
The Immune Correlates of Orthohantavirus Vaccine

 , ,
, ,
Abstract
1. Introduction
2. Antigenicity of Orthohantavirus Vaccine
2.1. Clinical Studies for Hantavax Vaccine
2.2. Evaluation of Hantavax Vaccine Effectiveness
2.3. Antigenicity of Orthohantaviruses
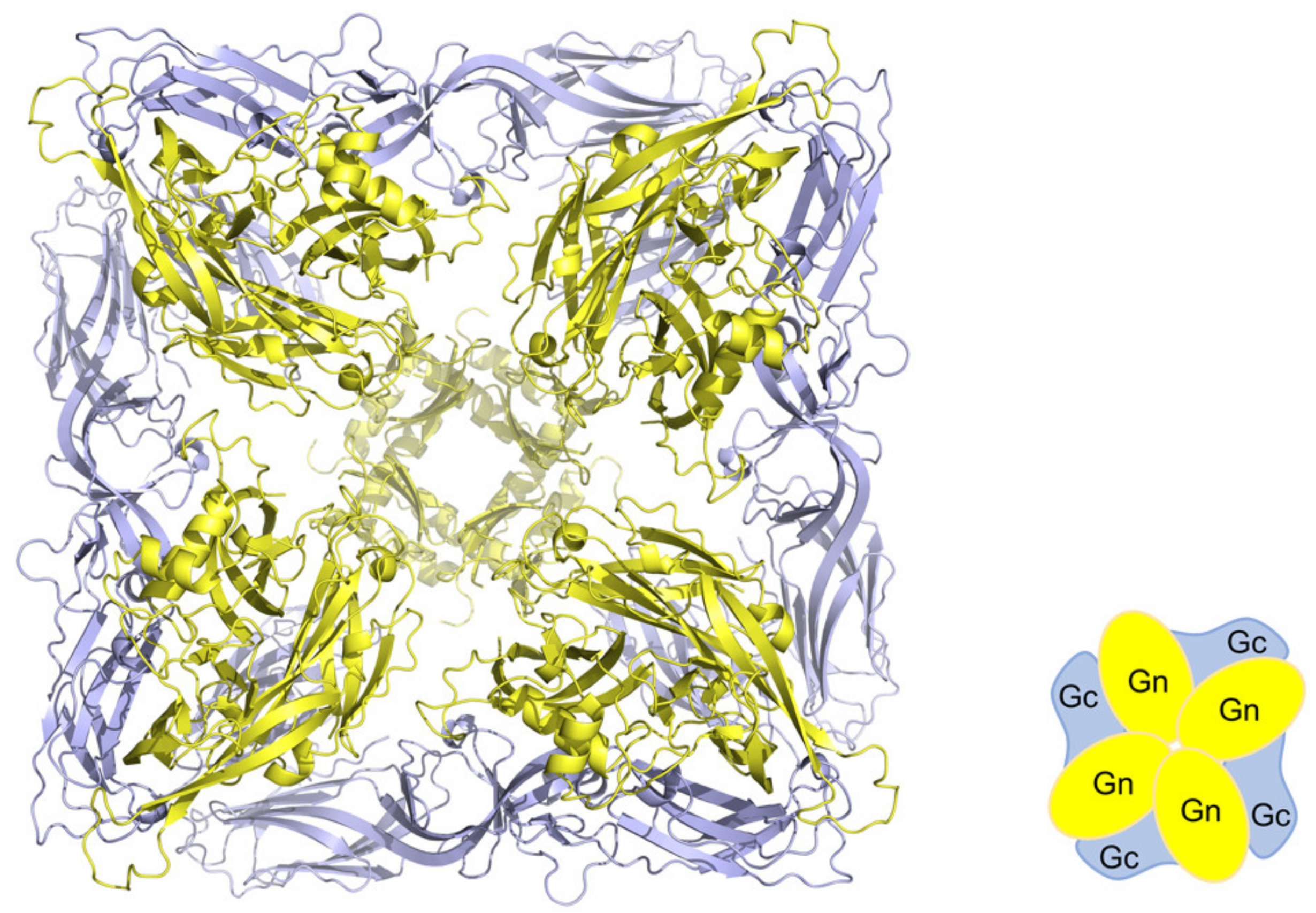
2.4. Antigenicity of Glycoproteins
2.5. Antigenicity of the Nucleocapsid Protein
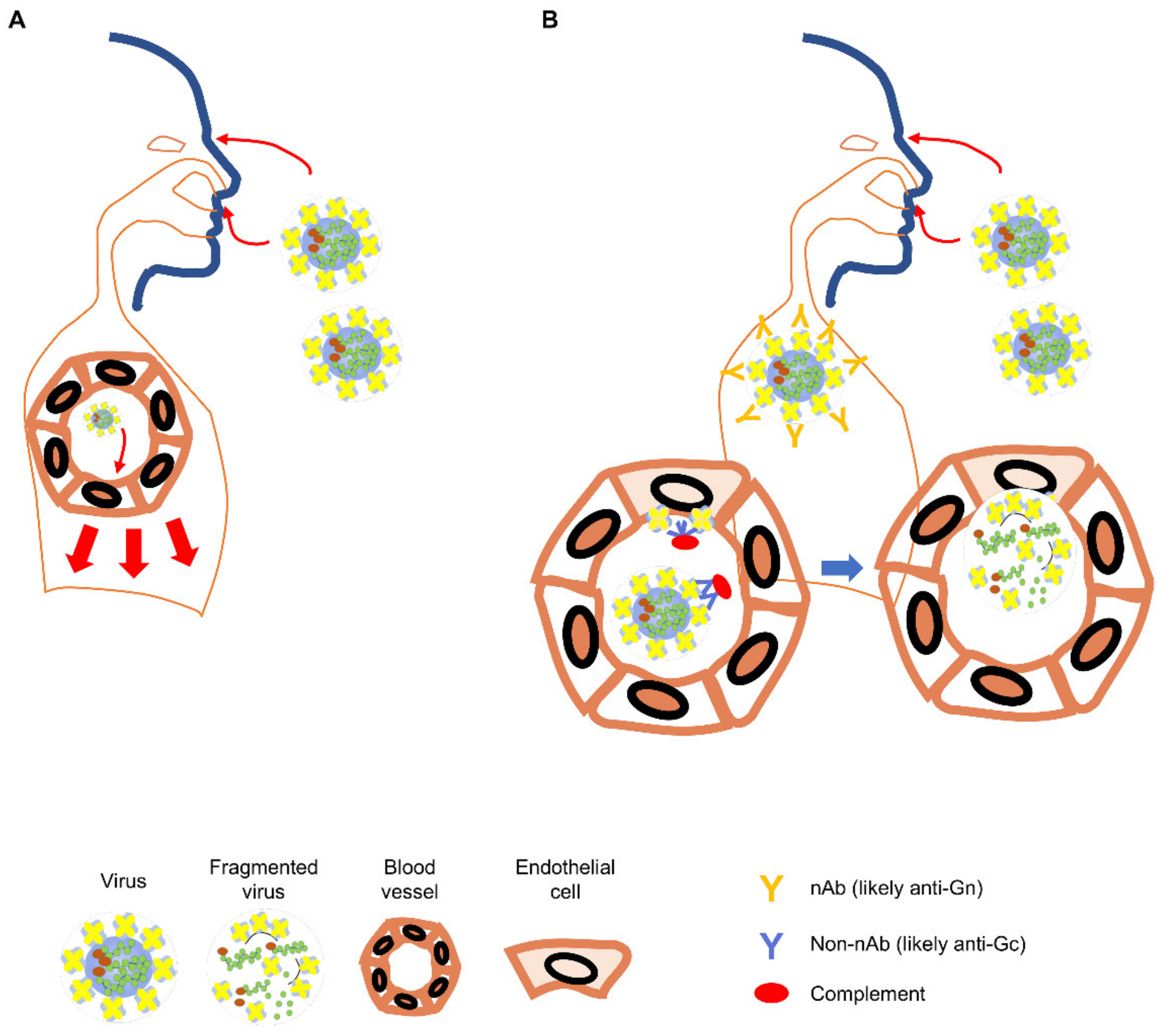
2.6. Role for Non-Neutralizing Antibody Responses
2.7. Strategy to Design Orthohantavirus Vaccines as Pandemic Preparedness
3. Conclusions and Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Elliott, R.M.; Schmaljohn, C.S. Bunyaviridae. In Fields Virology, 6th ed.; Knipe, D.M., Howley, P.M., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2013; pp. 1244–1282. [Google Scholar]
- Lee, H.W.; Lee, P.W.; Johnson, K.M. Isolation of the Etiologic Agent of Korean Hemorrhagic Fever. J. Infect. Dis. 1978, 137, 298–308. [Google Scholar] [CrossRef]
- Lee, H.W. Hemorrhagic fever with renal syndrome in Korea. Rev. Infect. Dis. 1989, 11 (Suppl. 4), S864–S876. [Google Scholar] [CrossRef] [PubMed]
- Schmaljohn, C.S.; Hasty, S.E.; Dalrymple, J.M.; le Duc, J.W.; Lee, H.W.; von Bonsdorff, C.H.; Brummer-Korvenkontio, M.; Vaheri, A.; Tsai, T.F.; Regnery, H.L.; et al. Antigenic and genetic properties of viruses linked to hemorrhagic fever with renal syndrome. Science 1985, 227, 1041–1044. [Google Scholar] [CrossRef]
- Lee, H.W.; Vaheri, A.; Schmaljohn, C.S. Discovery of hantaviruses and of the Hantavirus genus: Personal and historical perspectives of the Presidents of the International Society of Hantaviruses. Virus Res. 2014, 187, 2–5. [Google Scholar] [CrossRef]
- Clement, J.; Maes, P.; Lagrou, K.; van Ranst, M.; Lameire, N. A unifying hypothesis and a single name for a complex globally emerging infection: Hantavirus disease. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Yanagihara, R.; Gu, S.H.; Arai, S.; Kang, H.J.; Song, J.-W. Hantaviruses: Rediscovery and new beginnings. Virus Res. 2014, 187, 6–14. [Google Scholar] [CrossRef]
- Song, J.-W.; Kang, H.J.; Gu, S.H.; Moon, S.S.; Bennett, S.N.; Song, K.-J.; Baek, L.J.; Kim, H.-C.; O’Guinn, M.L.; Chong, S.-T.; et al. Characterization of Imjin Virus, a Newly Isolated Hantavirus from the Ussuri White-Toothed Shrew (Crocidura lasiura). J. Virol. 2009, 83, 6184–6191. [Google Scholar] [CrossRef] [PubMed]
- Golden, J.W.; Hammerbeck, C.D.; Mucker, E.M.; Brocato, R.L. Animal Models for the Study of Rodent-Borne Hemorrhagic Fever Viruses: Arenaviruses and Hantaviruses. BioMed Res. Int. 2015, 2015, 793257. [Google Scholar] [CrossRef] [PubMed]
- Schountz, T.; Prescott, J. Hantavirus Immunology of Rodent Reservoirs: Current Status and Future Directions. Viruses 2014, 6, 1317–1335. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.-D.; Guo, W.-P.; Wang, W.; Zou, Y.; Hao, Z.-Y.; Zhou, D.-J.; Dong, X.; Qu, Y.-G.; Li, M.-H.; Tian, H.-F.; et al. Migration of Norway Rats Resulted in the Worldwide Distribution of Seoul Hantavirus Today. J. Virol. 2011, 86, 972–981. [Google Scholar] [CrossRef] [PubMed]
