
Scale-Up Production of Type O and A Foot-and-Mouth Disease Bivalent Vaccine and Its Protective Efficacy in Pigs

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Cells
2.2. Viruses
2.3. Determination of Optimal Conditions to Propagate FMDV on a Flask Scale
2.4. Production of FMD Vaccine Antigens on a Pilot Scale
2.5. Virus Titration
2.6. Virus Neutralization Test
2.7. Quantification of Vaccine Antigen (146S)
2.8. Transmission Electron Microscopy
2.9. Immune Responses in Guinea Pigs Vaccinated with the Bivalent Vaccine
2.10. Vaccine Potency Test in Pigs
2.11. Ethical Statement
2.12. Statistical Analysis
3. Results
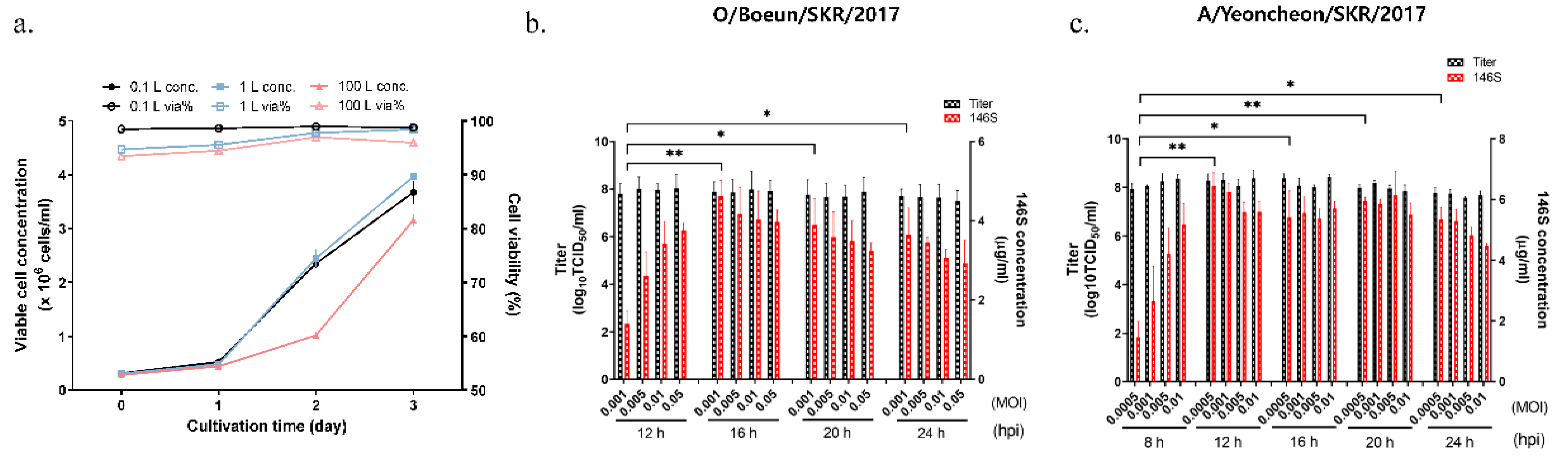
3.1. Determination of Optimal Conditions for Inoculating FMDV onto Suspension Cells
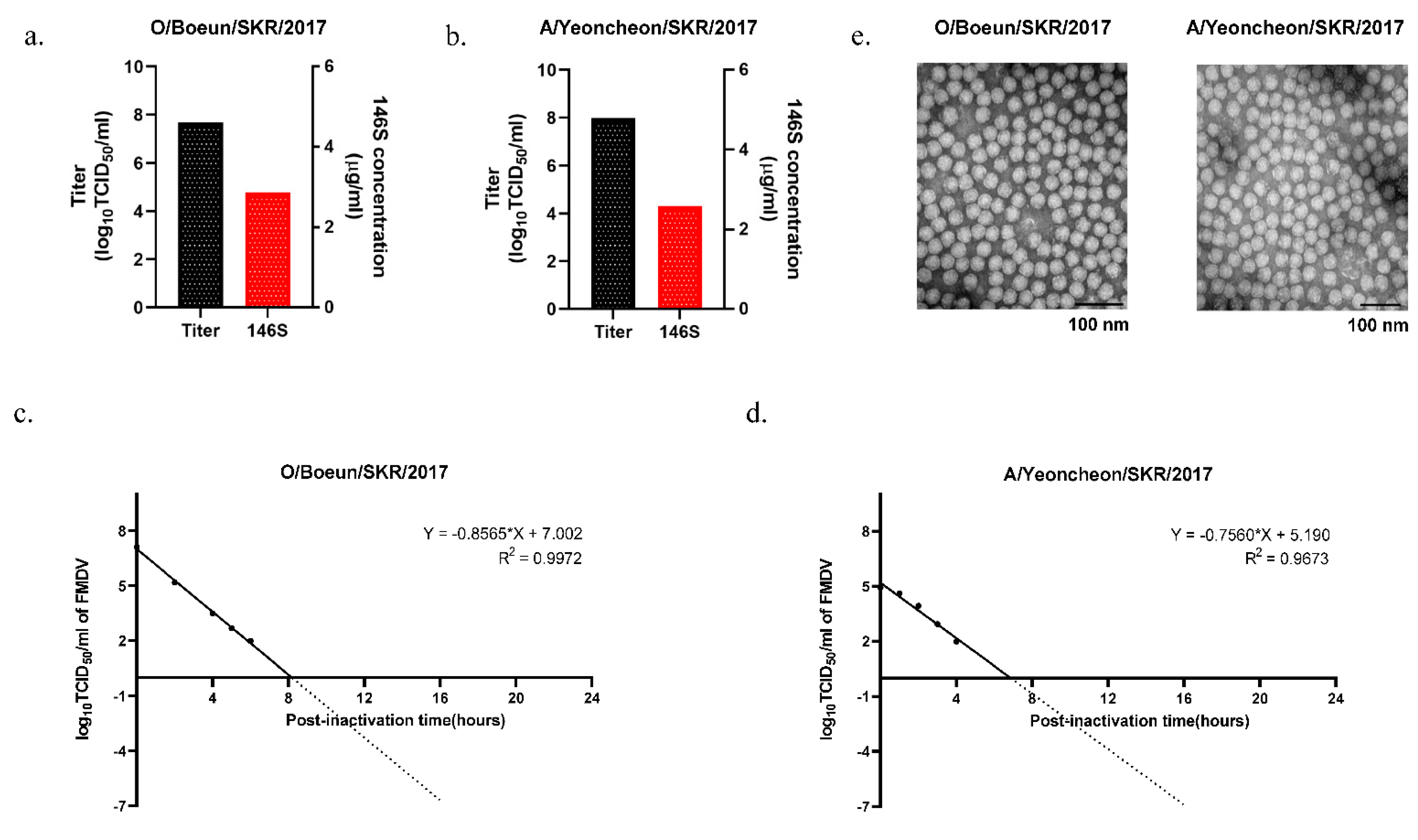
3.2. FMDV Propagation in a 100 L Bioreactor
3.3. Immune Responses in Guinea Pigs Vaccinated with the Bivalent Vaccine
3.4. Protective Dose of the Bivalent Vaccine in Pigs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jamal, S.M.; Belsham, G.J. Foot-and-mouth disease: Past, present and future. Vet. Res. 2013, 44, 116. [Google Scholar] [CrossRef]
- Domingo, E.; Baranowski, E.; Escarmís, C.; Sobrino, F. Foot-and-mouth disease virus. Comp. Immunol. Microbiol. Infect. Dis. 2002, 25, 297–308. [Google Scholar] [CrossRef]
