Lai, C.J. Chimeric Tick-Borne Encephalitis and Dengue Type 4 Viruses: Effects of Mutations on Neurovirulence in Mice. J. Virol. 1993, 67, 4956–4963. [Google Scholar] [CrossRef]
- Schlesinger, J.J.; Brandriss, M.W.; Cropp, C.B.; Monath, T.P. Protection against Yellow Fever in Monkeys by Immunization with Yellow Fever Virus Nonstructural Protein NS1. J. Virol. 1986, 60, 1153–1155. [Google Scholar] [CrossRef]
- Schlesinger, J.J.; Brandriss, M.W.; Walsh, E.E. Protection of Mice Against Dengue 2 Virus Encephalitis by Immunization with the Dengue 2 Virus Non-Structural Glycoprotein NS1. J. Gen. Virol. 1987, 68, 853–857. [Google Scholar] [CrossRef]
- Cane, P.A.; Gould, E.A. Reduction of Yellow Fever Virus Mouse Neurovirulence by Immunization with a Bacterially Synthesized Non-Structural Protein (NS1) Fragment. J. Gen. Virol. 1988, 69, 1241–1246. [Google Scholar] [CrossRef]
- Srivastava, A.K.; Putnak, J.R.; Warren, R.L.; Hoke, C.H. Mice Immunized with a Dengue Type 2 Virus E and NS1 Fusion Protein Made in Escherichia Coli Are Protected against Lethal Dengue Virus Infection. Vaccine 1995, 13, 1251–1258. [Google Scholar] [CrossRef]
- Lin, Y.-L.; Chen, L.-K.; Liao, C.-L.; Yeh, C.-T.; Ma, S.-H.; Chen, J.-L.; Huang, Y.-L.; Chen, S.-S.; Chiang, H.-Y. DNA Immunization with Japanese Encephalitis Virus Nonstructural Protein NS1 Elicits Protective Immunity in Mice. J. Virol. 1998, 72, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.-F.; Liao, C.-L.; Lin, Y.-L.; Yeh, C.-T.; Chen, L.-K.; Huang, Y.-F.; Chou, H.-Y.; Huang, J.-L.; Shaio, M.-F.; Sytwu, H.-K. Evaluation of Protective Efficacy and Immune Mechanisms of Using a Non-Structural Protein NS1 in DNA Vaccine against Dengue 2 Virus in Mice. Vaccine 2003, 21, 3919–3929. [Google Scholar] [CrossRef]
- Costa, S.M.; Paes, M.V.; Barreto, D.F.; Pinhão, A.T.; Barth, O.M.; Queiroz, J.L.S.; Armôa, G.R.G.; Freire, M.S.; Alves, A.M.B. Protection against Dengue Type 2 Virus Induced in Mice Immunized with a DNA Plasmid Encoding the Non-Structural 1 (NS1) Gene Fused to the Tissue Plasminogen Activator Signal Sequence. Vaccine 2006, 24, 195–205. [Google Scholar] [CrossRef]
- Li, A.; Xue, M.; Attia, Z.; Yu, J.; Lu, M.; Shan, C.; Liang, X.; Gao, T.Z.; Shi, P.-Y.; Peeples, M.E.; et al. Vesicular Stomatitis Virus and DNA Vaccines Expressing Zika Virus Nonstructural Protein 1 Induce Substantial but Not Sterilizing Protection against Zika Virus Infection. J. Virol. 2020, 94, e00048-20. [Google Scholar] [CrossRef]
- Grubor-Bauk, B.; Wijesundara, D.K.; Masavuli, M.; Abbink, P.; Peterson, R.L.; Prow, N.A.; Larocca, R.A.; Mekonnen, Z.A.; Shrestha, A.; Eyre, N.S.; et al. NS1 DNA Vaccination Protects against Zika Infection through T Cell–Mediated Immunity in Immunocompetent Mice. Sci. Adv. 2019, 5, eaax2388. [Google Scholar] [CrossRef]