- Plyusnina, A.; Heyman, P.; Baert, K.; Stuyck, J.; Cochez, C.; Plyusnin, A. Genetic characterization of seoul hantavirus originated from norway rats (Rattus norvegicus) captured in Belgium. J. Med. Virol. 2012, 84, 1298–1303. [Google Scholar] [CrossRef]
- Swanink, C.; Reimerink, J.; Gisolf, J.; de Vries, A.; Claassen, M.; Martens, L.; Waegemaekers, T.; Rozendaal, H.; Valkenburgh, S.; Hoornweg, T.; et al. Autochthonous Human Case of Seoul Virus Infection, The Netherlands. Emerg. Infect. Dis. 2018, 24, 2158–2163. [Google Scholar] [CrossRef]
- Miller, M.; Hagan, E. Integrated biological–behavioural surveillance in pandemic-threat warning systems. Bull. World Health Organ. 2016, 95, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Brocato, R.L.; Hooper, J.W. Progress on the Prevention and Treatment of Hantavirus Disease. Viruses 2019, 11, 610. [Google Scholar] [CrossRef]
- Martinez-Valdebenito, C.; Calvo, M.; Vial, C.; Mansilla, R.; Marco, C.; Palma, R.E.; Vial, P.A.; Valdivieso, F.; Mertz, G.; Ferrés, M. Person-to-Person Household and Nosocomial Transmission of Andes Hantavirus, Southern Chile, 2011. Emerg. Infect. Dis. 2014, 20, 1629–1636. [Google Scholar] [CrossRef]
- Martinez, V.P.; Bellomo, C.; san Juan, J.S.; Pinna, D.; Forlenza, R.; Elder, M.; Padula, P.J. Person-to-Person Transmission of Andes Virus. Emerg. Infect. Dis. 2005, 11, 1848–1853. [Google Scholar] [CrossRef] [PubMed]
- Ferrés, M.; Vial, P.; Marco, C.; Yañez, L.; Godoy, P.; Castillo, C.; Hjelle, B.; Delgado, I.; Lee, S.; Mertz, G.J.; et al. Prospective Evaluation of Household Contacts of Persons with Hantavirus Cardiopulmonary Syndrome in Chile. J. Infect. Dis. 2007, 195, 1563–1571. [Google Scholar] [CrossRef] [PubMed]
- Schmaljohn, C. Vaccines for hantaviruses. Vaccine 2009, 27 (Suppl. 4), D61–D64. [Google Scholar] [CrossRef]
- Park, M.S.; Kim, J.I.; Bae, J.-Y.; Park, M.-S. Animal models for the risk assessment of viral pandemic potential. Lab. Anim. Res. 2020, 36, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Dalrymple, N.A.; Mackow, E.R. Virus interactions with endothelial cell receptors: Implications for viral pathogenesis. Curr. Opin. Virol. 2014, 7, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Bostik, P.; Winter, J.; Ksiazek, T.G.; Rollin, P.E.; Villinger, F.; Zaki, S.R.; Peters, C.J.; Ansari, A.A. Sin Nombre Virus (SNV) Ig Isotype Antibody Response during Acute and Convalescent Phases of Hantavirus Pulmonary Syndrome. Emerg. Infect. Dis. 2000, 6, 184–188. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.-S.; Kariwa, H.; Takashima, I.; Yoshimatsu, K.; Arikawa, J.; Hashimoto, N. Antibody-dependent enhancement of hantavirus infection in macrophage cell lines. Arch. Virol. 