
Immune Responses to Irradiated Pneumococcal Whole Cell Vaccine

 , , , and
, , , and 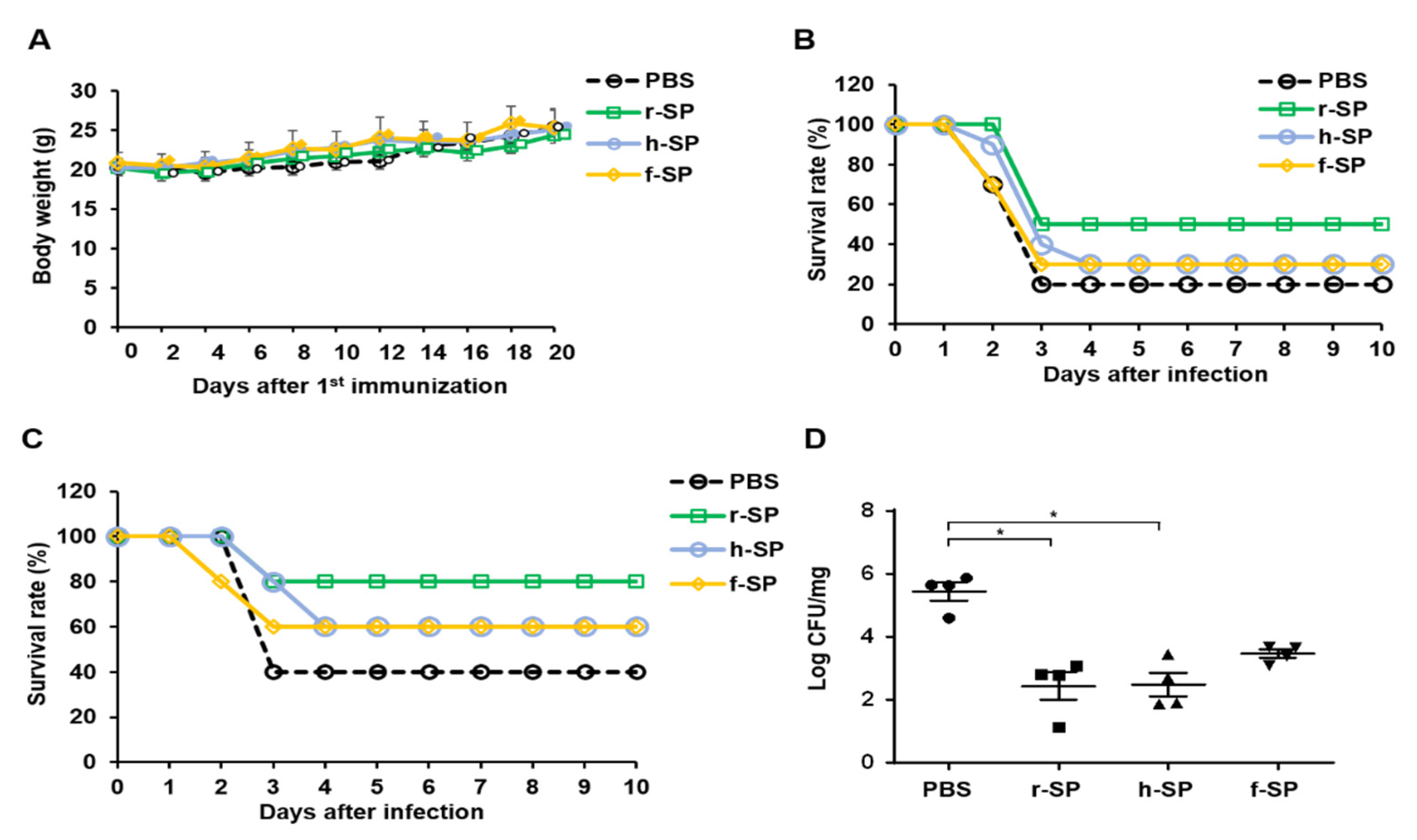{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents and Chemicals
2.2. Generation of Non-Encapsulated S. pneumoniae
2.3. Preparation of Inactivated S. pneumoniae Whole-Cell Vaccines
2.4. Intranasal Immunization
2.5. Measurement of S. pneumoniae-Specific Antibodies
2.6. Analysis of the Immune Cell Population and Intracellular Cytokine Expression
2.7. Pneumococcal Infection
2.8. Statistical Analysis
3. Results
3.1. Immunization with r-SP Induces Higher Protective Immunity Against Pneumococcal Systemic Infection Than Does h-SP or f-SP
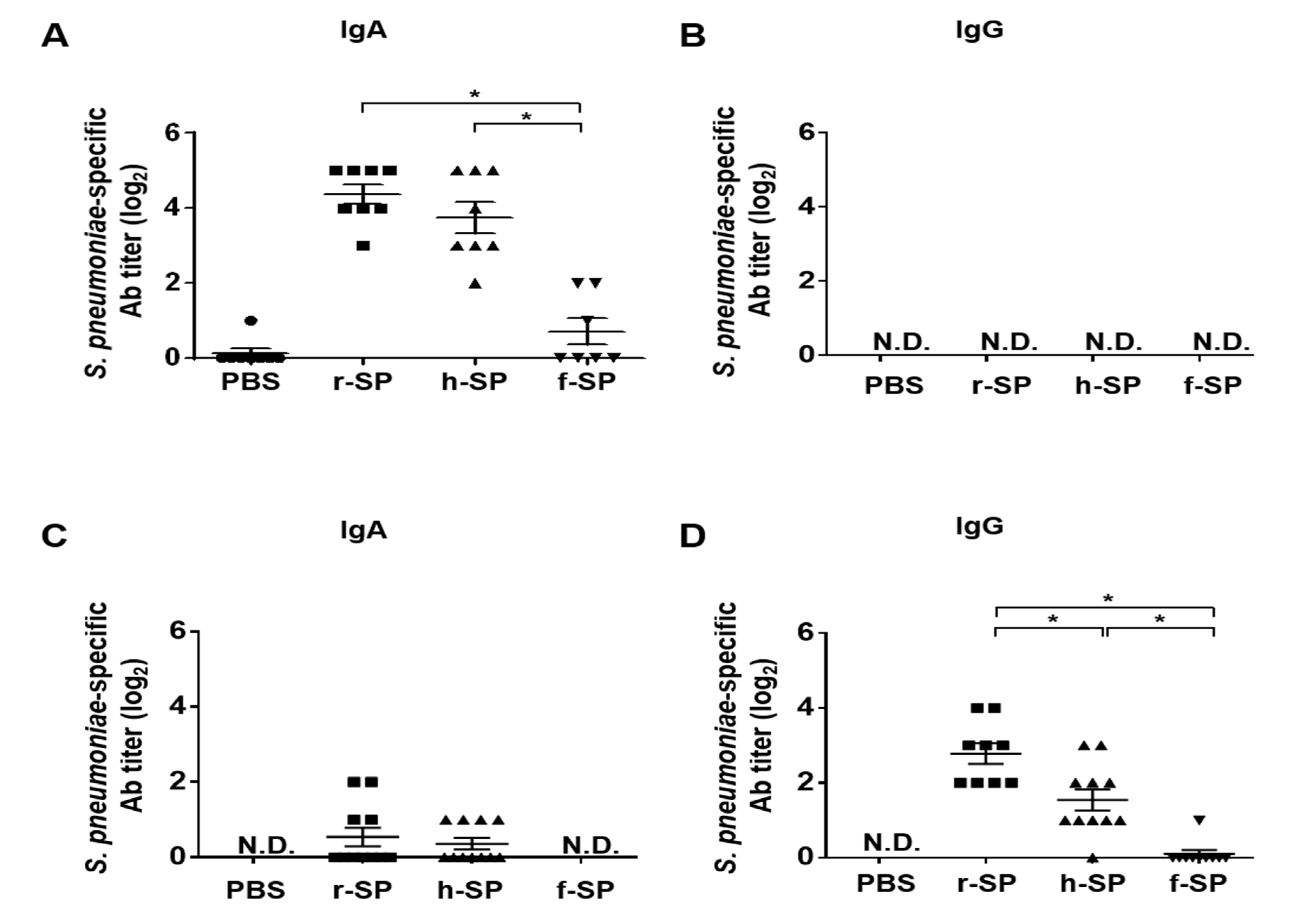
3.2. Immunization with r-SP Enhances S. pneumoniae-Specific Antibodies in the Nasal Cavity and Lung
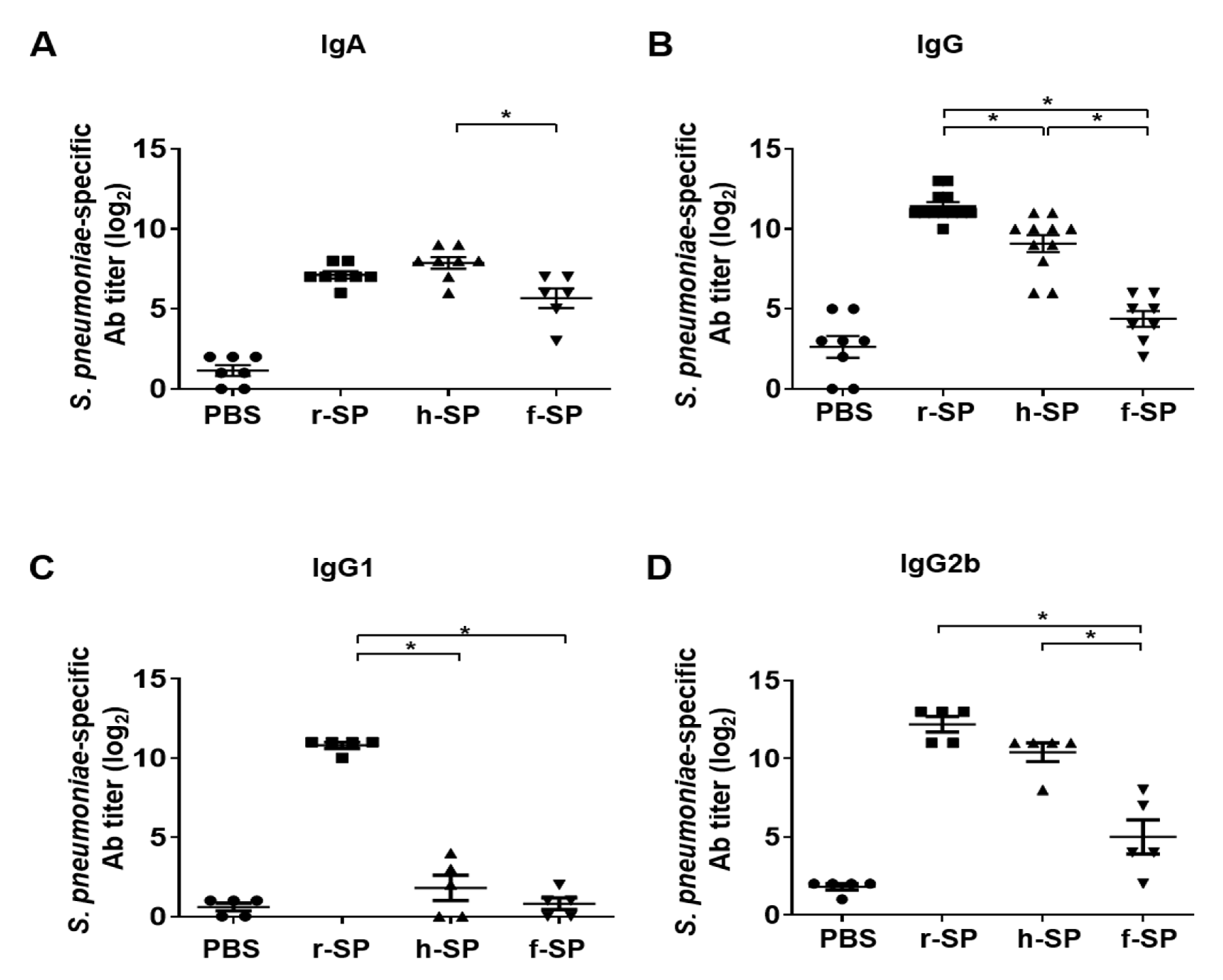
3.3. Immunization with r-SP Increases S. pneumoniae-Specific Serum IgG and IgA
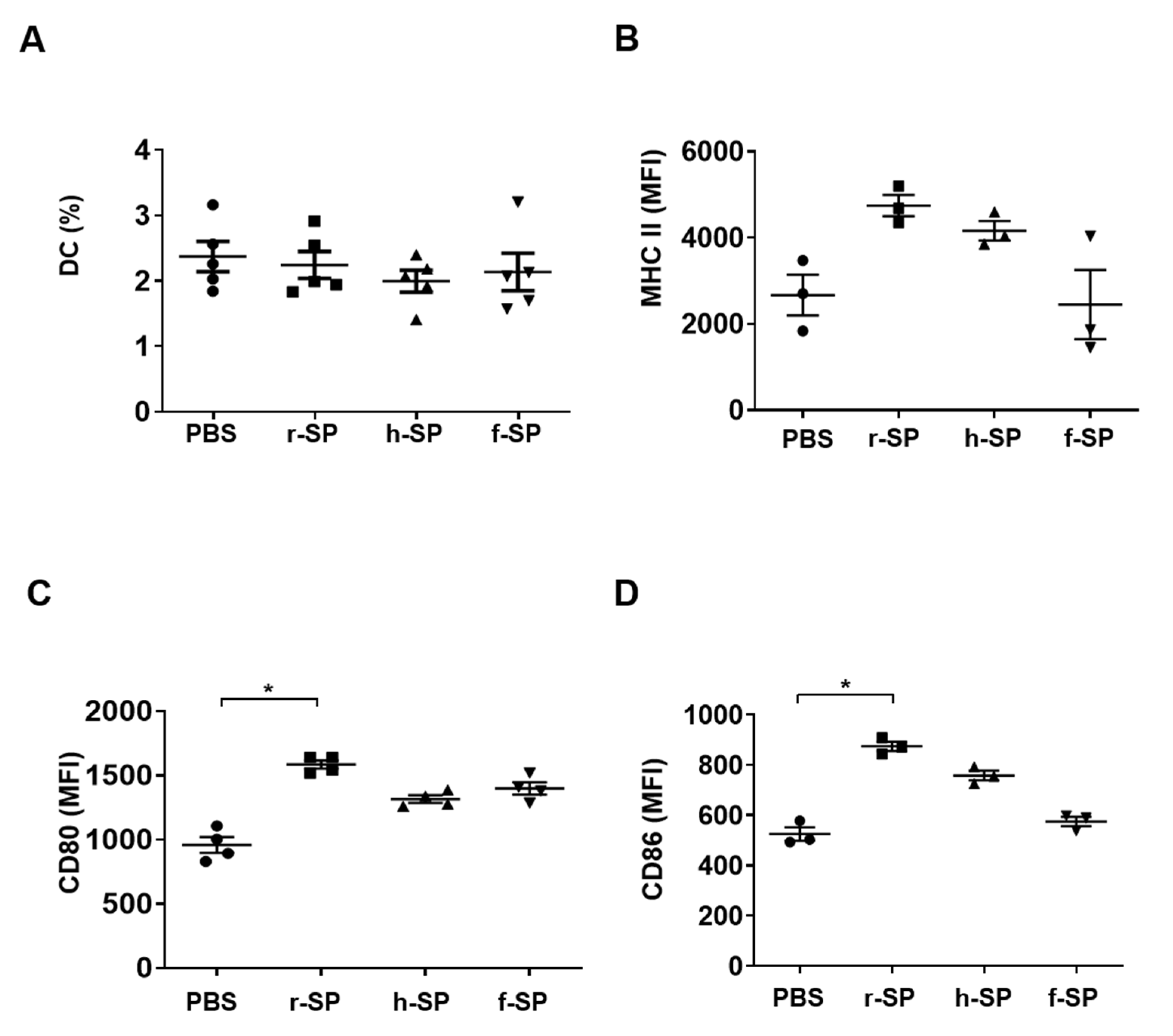
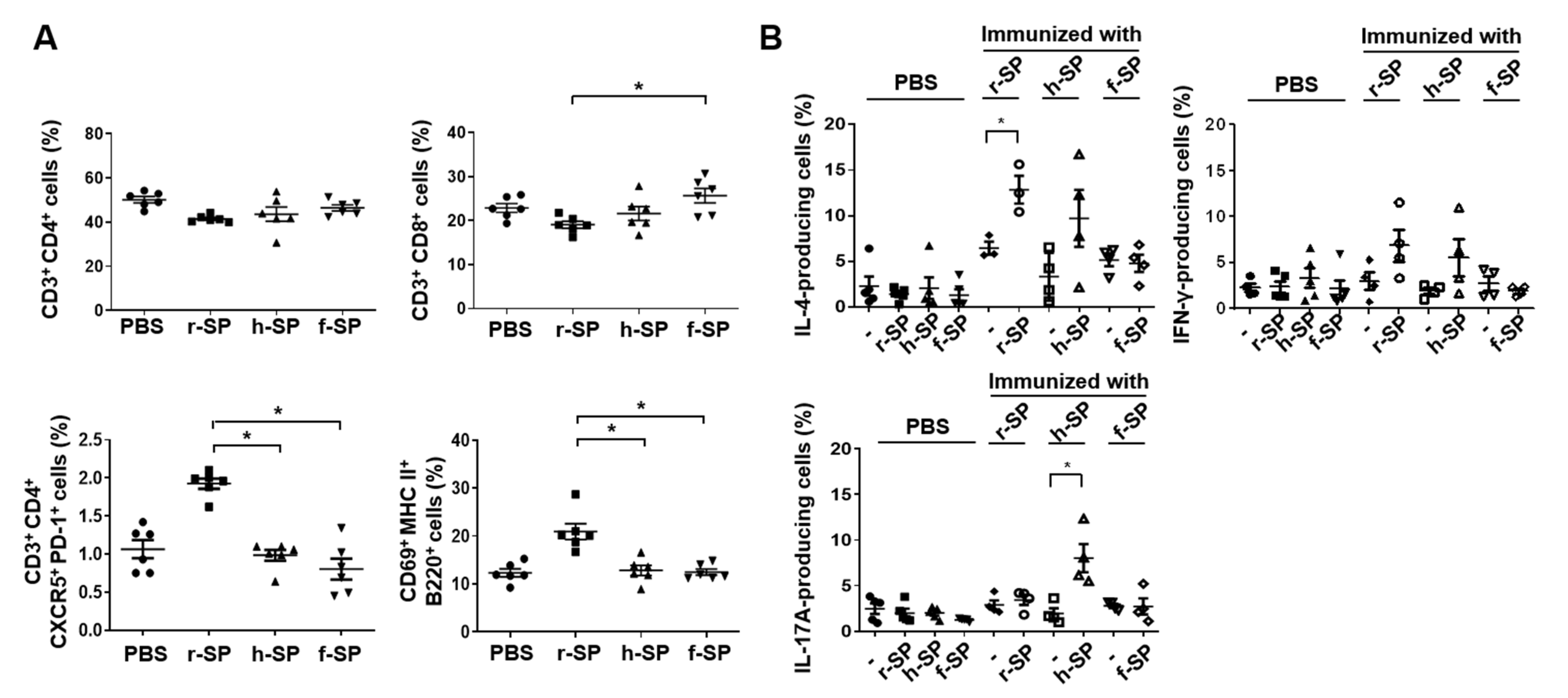
3.4. Enhancement of DC Maturation by r-SP Immunization
3.5. Increases in the Populations of Follicular Helper T (Tfh) Cells and B Cells in the Cervical Lymph Node After r-SP Immunization
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bogaert, D.; De Groot, R.; Hermans, P.W. Streptococcus pneumoniae colonisation: The key to pneumococcal disease. Lancet Infect. Dis. 2004, 4, 144–154. [Google Scholar] [CrossRef]
- Kadioglu, A.; Weiser, J.N.; Paton, J.C.; Andrew, P.W. The role of Streptococcus pneumoniae virulence factors in host respiratory colonization and disease. Nat. Rev. Microbiol. 2008, 6, 288–301. [Google Scholar] [CrossRef] [PubMed]