- Zhao, B.T.; Prince, G.; Horswood, R.; Eckels, K.; Summers, P.; Chanock, R.; Lai, C.J. Expression of Dengue Virus Structural Proteins and Nonstructural Protein NS1 by a Recombinant Vaccinia Virus. J. Virol. 1987, 61, 4019–4022. [Google Scholar] [CrossRef]
- Putnak, J.R.; Schlesinger, J.J. Protection of Mice against Yellow Fever Virus Encephalitis by Immunization with a Vaccinia Virus Recombinant Encoding the Yellow Fever Virus Non-Structural Proteins, NS1, NS2a and NS2b. J. Gen. Virol. 1990, 71, 1697–1702. [Google Scholar] [CrossRef]
- Falgout, B.; Bray, M.; Schlesinger, J.J.; Lai, C.J. Immunization of Mice with Recombinant Vaccinia Virus Expressing Authentic Dengue Virus Nonstructural Protein NS1 Protects against Lethal Dengue Virus Encephalitis. J. Virol. 1990, 64, 4356–4363. [Google Scholar] [CrossRef]
- Mason, P.W.; Pincus, S.; Fournier, M.J.; Mason, T.L.; Shope, R.E.; Paoletti, E. Japanese Encephalitis Virus-Vaccinia Recombinants Produce Particulate Forms of the Structural Membrane Proteins and Induce High Levels of Protection against Lethal JEV Infection. Virology 1991, 180, 294–305. [Google Scholar] [CrossRef]
- Jacobs, S.C.; Stephenson, J.R.; Wilkinson, G.W. High-Level Expression of the Tick-Borne Encephalitis Virus NS1 Protein by Using an Adenovirus-Based Vector: Protection Elicited in a Murine Model. J. Virol. 1992, 66, 2086–2095. [Google Scholar] [CrossRef]
- Timofeev, A.V.; Ozherelkov, S.V.; Pronin, A.V.; Deeva, A.V.; Karganova, G.G.; Elbert, L.B.; Stephenson, J.R. Immunological Basis for Protection in a Murine Model of Tick-Borne Encephalitis by a Recombinant Adenovirus Carrying the Gene Encoding the NS1 Non-Structural Protein. J. Gen. Virol. 1998, 79, 689–695. [Google Scholar] [CrossRef]
- Li, A.; Yu, J.; Lu, M.; Ma, Y.; Attia, Z.; Shan, C.; Xue, M.; Liang, X.; Craig, K.; Makadiya, N.; et al. A Zika Virus Vaccine Expressing Premembrane-Envelope-NS1 Polyprotein. Nat. Commun. 2018, 9, 3067. [Google Scholar] [CrossRef]
- Brault, A.C.; Domi, A.; McDonald, E.M.; Talmi-Frank, D.; McCurley, N.; Basu, R.; Robinson, H.L.; Hellerstein, M.; Duggal, N.K.; Bowen, R.A.; et al. A Zika Vaccine Targeting NS1 Protein Protects Immunocompetent Adult Mice in a Lethal Challenge Model. Sci. Rep. 2017, 7, 14769. [Google Scholar] [CrossRef]
- Qu, X.; Chen, W.; Maguire, T.; Austin, F. Immunoreactivity and Protective Effects in Mice of a Recombinant Dengue 2 Tonga Virus NS1 Protein Produced in a Baculovirus Expression System. J. Gen. Virol. 1993, 74, 89–97. [Google Scholar] [CrossRef]
- Eckels, K.H.; Dubois, D.R.; Summers, P.L.; Schlesinger, J.J.; Shelly, M.; Cohen, S.; Zhang, Y.-M.; Lai, C.-J.; Kurane, I.; Rothman, A.; et al. Immunization of Monkeys with Baculovirus-Dengue Type-4 Recombinants Containing Envelope and Nonstructural Proteins: Evidence of Priming and Partial Protection. Am. J. Trop. Med. Hyg. 1994, 50, 472–478. [Google Scholar] [CrossRef]