1992, 122, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Ghebrehiwet, B.; Kaplan, A.P.; Joseph, K.; Peerschke, E.I.B. The complement and contact activation systems: Partnership in pathogenesis beyond angioedema. Immunol. Rev. 2016, 274, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.; Kwon, Y.-C.; Kim, S.-I.; Park, J.-M.; Lee, K.-H.; Ahn, B.-Y. A hantavirus causing hemorrhagic fever with renal syndrome requires gC1qR/p32 for efficient cell binding and infection. J. Virol. 2008, 381, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Schountz, T.; Quackenbush, S.; Rovnak, J.; Haddock, E.; Black, W.C.; Feldmann, H.; Prescott, J. Differential Lymphocyte and Antibody Responses in Deer Mice Infected with Sin Nombre Hantavirus or Andes Hantavirus. J. Virol. 2014, 88, 8319–8331. [Google Scholar] [CrossRef]
- Spengler, J.R.; Haddock, E.; Gardner, D.; Hjelle, B.; Feldmann, H.; Prescott, J. Experimental Andes Virus Infection in Deer Mice: Characteristics of Infection and Clearance in a Heterologous Rodent Host. PLoS ONE 2013, 8, e55310. [Google Scholar] [CrossRef]
- Zheng, Y.; Zhou, B.-Y.; Wei, J.; Xu, Y.; Dong, J.-H.; Guan, L.-Y.; Ma, P.; Yu, P.-B.; Wang, J.-J. Persistence of immune responses to vaccine against haemorrhagic fever with renal syndrome in healthy adults aged 16–60 years: Results from an open-label2-year follow-up study. Infect. Dis. 2018, 50, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Ma, H.; Shu, J.; Zhang, Q.; Han, M.; Liu, Z.; Jin, X.; Zhang, F.; Wu, X. Vaccines and Therapeutics Against Hantaviruses. Front. Microbiol. 2019, 10, 2989. [Google Scholar] [CrossRef]
- Cho, H.-W.; Howard, C.R.; Lee, H.-W. Review of an Inactivated Vaccine against Hantaviruses. Intervirology 2002, 45, 328–333. [Google Scholar] [CrossRef]
- Jung, J.; Ko, S.-J.; Oh, H.S.; Moon, S.M.; Song, J.-W.; Huh, K. Protective Effectiveness of Inactivated Hantavirus Vaccine Against Hemorrhagic Fever with Renal Syndrome. J. Infect. Dis. 2018, 217, 1417–1420. [Google Scholar] [CrossRef]
- Jonsson, C.B.; Figueiredo, L.T.M.; Vapalahti, O. A Global Perspective on Hantavirus Ecology, Epidemiology, and Disease. Clin. Microbiol. Rev. 2010, 23, 412–441. [Google Scholar] [CrossRef]
- Cho, H.-W.; Howard, C.R. Antibody responses in humans to an inactivated hantavirus vaccine (Hantavax®). Vaccine 1999, 17, 2569–2575. [Google Scholar] [CrossRef]
- Sohn, Y.M.; Rho, H.O.; Park, M.S.; Kim, J.S.; Summers, P.L. Primary Humoral Immune Responses to Formalin Inactivated Hemorrhagic Fever with Renal Syndrome Vaccine (Hantavax®): Consideration of Active Immunization in South Korea. Yonsei Med. J. 2001, 42, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Song, J.Y.; Woo, H.J.; Cheong, H.J.; Noh, J.Y.; Baek, L.J.; Kim, W.J. Long-term immunogenicity and safety of inactivated Hantaan virus vaccine (Hantavax™) in healthy adults. Vaccine 2016, 34, 1289–1295. [Google Scholar] [CrossRef] [PubMed]