- Wuerth, B.A.; Bonnewell, J.P.; Wiemken, T.L.; Arnold, F.W. Trends in Pneumonia Mortality Rates and Hospitalizations by Organism, United States, 2002–2011(1). Emerg. Infect. Dis. 2016, 22, 1624–1627. [Google Scholar] [CrossRef]
- Johnson, H.L.; Deloria-Knoll, M.; Levine, O.S.; Stoszek, S.K.; Freimanis Hance, L.; Reithinger, R.; Muenz, L.R.; O’Brien, K.L. Systematic evaluation of serotypes causing invasive pneumococcal disease among children under five: The pneumococcal global serotype project. PLoS Med 2010, 7. [Google Scholar] [CrossRef]
- Jedrzejas, M.J. Pneumococcal virulence factors: Structure and function. Microbiol. Mol. Biol. Rev. 2001, 65, 187–207. [Google Scholar] [CrossRef]
- Brooks, L.R.K.; Mias, G.I. Streptococcus pneumoniae’s Virulence and Host Immunity: Aging, Diagnostics, and Prevention. Front. Immunol. 2018, 9, 1366. [Google Scholar] [CrossRef] [PubMed]
- Kadioglu, A.; Taylor, S.; Iannelli, F.; Pozzi, G.; Mitchell, T.J.; Andrew, P.W. Upper and lower respiratory tract infection by Streptococcus pneumoniae is affected by pneumolysin deficiency and differences in capsule type. Infect. Immun. 2002, 70, 2886–2890. [Google Scholar] [CrossRef] [PubMed]
- Hyams, C.; Camberlein, E.; Cohen, J.M.; Bax, K.; Brown, J.S. The Streptococcus pneumoniae capsule inhibits complement activity and neutrophil phagocytosis by multiple mechanisms. Infect. Immun. 2010, 78, 704–715. [Google Scholar] [CrossRef] [PubMed]