- Mahapatra, M.; Upadhyaya, S.; Aviso, S.; Babu, A.; Hutchings, G.; Parida, S. Selection of vaccine strains for serotype O foot-and-mouth disease viruses (2007-2012) circulating in Southeast Asia, East Asia and Far East. Vaccine 2017, 35, 7147–7153. [Google Scholar] [CrossRef] [PubMed]
- Blacksell, S.D.; Siengsanan-Lamont, J.; Kamolsiripichaiporn, S.; Gleeson, L.J.; Windsor, P.A. A history of FMD research and control programmes in Southeast Asia: Lessons from the past informing the future. Epidemiol. Infect. 2019, 147, 1–13. [Google Scholar] [CrossRef]
- Vierra, D.; Bertram, M.R.; Palinski, R.M.; Pauszek, S.J.; Hartwig, E.J.; Smoliga, G.R.; Vu, L.T.; Hoang, B.H.; Phuong, N.T.; Hung, V.V.; et al. Foot-and-mouth disease virus serotype O/CATHAY genome sequences from five outbreaks in Vietnam, 2017 to 2019. Microbiol. Res. Announc. 2020, 9, e01315–e01319. [Google Scholar] [CrossRef] [PubMed]
- Singanallur, N.B.; Dekker, A.; Eble, P.L.; van Hemert-Kluitenberg, F.; Weerdmeester, K.; Horsington, J.; Vosloo, W.W. Emergency foot-and-mouth disease vaccines A Malaysia 97 and A22 Iraq 64 offer good protection against heterologous challenge with A variant serotype A ASIA/G-IX/SEA-97 lineage virus. Vaccines 2020, 8, 80. [Google Scholar] [CrossRef] [PubMed]
- Mahapatra, M.; Parida, S. Foot and mouth disease vaccine strain selection: Current approaches and future perspectives. Expert Rev. Vaccines 2018, 17, 577–591. [Google Scholar] [CrossRef]