- Song, J.Y.; Jeong, H.W.; Yun, J.W.; Lee, J.; Woo, H.J.; Bae, J.-Y.; Park, M.-S.; Choi, W.S.; Park, D.W.; Noh, J.Y.; et al. Immunogenicity and safety of a modified three-dose priming and booster schedule for the Hantaan virus vaccine (Hantavax): A multi-center phase III clinical trial in healthy adults. Vaccine 2020, 38, 8016–8023. [Google Scholar] [CrossRef] [PubMed]
- Randolph, H.E.; Barreiro, L.B. Herd Immunity: Understanding COVID-19. Immunity 2020, 52, 737–741. [Google Scholar] [CrossRef]
- Park, K.; Kim, C.S.; Moon, K.-T. Protective Effectiveness of Hantavirus Vaccine. Emerg. Infect. Dis. 2004, 10, 2218–2220. [Google Scholar] [CrossRef]
- Choi, Y.; Ahn, C.-J.; Seong, K.-M.; Jung, M.-Y.; Ahn, B.-Y. Inactivated Hantaan virus vaccine derived from suspension culture of Vero cells. Vaccine 2003, 21, 1867–1873. [Google Scholar] [CrossRef]
- Elgh, F.; Lundkvist, A.; Alexeyev, O.A.; Stenlund, H.; Avsic-Zupanc, T.; Hjelle, B.; Lee, H.W.; Smith, K.J.; Vainionpää, R.; Wiger, D.; et al. Serological diagnosis of hantavirus infections by an enzyme-linked immunosorbent assay based on detection of immunoglobulin G and M responses to recombinant nucleocapsid proteins of five viral serotypes. J. Clin. Microbiol. 1997, 35, 1122–1130. [Google Scholar] [CrossRef] [PubMed]
- Niklasson, B.; Lundkvist, Å.; Hörling, J. The humoral response to Puumala virus infection (Nephropathia epidemica) investigated by viral protein specific immunoassays. Arch. Virol. 1993, 130, 121–130. [Google Scholar] [CrossRef]
- De Carvalho, N.C.D.C.; Björling, E.; Lundkvist, Å. Immunoglobulin A responses to Puumala hantavirus. J. Gen. Virol. 2000, 81, 1453–1461. [Google Scholar] [CrossRef]
- Lundkvist, A.; Niklasson, B. Bank vole monoclonal antibodies against Puumala virus envelope glycoproteins: Identification of epitopes involved in neutralization. Arch. Virol. 1992, 126, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Valdivieso, F.; Vial, P.; Ferres, M.; Ye, C.; Goade, D.; Cuiza, A.; Hjelle, B. Neutralizing Antibodies in Survivors of Sin Nombre and Andes Hantavirus Infection. Emerg. Infect. Dis. 2006, 12, 166–168. [Google Scholar] [CrossRef] [PubMed]
- Rissanen, I.; Stass, R.; Zeltina, A.; Li, S.; Hepojoki, J.; Harlos, K.; Gilbert, R.J.C.; Huiskonen, J.T.; Bowden, T.A. Structural Transitions of the Conserved and Metastable Hantaviral Glycoprotein Envelope. J. Virol. 2017, 91, 91. [Google Scholar] [CrossRef]
- Serris, A.; Stass, R.; Bignon, E.A.; Muena, N.A.; Manuguerra, J.-C.; Jangra, R.K.; Li, S.; Chandran, K.; Tischler, N.D.; Huiskonen, J.T.; et al. The Hantavirus Surface Glycoprotein Lattice and Its Fusion Control Mechanism. Cell 2020, 183, 442–456.e16. [Google Scholar] [CrossRef] [PubMed]
- Krarup, A.; Truan, D.; Furmanova-Hollenstein, P.; Bogaert, L.; Bouchier, P.; Bisschop, I.J.M.; Widjojoatmodjo, M.N.; Zahn, R.; Schuitemaker, H.; McLellan, J.S.; et al. A highly stable prefusion RSV F vaccine derived from structural analysis of the fusion mechanism. Nat. Commun. 2015, 6, 8143. [Google Scholar] [CrossRef]