- Ndlangisa, K.; du Plessis, M.; Allam, M.; Wolter, N.; de Gouveia, L.; Klugman, K.P.; Cohen, C.; Gladstone, R.A.; von Gottberg, A. Invasive Disease Caused Simultaneously by Dual Serotypes of Streptococcus pneumoniae. J. Clin. Microbiol. 2018, 56. [Google Scholar] [CrossRef]
- Finkelstein, J.A.; Huang, S.S.; Daniel, J.; Rifas-Shiman, S.L.; Kleinman, K.; Goldmann, D.; Pelton, S.I.; DeMaria, A.; Platt, R. Antibiotic-resistant Streptococcus pneumoniae in the heptavalent pneumococcal conjugate vaccine era: Predictors of carriage in a multicommunity sample. Pediatrics 2003, 112, 862–869. [Google Scholar] [CrossRef]
- Linares, J.; Ardanuy, C.; Pallares, R.; Fenoll, A. Changes in antimicrobial resistance, serotypes and genotypes in Streptococcus pneumoniae over a 30-year period. Clin. Microbiol. Infect. 2010, 16, 402–410. [Google Scholar] [CrossRef] [PubMed]
- Pollard, A.J.; Perrett, K.P.; Beverley, P.C. Maintaining protection against invasive bacteria with protein-polysaccharide conjugate vaccines. Nat. Rev. Immunol. 2009, 9, 213–220. [Google Scholar] [CrossRef] [PubMed]
- de Roux, A.; Schmole-Thoma, B.; Siber, G.R.; Hackell, J.G.; Kuhnke, A.; Ahlers, N.; Baker, S.A.; Razmpour, A.; Emini, E.A.; Fernsten, P.D.; et al. Comparison of pneumococcal conjugate polysaccharide and free polysaccharide vaccines in elderly adults: Conjugate vaccine elicits improved antibacterial immune responses and immunological memory. Clin. Infect. Dis. 2008, 46, 1015–1023. [Google Scholar] [CrossRef]
- Musher, D.M.; Manof, S.B.; Liss, C.; McFetridge, R.D.; Marchese, R.D.; Bushnell, B.; Alvarez, F.; Painter, C.; Blum, M.D.; Silber, J.L. Safety and antibody response, including antibody persistence for 5 years, after primary vaccination or revaccination with pneumococcal polysaccharide vaccine in middle-aged and older adults. J. Infect. Dis. 2010, 201, 516–524. [Google Scholar] [CrossRef]