- Desprès, P.; Dietrich, J.; Girard, M.; Bouloy, M. Recombinant Baculoviruses Expressing Yellow Fever Virus E and NS1 Proteins Elicit Protective Immunity in Mice. J. Gen. Virol. 1991, 72, 2811–2816. [Google Scholar] [CrossRef]
- Whiteman, M.C.; Wicker, J.A.; Kinney, R.M.; Huang, C.Y.-H.; Solomon, T.; Barrett, A.D.T. Multiple Amino Acid Changes at the First Glycosylation Motif in NS1 Protein of West Nile Virus Are Necessary for Complete Attenuation for Mouse Neuroinvasiveness. Vaccine 2011, 29, 9702–9710. [Google Scholar] [CrossRef]
- Crabtree, M.B.; Kinney, R.M.; Miller, B.R. Deglycosylation of the NS1 Protein of Dengue 2 Virus, Strain 16681: Construction and Characterization of Mutant Viruses. Arch. Virol. 2005, 150, 771–786. [Google Scholar] [CrossRef]
- Ishikawa, T.; Wang, G.; Widman, D.G.; Infante, E.; Winkelmann, E.R.; Bourne, N.; Mason, P.W. Enhancing the Utility of a PrM/E-Expressing Chimeric Vaccine for Japanese Encephalitis by Addition of the JEV NS1 Gene. Vaccine 2011, 29, 7444–7455. [Google Scholar] [CrossRef]
- Osorio, J.E.; Huang, C.Y.-H.; Kinney, R.M.; Stinchcomb, D.T. Development of DENVax: A Chimeric Dengue-2 PDK-53-Based Tetravalent Vaccine for Protection against Dengue Fever. Vaccine 2011, 29, 7251–7260. [Google Scholar] [CrossRef]
- Butrapet, S.; Huang, C.Y.-H.; Pierro, D.J.; Bhamarapravati, N.; Gubler, D.J.; Kinney, R.M. Attenuation Markers of a Candidate Dengue Type 2 Vaccine Virus, Strain 16681 (PDK-53), Are Defined by Mutations in the 5′ Noncoding Region and Nonstructural Proteins 1 and 3. J. Virol. 2000, 74, 3011–3019. [Google Scholar] [CrossRef]
- Choy, M.M.; Ng, D.H.L.; Siriphanitchakorn, T.; Ng, W.C.; Sundstrom, K.B.; Tan, H.C.; Zhang, S.L.; Chan, K.W.K.; Manuel, M.; Kini, R.M.; et al. A Non-Structural 1 Protein G53D Substitution Attenuates a Clinically Tested Live Dengue Vaccine. Cell Rep. 2020, 31, 107617. [Google Scholar] [CrossRef]
- Chan, K.W.K.; Watanabe, S.; Jin, J.Y.; Pompon, J.; Teng, D.; Alonso, S.; Vijaykrishna, D.; Halstead, S.B.; Marzinek, J.K.; Bond, P.J.; et al. A T164S Mutation in the Dengue Virus NS1 Protein Is Associated with Greater Disease Severity in Mice. Sci. Transl. Med. 2019, 11, eaat7726. [Google Scholar] [CrossRef]
- Mishra, N.; Boudewijns, R.; Schmid, M.A.; Marques, R.E.; Sharma, S.; Neyts, J.; Dallmeier, K. A Chimeric Japanese Encephalitis Vaccine Protects against Lethal Yellow Fever Virus Infection without Inducing Neutralizing Antibodies. mBio 2020, 11, e02494-19. [Google Scholar] [CrossRef]
- Annamalai, A.S.; Pattnaik, A.; Sahoo, B.R.; Muthukrishnan, E.; Natarajan, S.K.; Steffen, D.; Vu, H.L.X.; Delhon, G.; Osorio, F.A.; Petro, T.M.; et al. Zika Virus Encoding Nonglycosylated Envelope Protein Is Attenuated and Defective in Neuroinvasion. J. Virol. 2017, 91, e01348-17. [Google Scholar] [CrossRef]