- Elgh, F.; Lundkvist, Å.; Alexeyev, O.A.; Wadell, G.; Juto, P. A major antigenic domain for the human humoral response to Puumala virus nucleocapsid protein is located at the aminoterminus. J. Virol. Methods 1996, 59, 161–172. [Google Scholar] [CrossRef]
- Kang, J.-I.; Lee, Y.-S.; Ahn, K.; Song, J.-W.; Song, K.-J.; Baek, L.J.; Lee, P.-W.; Ahn, B.-Y. A dominant antigenic region of the hantaan virus nucleocapsid protein is located within a amino-terminal short stretch of hydrophilic residues. Virus Genes 2001, 23, 183–186. [Google Scholar] [CrossRef]
- Safronetz, D.; Hegde, N.R.; Ebihara, H.; Denton, M.; Kobinger, G.P.; Jeor, S.S.; Feldmann, H.; Johnson, D.C. Adenovirus Vectors Expressing Hantavirus Proteins Protect Hamsters against Lethal Challenge with Andes Virus. J. Virol. 2009, 83, 7285–7295. [Google Scholar] [CrossRef]
- De Carvalho, N.C.D.C.; Gonzalez, D.; Della-Valle, M.G.; Padula, P.; Bjoörling, E.; Plyusnin, A.; Lundkvist, A. Cross-Protection against Challenge with Puumala Virus after Immunization with Nucleocapsid Proteins from Different Hantaviruses. J. Virol. 2002, 76, 6669–6677. [Google Scholar] [CrossRef]
- Maes, P.; Keyaerts, E.; Bonnet, V.; Clement, J.; Avsic-Zupanc, T.; Robert, A.; van Ranst, M. Truncated Recombinant Dobrava Hantavirus Nucleocapsid Proteins Induce Strong, Long-Lasting Immune Responses in Mice. Intervirology 2006, 49, 253–260. [Google Scholar] [CrossRef]
- Auladell, M.; Jia, X.; Hensen, L.; Chua, B.; Fox, A.; Nguyen, T.H.O.; Doherty, P.C.; Kedzierska, K. Recalling the Future: Immunological Memory Toward Unpredictable Influenza Viruses. Front. Immunol. 2019, 10, 1400. [Google Scholar] [CrossRef] [PubMed]
- St. John, A.L.S.; Rathore, A.P.S. Adaptive immune responses to primary and secondary dengue virus infections. Nat. Rev. Immunol. 2019, 19, 218–230. [Google Scholar] [CrossRef] [PubMed]
- Phan, T.G.; Tangye, S.G. Memory B cells: Total recall. Curr. Opin. Immunol. 2017, 45, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Youngblood, B.; Hale, J.S.; Ahmed, R. T-cell memory differentiation: Insights from transcriptional signatures and epigenetics. Immunology 2013, 139, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Fazilleau, N.; Mark, L.; McHeyzer-Williams, L.J.; McHeyzer-Williams, M.G. Follicular Helper T Cells: Lineage and Location. J. Immun. 2009, 30, 324–335. [Google Scholar] [CrossRef]
- Colbert, J.D.; Cruz, F.M.; Rock, K.L. Cross-presentation of exogenous antigens on MHC I molecules. Curr. Opin. Immunol. 2020, 64, 1–8. [Google Scholar] [CrossRef]
- Getahun, A.; Heyman, B. How antibodies act as natural adjuvants. Immunol. Lett. 2006, 104, 38–45. [Google Scholar] [CrossRef]
- Heyman, B. Antibodies as Natural Adjuvants. Curr. Top. Microbiol. Immunol. 2014, 382, 201–219. [Google Scholar] [CrossRef]