- Moffitt, K.L.; Malley, R. Next generation pneumococcal vaccines. Curr. Opin. Immunol. 2011, 23, 407–413. [Google Scholar] [CrossRef] [PubMed]
- Waight, P.A.; Andrews, N.J.; Ladhani, S.N.; Sheppard, C.L.; Slack, M.P.; Miller, E. Effect of the 13-valent pneumococcal conjugate vaccine on invasive pneumococcal disease in England and Wales 4 years after its introduction: An observational cohort study. Lancet Infect. Dis. 2015, 15, 535–543. [Google Scholar] [CrossRef]
- Plotkin, S.A. Vaccines: Past, present and future. Nat. Med. 2005, 11, S5–S11. [Google Scholar] [CrossRef] [PubMed]
- Pace, J.L.; Rossi, H.A.; Esposito, V.M.; Frey, S.M.; Tucker, K.D.; Walker, R.I. Inactivated whole-cell bacterial vaccines: Current status and novel strategies. Vaccine 1998, 16, 1563–1574. [Google Scholar] [CrossRef]
- Datta, S.K.; Okamoto, S.; Hayashi, T.; Shin, S.S.; Mihajlov, I.; Fermin, A.; Guiney, D.G.; Fierer, J.; Raz, E. Vaccination with irradiated Listeria induces protective T cell immunity. Immunity 2006, 25, 143–152. [Google Scholar] [CrossRef]
- Ingram, L.O. Ethanol tolerance in bacteria. Crit. Rev. Biotechnol. 1990, 9, 305–319. [Google Scholar] [CrossRef]
- Magnani, D.M.; Harms, J.S.; Durward, M.A.; Splitter, G.A. Nondividing but metabolically active gamma-irradiated Brucella melitensis is protective against virulent B. melitensis challenge in mice. Infect. Immun. 2009, 77, 5181–5189. [Google Scholar] [CrossRef]