- Lee, G.; Hwang, J.H.; Park, J.H.; Lee, M.J.; Kim, B.; Kim, S.M. Vaccine strain of O/ME-SA/Ind-2001e of foot-and-mouth disease virus provides high immunogenicity and broad antigenic coverage. Antiviral Res. 2020, 182, 104920. [Google Scholar] [CrossRef]
- Lee, S.-Y.; Lee, Y.-J.; Kim, R.-H.; Park, J.-N.; Park, M.-E.; Ko, M.-K.; Choi, J.-H.; Chu, J.-Q.; Lee, K.-N.; Kim, S.-M.; et al. Rapid engineering of foot-and-mouth disease vaccine and challenge viruses. J. Virol. 2017, 91, e00115–e00117. [Google Scholar] [CrossRef]
- Spearman, C. The method of ‘right and wrong cases’ (‘constant stimuli’) without Gauss’s formulae. Br. J. Psychol. 1908, 2, 227–242. [Google Scholar] [CrossRef]
- Kärber, G. Beitrag zur kollektiven Behandlung pharmakologischer Reihenversuche. Naunyn-Schmiedebergs Archiv für experimentelle Pathologie und Pharmakologie 1931, 162, 480–483. [Google Scholar] [CrossRef]
- International Committee; Biological Standards Commission; International Office of Epizootics. Manual of Diagnostic Tests and Vaccines for Terrestrial Animals: Mammals, Birds, and Bees; Renouf Publishing Co. Ltd.: Ottawa, ON, Canada, 2018; pp. 433–464. Available online: https://www.oie.int/fileadmin/Home/eng/Health_standards/tahm/3.01.08_FMD.pdf (accessed on 5 October 2020).