- Mackow, E.R.; Gavrilovskaya, I.N. Cellular Receptors and Hantavirus Pathogenesis. Curr. Top. Microbiol. Immunol. 2001, 256, 91–115. [Google Scholar] [CrossRef] [PubMed]
- Harris, A.; Cardone, G.; Winkler, D.C.; Heymann, J.B.; Brecher, M.; White, J.M.; Steven, A.C. Influenza virus pleiomorphy characterized by cryoelectron tomography. Proc. Natl. Acad. Sci. USA 2006, 103, 19123–19127. [Google Scholar] [CrossRef]
- Yamanaka, K.; Ishihama, A.; Nagata, K. Reconstitution of influenza virus RNA-nucleoprotein complexes structurally resembling native viral ribonucleoprotein cores. J. Biol. Chem. 1990, 265, 11151–11155. [Google Scholar] [CrossRef]
- Wang, M.; Wang, J.; Zhu, Y.; Xu, Z.; Yang, K.; Yang, A.; Jin, B. Cellular Immune Response to Hantaan Virus Nucleocapsid Protein in the Acute Phase of Hemorrhagic Fever with Renal Syndrome: Correlation with Disease Severity. J. Infect. Dis. 2009, 199, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Bucht, G.; Sjölander, K.B.; Eriksson, S.; Lindgren, L.; Lundkvist, Å.; Elgh, F. Modifying the cellular transport of DNA-based vaccines alters the immune response to hantavirus nucleocapsid protein. Vaccine 2001, 19, 3820–3829. [Google Scholar] [CrossRef]
- Sohn, Y.; Jeong, S.J.; Chung, W.S.; Hyun, J.H.; Baek, Y.J.; Cho, Y.; Kim, J.H.; Ahn, J.Y.; Choi, J.Y.; Yeom, J.-S. Assessing Viral Shedding and Infectivity of Asymptomatic or Mildly Symptomatic Patients with COVID-19 in a Later Phase. J. Clin. Med. 2020, 9, 2924. [Google Scholar] [CrossRef] [PubMed]
- Molenaar-de Backer, M.M.-D.; Russcher, A.; Kroes, A.; Koppelman, M.; Lanfermeijer, M.; Zaaijer, H. Detection of parvovirus B19 DNA in blood: Viruses or DNA remnants? J. Clin. Virol. 2016, 84, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Sheshberadaran, H.; Niklasson, B.; Tkachenko, E.A. Antigenic Relationship between Hantaviruses Analysed by Immunoprecipitation. J. Gen. Virol. 1988, 69, 2645–2651. [Google Scholar] [CrossRef]
- Schubert, J.; Tollmann, F.; Weissbrich, B. Evaluation of a pan-reactive hantavirus enzyme immunoassay and of a hantavirus immunoblot for the diagnosis of Nephropathia epidemica. J. Clin. Virol. 2001, 21, 63–74. [Google Scholar] [CrossRef]
- Araki, K.; Yoshimatsu, K.; Ogino, M.; Ebihara, H.; Lundkvist, Å.; Kariwa, H.; Takashima, I.; Arikawa, J. Truncated Hantavirus Nucleocapsid Proteins for Serotyping Hantaan, Seoul, and Dobrava Hantavirus Infections. J. Clin. Microbiol. 2001, 39, 2397–2404. [Google Scholar] [CrossRef]
- Schmidt, J.; Meisel, H.; Capria, S.G.; Petraityte, R.; Lundkvist, Å.; Hjelle, B.; Vial, P.A.; Padula, P.; Krüger, D.H.; Ulrich, R. Serological Assays for the Detection of Human Andes Hantavirus Infections Based on Its Yeast-Expressed Nucleocapsid Protein. Intervirology 2006, 49, 173–184. [Google Scholar] [CrossRef] [PubMed]