- Richner, J.M.; Jagger, B.W.; Shan, C.; Fontes, C.R.; Dowd, K.A.; Cao, B.; Himansu, S.; Caine, E.A.; Nunes, B.T.D.; Medeiros, D.B.A.; et al. Vaccine Mediated Protection Against Zika Virus-Induced Congenital Disease. Cell 2017, 170, 273–283.e12. [Google Scholar] [CrossRef]
- Annamalai, A.S.; Pattnaik, A.; Sahoo, B.R.; Guinn, Z.P.; Bullard, B.L.; Weaver, E.A.; Steffen, D.; Natarajan, S.K.; Petro, T.M.; Pattnaik, A.K. An Attenuated Zika Virus Encoding Non-Glycosylated Envelope (E) and Non-Structural Protein 1 (NS1) Confers Complete Protection against Lethal Challenge in a Mouse Model. Vaccines 2019, 7, 112. [Google Scholar] [CrossRef]
- Hahn, C.S.; Dalrymple, J.M.; Strauss, J.H.; Rice, C.M. Comparison of the Virulent Asibi Strain of Yellow Fever Virus with the 17D Vaccine Strain Derived from It. Proc. Natl. Acad. Sci. USA 1987, 84, 2019–2023. [Google Scholar] [CrossRef]
- Collins, N.D.; Beck, A.S.; Widen, S.G.; Wood, T.G.; Higgs, S.; Barrett, A.D.T. Structural and Nonstructural Genes Contribute to the Genetic Diversity of RNA Viruses. mBio 2018, 9, e01871-18. [Google Scholar] [CrossRef]
- Davis, E.H.; Beck, A.S.; Strother, A.E.; Thompson, J.K.; Widen, S.G.; Higgs, S.; Wood, T.G.; Barrett, A.D.T. Attenuation of Live-Attenuated Yellow Fever 17D Vaccine Virus Is Localized to a High-Fidelity Replication Complex. mBio 2019, 10, e02294-19. [Google Scholar] [CrossRef] [PubMed]
- Nitayaphan, S.; Grant, J.A.; Chang, G.-J.J.; Trent, D.W. Nucleotide Sequence of the Virulent SA-14 Strain of Japanese Encephalitis Virus and Its Attenuated Vaccine Derivative, SA-14-14-2. Virology 1990, 177, 541–552. [Google Scholar] [CrossRef]
- Davis, E.H.; Barrett, A.D.T. Japanese Encephalitis Virus Live Attenuated Vaccine Strains Display Altered Immunogenicity, Virulence and Genetic Diversity. 2021; in review. [Google Scholar]
- Li, N.; Zhang, Y.-N.; Deng, C.-L.; Shi, P.-Y.; Yuan, Z.-M.; Zhang, B. Replication-Defective West Nile Virus with NS1 Deletion as a New Vaccine Platform for Flavivirus. J. Virol. 2019, 93. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Zhang, Z.-R.; Zhang, Y.-N.; Liu, J.; Deng, C.-L.; Shi, P.-Y.; Yuan, Z.-M.; Ye, H.-Q.; Zhang, B. A Replication-Defective Japanese Encephalitis Virus (JEV) Vaccine Candidate with NS1 Deletion Confers Dual Protection against JEV and West Nile Virus in Mice. NPJ Vaccines 2020, 5, 73. [Google Scholar] [CrossRef]
- Sarathy, V.V.; Milligan, G.N.; Bourne, N.; Barrett, A.D. Mouse Models of Dengue Virus Infection for Vaccine Testing. Vaccine 2015, 33, 7051–7060. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Monoclonal Antibodies | Antigenic Specificity 1,2,3,4,5,6 | Epitope (NS1 Residues) | Reference |
|---|---|---|---|
| 2B7 | DENV complex | [52] | |
| 3D1.4 | DENV complex | 111–121 | [53] |
| 1A12.3 | DENV complex | 111–121 | [53] |
| 4H3.4 | DENV complex | 111–121 | [53] |
| 3A5.4 | DENV complex | 111–121 | [53] |
| 33D2 | DENV complex | 109–122 | [14] |
| 19–5 | DENV complex | 109–122 | [14] |
| DB16-1 | DENV complex | [54] | |