- Seo, H.S. Application of radiation technology in vaccines development. Clin. Exp. Vaccine Res. 2015, 4, 145–158. [Google Scholar] [CrossRef]
- Babb, R.; Chen, A.; Hirst, T.R.; Kara, E.E.; McColl, S.R.; Ogunniyi, A.D.; Paton, J.C.; Alsharifi, M. Intranasal vaccination with gamma-irradiated Streptococcus pneumoniae whole-cell vaccine provides serotype-independent protection mediated by B-cells and innate IL-17 responses. Clin. Sci. 2016, 130, 697–710. [Google Scholar] [CrossRef]
- Jwa, M.Y.; Jeong, S.; Ko, E.B.; Kim, A.R.; Kim, H.Y.; Kim, S.K.; Seo, H.S.; Yun, C.H.; Han, S.H. Gamma-irradiation of Streptococcus pneumoniae for the use as an immunogenic whole cell vaccine. J. Microbiol. 2018, 56, 579–585. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.Y.; Kim, S.K.; Seo, H.S.; Jeong, S.; Ahn, K.B.; Yun, C.H.; Han, S.H. Th17 activation by dendritic cells stimulated with gamma-irradiated Streptococcus pneumoniae. Mol. Immunol. 2018, 101, 344–352. [Google Scholar] [CrossRef]
- Seo, H.S.; Xiong, Y.Q.; Mitchell, J.; Seepersaud, R.; Bayer, A.S.; Sullam, P.M. Bacteriophage lysin mediates the binding of Streptococcus mitis to human platelets through interaction with fibrinogen. PLoS Pathog. 2010, 6, e1001047. [Google Scholar] [CrossRef]
- Alloing, G.; Martin, B.; Granadel, C.; Claverys, J.P. Development of competence in Streptococcus pneumoniae: Pheromone autoinduction and control of quorum sensing by the oligopeptide permease. Mol. Microbiol. 1998, 29, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Inzana, T.J.; Champion, A. Use of an inhibition enzyme-linked immunosorbent assay for quantification of capsular polysaccharide or proteins in vaccines. Clin. Vaccine Immunol. 2007, 14, 323–327. [Google Scholar] [CrossRef] [PubMed]