- Lindkvist, M.; Näslund, J.; Ahlm, C.; Bucht, G. Cross-reactive and serospecific epitopes of nucleocapsid proteins of three hantaviruses: Prospects for new diagnostic tools. Virus Res. 2008, 137, 97–105. [Google Scholar] [CrossRef]
- Hooper, J.W.; Custer, D.M.; Thompson, E.; Schmaljohn, C.S. DNA Vaccination with the Hantaan Virus M Gene Protects Hamsters against Three of Four HFRS Hantaviruses and Elicits a High-Titer Neutralizing Antibody Response in Rhesus Monkeys. J. Virol. 2001, 75, 8469–8477. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.W.; Baek, L.J.; Johnson, K.M. Isolation of Hantaan Virus, the Etiologic Agent of Korean Hemorrhagic Fever, from Wild Urban Rats. J. Infect. Dis. 1982, 146, 638–644. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.W.; Gibbs, C.J., Jr.; Gajdusek, D.C.; Yanagihara, R. Serotypic classification of hantaviruses by indirect immunofluorescent antibody and plaque reduction neutralization tests. J. Clin. Microbiol. 1985, 22, 940–944. [Google Scholar] [CrossRef] [PubMed]
- Hooper, J.W.; Custer, D.M.; Smith, J.; Wahl-Jensen, V. Hantaan/Andes virus DNA vaccine elicits a broadly cross-reactive neutralizing antibody response in nonhuman primates. J. Virol. 2006, 347, 208–216. [Google Scholar] [CrossRef]
- Hooper, J.W.; Josleyn, M.; Ballantyne, J.; Brocato, R. A novel Sin Nombre virus DNA vaccine and its inclusion in a candidate pan-hantavirus vaccine against hantavirus pulmonary syndrome (HPS) and hemorrhagic fever with renal syndrome (HFRS). J. Vaccine 2013, 31, 4314–4321. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hooper, J.W.; Moon, J.E.; Paolino, K.M.; Newcomer, R.; McLain, D.E.; Josleyn, M.; Hannaman, D.; Schmaljohn, C. A Phase 1 clinical trial of Hantaan virus and Puumala virus M-segment DNA vaccines for haemorrhagic fever with renal syndrome delivered by intramuscular electroporation. Clin. Microbiol. Infect. 2014, 20 (Suppl. 5), 110–117. [Google Scholar] [CrossRef] [PubMed]
- Caddy, S.L.; Vaysburd, M.; Wing, M.; Foss, S.; Andersen, J.T.; O‘Connell, K.; Mayes, K.; Higginson, K.; Iturriza-Gómara, M.; Desselberger, U.; et al. Intracellular neutralisation of rotavirus by VP6-specific IgG. PLoS Pathog. 2020, 16, e1008732. [Google Scholar] [CrossRef] [PubMed]
- Carragher, D.M.; Kaminski, D.A.; Moquin, A.; Hartson, L.; Randall, T.D. A Novel Role for Non-Neutralizing Antibodies against Nucleoprotein in Facilitating Resistance to Influenza Virus. J. Immunol. 2008, 181, 4168–4176. [Google Scholar] [CrossRef] [PubMed]
- Leifer, C.A.; Medvedev, A.E. Molecular mechanisms of regulation of Toll-like receptor signaling. J. Leukoc. Biol. 2016, 100, 927–941. [Google Scholar] [CrossRef]
- Shaw, M.; Palese, P. Chapter 40. Orthomyxoviridae. In Fields Virology, 6th ed.; Knipe, D., Howley, P., Eds.; Lippincott Williams & Wilkins: Philadephia, PA, USA, 2013; pp. 1151–1185. [Google Scholar]