- Doel, T.R.; Baccarini, P.J. Thermal stability of foot-and-mouth disease virus. Arch. Virol. 1981, 70, 21–32. [Google Scholar] [CrossRef]
- Alves, M.P.; Guzylack-Piriou, L.; Juillard, V.; Audonnet, J.C.; Doel, T.; Dawson, H.; Golde, W.T.; Gerber, H.; Peduto, N.; McCullough, K.C.; et al. Innate immune defenses induced by CpG do not promote vaccine-induced protection against foot-and-mouth disease virus in pigs. Clin. Vaccine Immunol. 2009, 16, 1151–1157. [Google Scholar] [CrossRef] [PubMed]
- Brito, B.; Pauszek, S.J.; Hartwig, E.J.; Smoliga, G.R.; Vu, L.T.; Dong, P.V.; Stenfeldt, C.; Rodriguez, L.L.; King, D.P.; Knowles, N.J.; et al. A traditional evolutionary history of foot-and-mouth disease viruses in Southeast Asia challenged by analyses of non-structural protein coding sequences. Sci. Rep. 2018, 8, 6472. [Google Scholar] [CrossRef] [PubMed]
- Horsington, J.; Zhang, Z.; Bittner, H.; Hole, K.; Singanallur, N.B.; Alexandersen, S.; Vosloo, W. Early protection in sheep against intratypic heterologous challenge with serotype O foot-and-mouth disease virus using high-potency, emergency vaccine. Vaccine 2015, 33, 422–429. [Google Scholar] [CrossRef] [PubMed]
- Gallo-Ramirez, L.E.; Nikolay, A.; Genzel, Y.; Reichl, U. Bioreactor concepts for cell culture-based viral vaccine production. Expert Rev. Vaccines 2015, 14, 1181–1195. [Google Scholar] [CrossRef]
- Li, X.R.; Yang, Y.K.; Wang, R.B.; An, F.L.; Zhang, Y.D.; Nie, J.Q.; Ahamada, H.; Liu, X.X.; Liu, C.L.; Deng, Y.; et al. A scale-down model of 4000-L cell culture process for inactivated foot-and-mouth disease vaccine production. Vaccine 2019, 37, 6380–6389. [Google Scholar] [CrossRef]
- Genzel, Y.; Vogel, T.; Buck, J.; Behrendt, I.; Ramirez, D.V.; Schiedner, G.; Jordan, I.; Reichl, U. High cell density cultivations by alternating tangential flow (ATF) perfusion for influenza A virus production using suspension cells. Vaccine 2014, 32, 2770–2781. [Google Scholar] [CrossRef]
- Kim, A.Y.; Kim, H.; Park, S.Y.; Park, S.H.; Lee, J.M.; Kim, J.S.; Park, J.W.; Park, C.K.; Park, J.H.; Ko, Y.J. Investigation of the optimal medium and application strategy for foot-and-mouth disease vaccine antigen production. J. Appl. Microbiol. 2021. [Google Scholar] [CrossRef]
- Barteling, S.J.; Meloen, R.H. A simple method for the quantification of 140 s particles of foot-and-mouth disease virus (FMDV). Archiv für die gesamte Virusforschung 1974, 45, 362–364. [Google Scholar] [CrossRef]
- Aarthi, D.; Ananda Rao, K.; Robinson, R.; Srinivasan, V.A. Validation of binary ethyleneimine (BEI) used as an inactivant for foot and mouth disease tissue culture vaccine. Biologicals 2004, 32, 153–156. [Google Scholar] [CrossRef] [PubMed]