- Zapotoczna, M.; McCarthy, H.; Rudkin, J.K.; O’Gara, J.P.; O’Neill, E. An essential role for coagulase in Staphylococcus aureus biofilm development reveals new therapeutic possibilities for device-related infections. J. Infect. Dis. 2015, 212, 1883–1893. [Google Scholar] [CrossRef]
- Hvalbye, B.K.; Aaberge, I.S.; Lovik, M.; Haneberg, B. Intranasal immunization with heat-inactivated Streptococcus pneumoniae protects mice against systemic pneumococcal infection. Infect. Immun. 1999, 67, 4320–4325. [Google Scholar] [CrossRef]
- Xu, X.; Meng, J.; Wang, Y.; Zheng, J.; Wu, K.; Zhang, X.; Yin, Y.; Zhang, Q. Serotype-independent protection against pneumococcal infections elicited by intranasal immunization with ethanol-killed pneumococcal strain, SPY1. J. Microbiol. 2014, 52, 315–323. [Google Scholar] [CrossRef]
- Miller, M.A.; Stabenow, J.M.; Parvathareddy, J.; Wodowski, A.J.; Fabrizio, T.P.; Bina, X.R.; Zalduondo, L.; Bina, J.E. Visualization of murine intranasal dosing efficiency using luminescent Francisella tularensis: Effect of instillation volume and form of anesthesia. PLoS ONE 2012, 7, e31359. [Google Scholar] [CrossRef] [PubMed]
- Silva-Carvalho, R.; Silva, J.P.; Ferreirinha, P.; Leitao, A.F.; Andrade, F.K.; Gil da Costa, R.M.; Cristelo, C.; Rosa, M.F.; Vilanova, M.; Gama, F.M. Inhalation of bacterial cellulose nanofibrils triggers an inflammatory response and changes lung tissue morphology of mice. Toxicol. Res. 2019, 35, 45–63. [Google Scholar] [CrossRef]