| DB20-6 | DENV complex | 115–120 | [54] |
| DB29-1 | DENV complex | 115–120 | [54] |
| 15F3-1 | DENV1 | 110–117 | [55] |
| 4F6 | DENV2 | 25–38 | [56] |
| 4H2 | DENV2 | 127–143 | [56] |
| DB6-1 | DENV2 | [54] | |
| DB12-3 | DENV2 | [54] | |
| DB38-1 | DENV2 | [54] | |
| DB41-2 | DENV2 & DENV4 | [54] | |
| 5H5.4 | DENV2 & DENV4 | 299–309 | [53] |
| 1G5.3 | DENV2 & DENV4 | 299–309 | [53,57] |
| 1H6 | JEV | 146–150 | [58] |
| 10F7 | JEV | 1–16 | [59] |
| 8B5 | JEV | 65–80 | [59] |
| 7G1 | JEV | 249–264 | [59] |
| 7C11 | JEV | 265–280 | [59] |
| 4E3 | JEV | 265–280 | [59] |
| HA12 | JEV | 265–280 | [59] |
| 3H11 | JEV | 337–352 | [59] |
| 3G11 | JEV | 337–352 | [59] |
| 4E8 | JEV | 337–352 | [59] |
| 5C6 | JEV | 337–352 | [59] |
| 6D8 | JEV | 337–352 | [59] |
| 2B8 | JEV | 225–233 | [60] |
| 1E8 | JEV serocomplex | 227–232 | [59] |
| 4D1 | JEV serocomplex | 925–934 | [61] |
| 22NS1 | JEV serocomplex | 172–352 | [58] |
| 20B4 | TBEV | 269–333 | [62] |
| 29G9 | TBEV | 1–33 | [62] |
| 3G2 | Tembusu virus | 269–274 | [63] |
| 3C7 | WNV | 895–901 | [61] |
| 10NS1 | WNV | 1–157 & 158–235 | [64] |
| 14NS1 | WNV | 236–352 | [64] |
| 16NS1 | WNV | 1–157 | [64] |
| 17NS1 | WNV | 1–157 & 236–352 | [64] |
| 428, 423, 992, 917, 999, 979, 871, 925 | YFV & Alfuy virus | [51] | |
| Z11 | ZIKV | 102 | [65] |
| Z15 | ZIKV | 146 | [65] |
| Z18 | ZIKV | 102 | [65] |
| Z17 | ZIKV | 289 & 338 | [65] |
| 749-A4 | ZIKV | 289 & 338 | [65] |
| ZIKV-292 | ZIKV | 101 & 177–178 | [65] |
| ZIKV-231 | ZIKV | 265 & 314 | [65] |
| 2H5 | ZIKV & DENV2 | 193–209 | [56] |
| 4H1BC | ZIKV & DENV2 | 193–209 | [56] |
| Replication | Immune Response | Pathogenesis |
|---|---|---|
| Formation of replication complex | Suppress IFN-β induction | Promotes vascular leakage and platelet aggregation in DENV infection |
| Virus assembly | Promote complement-mediated cytolysis | Contributes to hyperpermeability in umbilical vein and brain endothelial cells in ZIKV infection |
| Virus maturation | Promotes antibody-dependent cellular cytotoxicity (ADCC) | YFV infects liver |
| Antibody-dependent cellular phagocytosis (ADCP) | JE serocomplex viruses are neuroinvasive |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carpio, K.L.; Barrett, A.D.T. Flavivirus NS1 and Its Potential in Vaccine Development. Vaccines 2021, 9, 622. https://doi.org/10.3390/vaccines9060622
Carpio KL, Barrett ADT. Flavivirus NS1 and Its Potential in Vaccine Development. Vaccines. 2021; 9(6):622. https://doi.org/10.3390/vaccines9060622
Chicago/Turabian StyleCarpio, Kassandra L., and Alan D. T. Barrett. 2021. "Flavivirus NS1 and Its Potential in Vaccine Development" Vaccines 9, no. 6: 622. https://doi.org/10.3390/vaccines9060622
APA StyleCarpio, K. L., & Barrett, A. D. T. (2021). Flavivirus NS1 and Its Potential in Vaccine Development. Vaccines, 9(6), 622. https://doi.org/10.3390/vaccines9060622