- Doel, T.R. FMD vaccines. Virus Res. 2003, 91, 19. [Google Scholar] [CrossRef]
- Park, J.-N.; Lee, S.-Y.; Chu, J.-Q.; Lee, Y.-J.; Kim, R.-H.; Lee, K.-N.; Kim, S.-M.; Tark, D.-S.; Kim, B.; Park, J.-H.J.V. Protection to homologous and heterologous challenge in pigs immunized with vaccine against foot-and-mouth disease type O caused an epidemic in East Asia during 2010/2011. Vaccine 2014, 32, 1882–1889. [Google Scholar] [CrossRef] [PubMed]
- Lyons, N.A.; Lyoo, Y.S.; King, D.P.; Paton, D.J. Challenges of generating and maintaining protective vaccine-induced immune responses for foot-and-mouth disease virus in pigs. Front. Vet. Sci. 2016, 3, 102. [Google Scholar] [CrossRef]
- Ko, E.Y.; Jung, S.; Jeong, H.K.; Han, J.H.; Son, J.H. Effects of foot-and-mouth disease vaccination location and injection device on the incidence of site lesions in pork. Korean J. Food Sci. Anim. Resour. 2018, 38, 498–505. [Google Scholar] [CrossRef]
- Park, M.-E.; You, S.-H.; Lee, S.-Y.; Lee, K.-N.; Ko, M.-K.; Choi, J.-H.; Kim, B.; Lee, J.-S.; Park, J.-H. Immune responses in pigs and cattle vaccinated with half-volume foot-and-mouth disease vaccine. J. Vet. Sci. 2017, 18, 323–331. [Google Scholar] [CrossRef]
- Dekker, A.; Sanz-Bernardo, B.; Singanallur, N.B.; Ludi, A.B.; Horsington, J.; Eble, P.L.; King, D.P.; Vosloo, W. Cross-protection induced by a A/MAY/97 emergency vaccine against intra-serotype heterologous challenge with a foot-and-mouth disease virus from the A/ASIA/G-VII lineage. Vaccines 2020, 8, 24. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Pig ID | Virus Neutralizing Antibody Titers (log10) | Clinical Score | Protection | ||||
|---|---|---|---|---|---|---|---|---|
| 0 dpv | 7 dpv | 14 dpv | 21 dpv | 28 dpv | ||||
| Full dose | #3 | <0.9 | 1.20 | 1.81 | 1.95 | 1.95 | 1 * | Yes a |
| #4 | <0.9 | 1.04 | 1.51 | 1.51 | 1.95 | 0 | Yes | |
| #5 | <0.9 | 1.04 | 1.04 | 1.95 | 1.95 | 0 | Yes | |
| #6 | <0.9 | 0.9 | 1.34 | 1.51 | 1.51 | 0 | Yes | |
| #7 | <0.9 | 1.34 | 1.04 | 1.95 | 2.41 | 0 | Yes | |
| 1/3 dose | #10 | <0.9 | 1.20 | <0.9 | <0.9 | 1.34 | 0 | Yes |
| #11 | <0.9 | 1.04 | 1.34 | 1.51 | 1.34 | 0 | Yes | |
| #13 | <0.9 | <0.9 | <0.9 | 0.9 | 1.2 | 0 | Yes | |
| #14 | <0.9 | 1.34 | 1.20 | 0.9 | 1.04 | 0 | Yes | |
| #15 | <0.9 | <0.9 | <0.9 | 1.2 | 1.04 | 0 | Yes | |
| 1/9 dose | #18 | <0.9 | 1.34 | 1.04 | 1.04 | 1.04 | 9 | No |
| #19 | <0.9 | <0.9 | <0.9 | <0.9 | 1.04 | 0 | Yes | |
| #20 | <0.9 | 0.9 | <0.9 | 1.04 | 1.51 | 0 | Yes | |
| #24 | <0.9 | 1.2 | 1.04 | 1.20 | 0.9 | 0 | Yes | |
| #25 | <0.9 | <0.9 | <0.9 | <0.9 | <0.9 | 0 | Yes | |
| Control | #26 | <0.9 | <0.9 | <0.9 | <0.9 | <0.9 | 10 | No |