- Steinman, R.M.; Hemmi, H. Dendritic cells: Translating innate to adaptive immunity. Curr. Top. Microbiol. Immunol. 2006, 311, 17–58. [Google Scholar] [CrossRef]
- Yamamoto, T. Current status of cholera and rise of novel mucosal vaccine. Jpn. J. Infect. Dis. 2000, 53, 181–188. [Google Scholar] [PubMed]
- Gaidamakova, E.K.; Myles, I.A.; McDaniel, D.P.; Fowler, C.J.; Valdez, P.A.; Naik, S.; Gayen, M.; Gupta, P.; Sharma, A.; Glass, P.J.; et al. Preserving immunogenicity of lethally irradiated viral and bacterial vaccine epitopes using a radio- protective Mn2+-Peptide complex from Deinococcus. Cell Host Microbe 2012, 12, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.C.; Chiu, H.C.; Chen, L.K.; Chang, G.J.; Chiou, S.S. Formalin inactivation of Japanese encephalitis virus vaccine alters the antigenicity and immunogenicity of a neutralization epitope in envelope protein domain III. PLoS Negl. Trop. Dis. 2015, 9, e0004167. [Google Scholar] [CrossRef] [PubMed]
- Crotty, S. T follicular helper cell differentiation, function, and roles in disease. Immunity 2014, 41, 529–542. [Google Scholar] [CrossRef]
- Crotty, S. Follicular helper CD4 T cells (TFH). Annu. Rev. Immunol. 2011, 29, 621–663. [Google Scholar] [CrossRef]
- Ma, C.S.; Deenick, E.K.; Batten, M.; Tangye, S.G. The origins, function, and regulation of T follicular helper cells. J. Exp. Med. 2012, 209, 1241–1253. [Google Scholar] [CrossRef]