- Kim, J.I.; Lee, I.; Park, S.; Hwang, M.-W.; Bae, J.-Y.; Lee, S.; Heo, J.; Park, M.S.; García-Sastre, A.; Park, M.S. Genetic Requirement for Hemagglutinin Glycosylation and Its Implications for Influenza A H1N1 Virus Evolution. J. Virol. 2013, 87, 7539–7549. [Google Scholar] [CrossRef] [PubMed]
- Park, M.S.; Kim, J.I.; Park, S.; Lee, I.; Park, M.-S. Original Antigenic Sin Response to RNA Viruses and Antiviral Immunity. Immune Netw. 2016, 16, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Lillie, P.J.; Berthoud, T.K.; Powell, T.J.; Lambe, T.; Mullarkey, C.E.; Spencer, A.J.; Hamill, M.; Peng, Y.; Blais, M.-E.; Duncan, C.J.A.; et al. Preliminary Assessment of the Efficacy of a T-Cell–Based Influenza Vaccine, MVA-NP+M1, in Humans. Clin. Infect. Dis. 2012, 55, 19–25. [Google Scholar] [CrossRef]
- Hooper, J.W.; Larsen, T.; Custer, D.M.; Schmaljohn, C.S. A Lethal Disease Model for Hantavirus Pulmonary Syndrome. Virology 2001, 289, 6–14. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
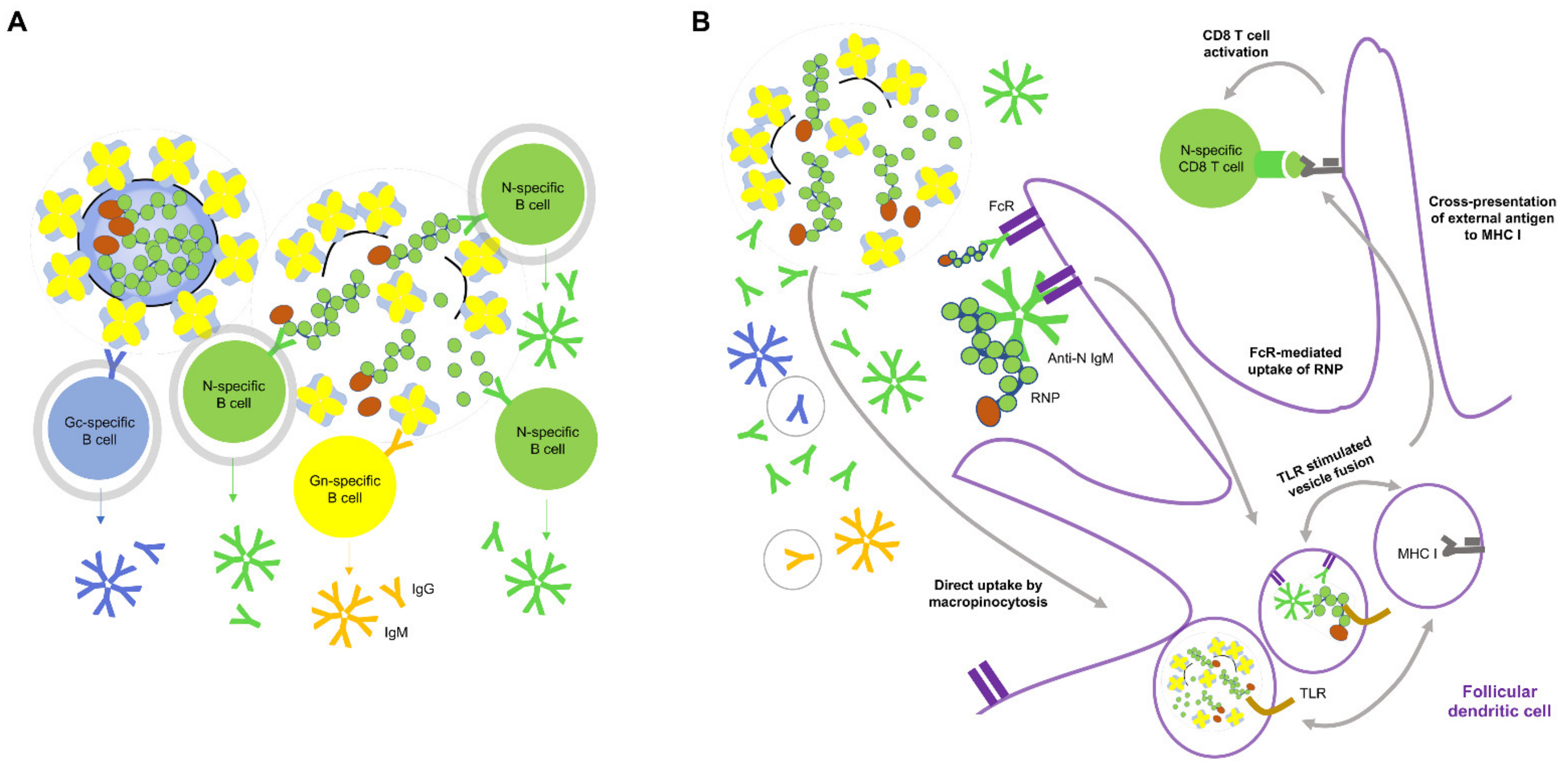{kind=link}
| Serum Antibody Positivity (%) at a Given Time Point after Vaccination (Month) b | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Study | No. of Study Participants a | Vaccine Dosage | Test Method | Day 0 | 1 | 2 | 3 | 12 | 13 | 14 | 15 | 17 | 25 | 37 | 49 |
| ref. 33 | 64 −> 14 | 3 | Neutralization | 0 | 13 | 75 | 14.2 | 50 | |||||||
| IFA | 0 | 79 | 97 | 37 | 94 | ||||||||||
| ref. 34 | 30 −> 30 | 2 | Neutralization | 0 | 3.3 | 16.7 | |||||||||
| c Neutralization + | 0 | 6.7 | 33.3 | ||||||||||||
| ELISA | 0 | 46.7 | 76.7 | ||||||||||||
| ref. 35 | 142 −> 64 | 3 | Neutralization | 0 | n.d. | 23.24 | 1.41 | 45.07 | 40.63 | 15.63 | 12.5 | ||||
| IFA | 0 | n.d. | 90 | 10.56 | 87.32 | 34.68 | 17.74 | 10.48 | |||||||
| ref. 36 | 289 −> 277 | 4 | Neutralization | 0 | n.d. | 40.97 | 72.32 | 7.61 | 55.71 | 41.91 | 27.44 | ||||
| IFA | 0 | n.d. | 83.03 | 92.81 | 21.22 | 95.68 | 67.16 | 51.13 | |||||||
| Vaccine | Immune Responses after Vaccination | Recall Responses after Virus Infection |
|---|---|---|
| Hantavax (iIWV a) |
|
|
| Hantavax (semi-iIWV) |
|
|
| Recombinant N vaccine (rN) |
|
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bae, J.-Y.; Kim, J.I.; Park, M.S.; Lee, G.E.; Park, H.; Song, K.-J.; Park, M.-S. The Immune Correlates of Orthohantavirus Vaccine. Vaccines 2021, 9, 518. https://doi.org/10.3390/vaccines9050518
Bae J-Y, Kim JI, Park MS, Lee GE, Park H, Song K-J, Park M-S. The Immune Correlates of Orthohantavirus Vaccine. Vaccines. 2021; 9(5):518. https://doi.org/10.3390/vaccines9050518
Chicago/Turabian StyleBae, Joon-Yong, Jin Il Kim, Mee Sook Park, Gee Eun Lee, Heedo Park, Ki-Joon Song, and Man-Seong Park. 2021. "The Immune Correlates of Orthohantavirus Vaccine" Vaccines 9, no. 5: 518. https://doi.org/10.3390/vaccines9050518
APA StyleBae, J.-Y., Kim, J. I., Park, M. S., Lee, G. E., Park, H., Song, K.-J., & Park, M.-S. (2021). The Immune Correlates of Orthohantavirus Vaccine. Vaccines, 9(5), 518. https://doi.org/10.3390/vaccines9050518