| #27 | <0.9 | <0.9 | <0.9 | <0.9 | <0.9 | 13 | No | |
| #30 | <0.9 | <0.9 | <0.9 | <0.9 | <0.9 | 12 | No | |
| Groups | Pig ID | Virus Neutralizing Antibody Titers (log10) | Clinical Score | Protection | ||||
|---|---|---|---|---|---|---|---|---|
| 0 dpv | 7 dpv | 14 dpv | 21 dpv | 28 dpv | ||||
| Full dose | #2 | <0.9 | 1.51 | 2.41 | 2.71 | 2.86 | 0 | Yes |
| #4 | <0.9 | 1.20 | 2.26 | 2.11 | 3.01 | 0 | Yes | |
| #7 | <0.9 | 1.20 | 2.56 | 2.41 | 2.56 | 0 | Yes | |
| #17 | <0.9 | 1.51 | 2.41 | 2.41 | 2.71 | 0 | Yes | |
| #25 | <0.9 | 1.34 | 1.51 | 2.56 | 2.26 | 0 | Yes | |
| 1/3 dose | #5 | <0.9 | 1.20 | 2.26 | 2.26 | 2.11 | 0 | Yes |
| #9 | <0.9 | 1.51 | 1.95 | 1.81 | 2.26 | 0 | Yes | |
| #18 | <0.9 | 1.34 | 2.11 | 1.81 | 1.81 | 0 | Yes | |
| #22 | <0.9 | 1.34 | 2.26 | 1.65 | 1.81 | 0 | Yes | |
| #27 | <0.9 | 1.04 | 1.95 | 2.26 | 1.65 | 0 | Yes | |
| 1/9 dose | #12 | <0.9 | <0.9 | 1.51 | 2.26 | 1.81 | 0 | Yes |
| #14 | <0.9 | 1.34 | 1.81 | 1.65 | 1.65 | 0 | Yes | |
| #26 | <0.9 | 1.34 | 1.34 | 1.51 | 1.34 | 0 | Yes | |
| #28 | <0.9 | 1.04 | 1.65 | 1.81 | 1.65 | 0 | Yes | |
| #29 | <0.9 | 1.04 | 1.20 | 1.20 | 1.34 | 0 | Yes | |
| Control | #20 | <0.9 | 0.9 | <0.9 | <0.9 | <0.9 | 11 | No |
| #21 | 0.9 | <0.9 | <0.9 | <0.9 | <0.9 | 10 | No | |
| #30 | 1.2 | 1.04 | 0.9 | <0.9 | <0.9 | 12 | No | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, S.-H.; Lee, S.-Y.; Kim, J.-S.; Kim, A.-Y.; Park, S.-Y.; Lee, J.-H.; Lee, M.; Kim, H.; Lee, S.-I.; Kang, N.-Y.; et al. Scale-Up Production of Type O and A Foot-and-Mouth Disease Bivalent Vaccine and Its Protective Efficacy in Pigs. Vaccines 2021, 9, 586. https://doi.org/10.3390/vaccines9060586
Park S-H, Lee S-Y, Kim J-S, Kim A-Y, Park S-Y, Lee J-H, Lee M, Kim H, Lee S-I, Kang N-Y, et al. Scale-Up Production of Type O and A Foot-and-Mouth Disease Bivalent Vaccine and Its Protective Efficacy in Pigs. Vaccines. 2021; 9(6):586. https://doi.org/10.3390/vaccines9060586
Chicago/Turabian StylePark, Sang-Hyun, Seo-Yong Lee, Jae-Seok Kim, Ah-Young Kim, Sun-Young Park, Ji-Hye Lee, Mijung Lee, Hyejin Kim, Sim-In Lee, Na-Young Kang, and et al. 2021. "Scale-Up Production of Type O and A Foot-and-Mouth Disease Bivalent Vaccine and Its Protective Efficacy in Pigs" Vaccines 9, no. 6: 586. https://doi.org/10.3390/vaccines9060586
APA StylePark, S.-H., Lee, S.-Y., Kim, J.-S., Kim, A.-Y., Park, S.-Y., Lee, J.-H., Lee, M., Kim, H., Lee, S.-I., Kang, N.-Y., Park, J.-W., Kim, S.-M., Park, J.-H., & Ko, Y.-J. (2021). Scale-Up Production of Type O and A Foot-and-Mouth Disease Bivalent Vaccine and Its Protective Efficacy in Pigs. Vaccines, 9(6), 586. https://doi.org/10.3390/vaccines9060586