- Gray, C.; Ahmed, M.S.; Mubarak, A.; Kasbekar, A.V.; Derbyshire, S.; McCormick, M.S.; Mughal, M.K.; McNamara, P.S.; Mitchell, T.; Zhang, Q. Activation of memory Th17 cells by domain 4 pneumolysin in human nasopharynx-associated lymphoid tissue and its association with pneumococcal carriage. Mucosal Immunol. 2014, 7, 705–717. [Google Scholar] [CrossRef] [PubMed]
- Jwa, M.Y.; Ko, E.B.; Kim, H.Y.; Kim, S.K.; Jeong, S.; Seo, H.S.; Yun, C.H.; Han, S.H. Gamma-irradiation-killed Streptococcus pneumoniae potently induces the expression of IL-6 and IL-8 in human bronchial epithelial cells. Microb. Pathog. 2018, 124, 38–46. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ko, E.; Jeong, S.; Jwa, M.Y.; Kim, A.R.; Ha, Y.-E.; Kim, S.K.; Jeong, S.; Ahn, K.B.; Seo, H.S.; Yun, C.-H.; et al. Immune Responses to Irradiated Pneumococcal Whole Cell Vaccine. Vaccines 2021, 9, 405. https://doi.org/10.3390/vaccines9040405
Ko E, Jeong S, Jwa MY, Kim AR, Ha Y-E, Kim SK, Jeong S, Ahn KB, Seo HS, Yun C-H, et al. Immune Responses to Irradiated Pneumococcal Whole Cell Vaccine. Vaccines. 2021; 9(4):405. https://doi.org/10.3390/vaccines9040405
Chicago/Turabian StyleKo, Eunbyeol, Soyoung Jeong, Min Yong Jwa, A Reum Kim, Ye-Eun Ha, Sun Kyung Kim, Sungho Jeong, Ki Bum Ahn, Ho Seong Seo, Cheol-Heui Yun, and et al. 2021. "Immune Responses to Irradiated Pneumococcal Whole Cell Vaccine" Vaccines 9, no. 4: 405. https://doi.org/10.3390/vaccines9040405
APA StyleKo, E., Jeong, S., Jwa, M. Y., Kim, A. R., Ha, Y.-E., Kim, S. K., Jeong, S., Ahn, K. B., Seo, H. S., Yun, C.-H., & Han, S. H. (2021). Immune Responses to Irradiated Pneumococcal Whole Cell Vaccine. Vaccines, 9(4), 405. https://doi.org/10.3390/vaccines9040405