
Selection of Vaccine Candidate for Foot-and-Mouth Disease Virus Serotype O Using a Blocking Enzyme-Linked Immunosorbent Assay


Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Serum Samples
2.3. Cells and Viruses
2.4. Vaccine Preparation
2.5. Animal Vaccination
2.6. Virus Neutralizing Test
2.7. NA-ELISA
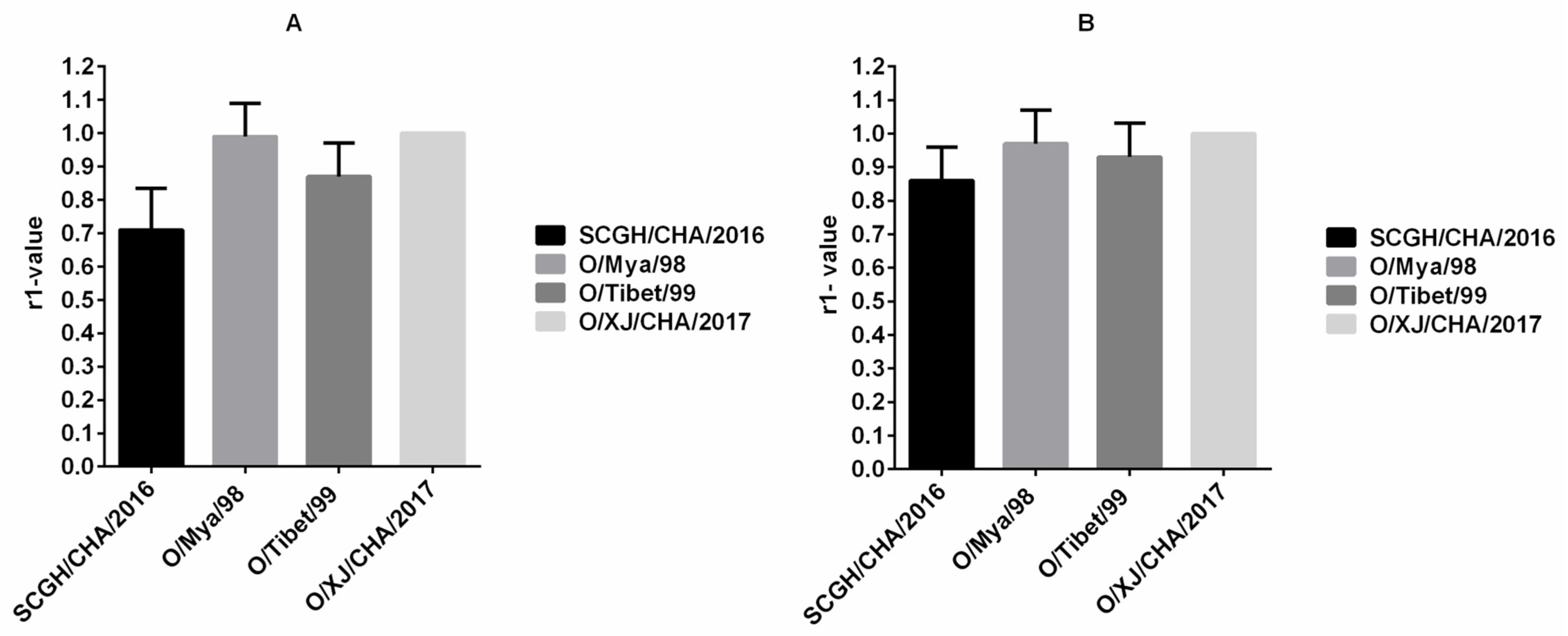
2.8. The r1-Value Determination
2.9. Statistical Analysis
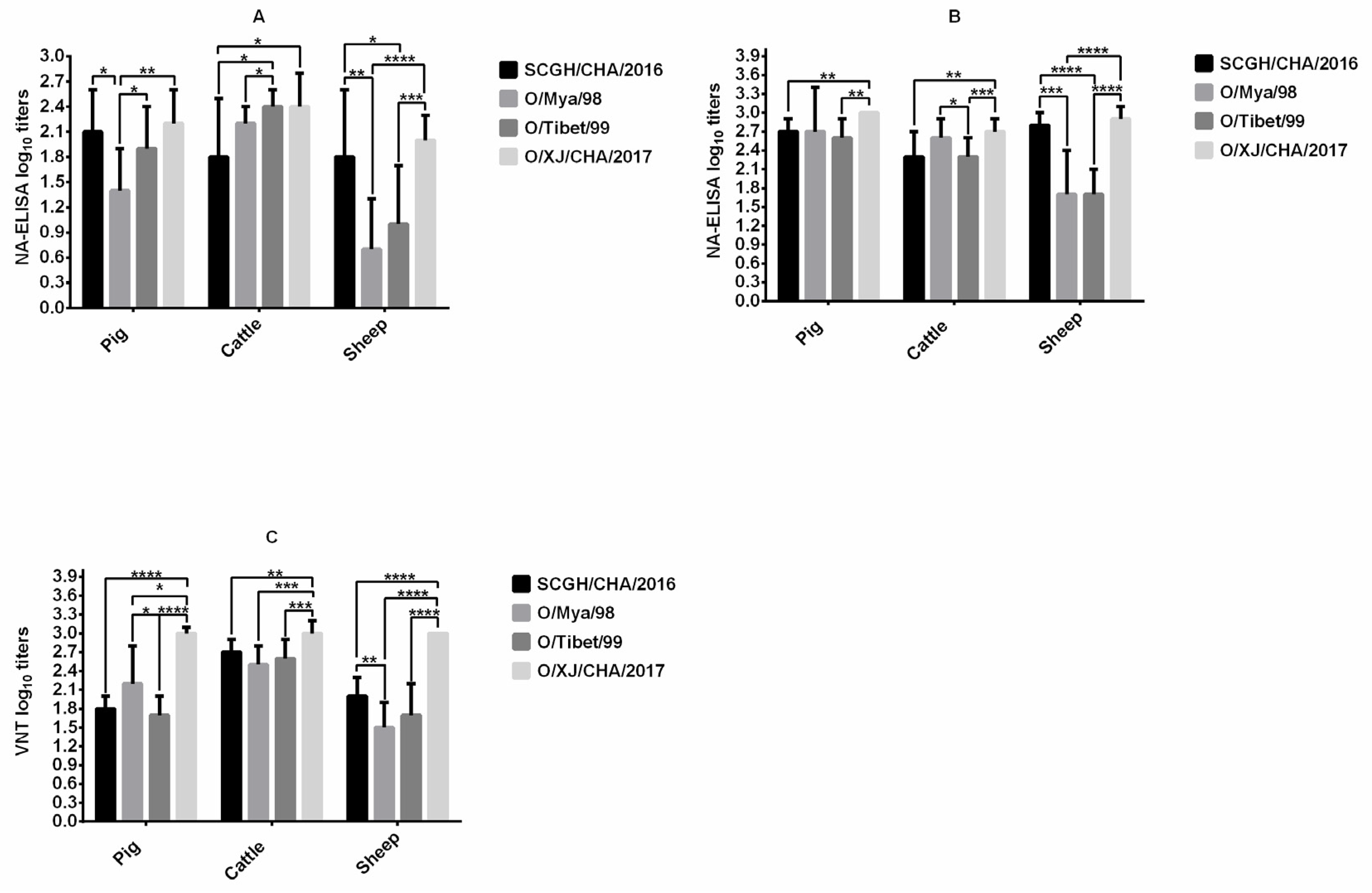
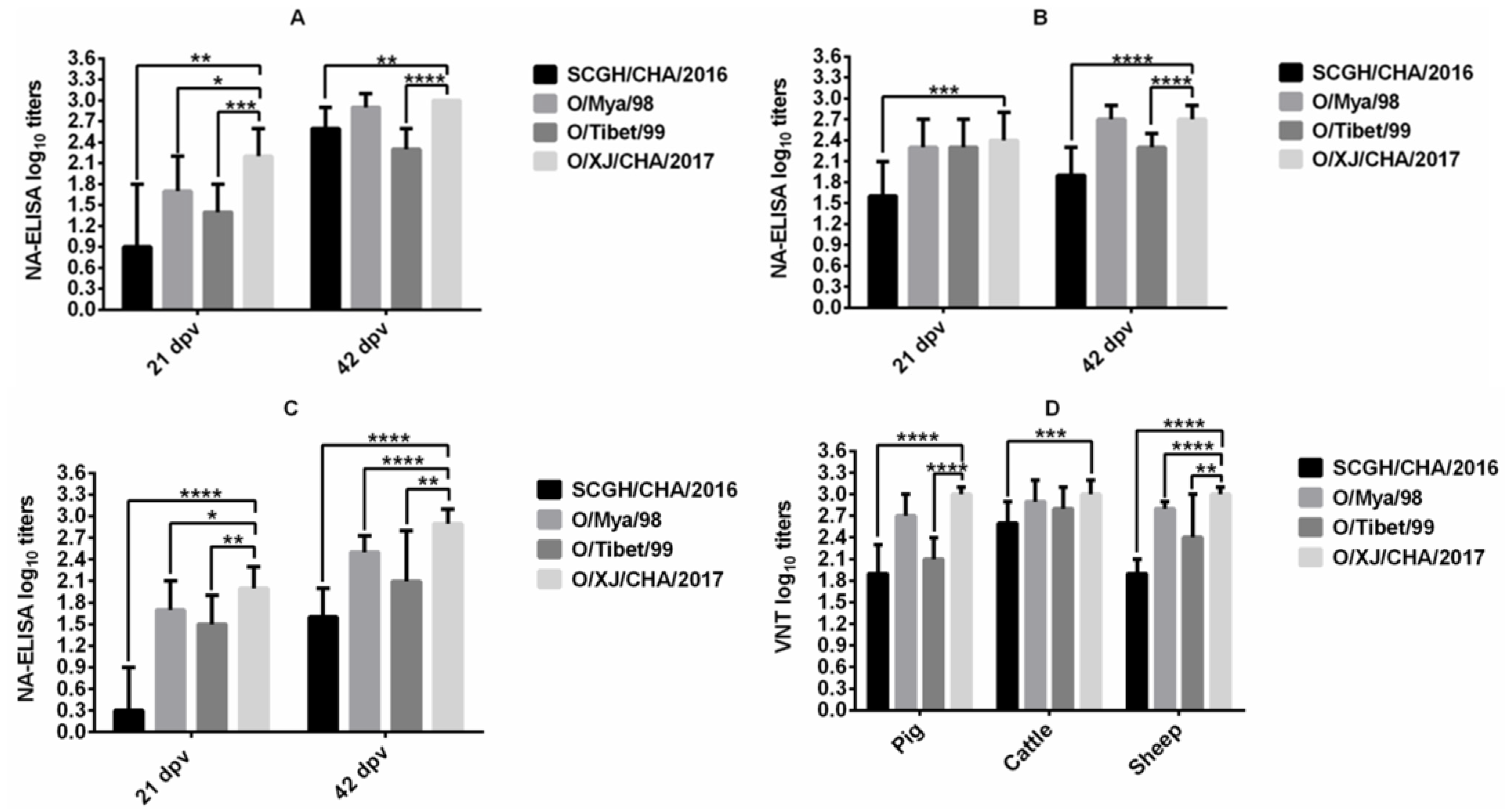
3. Results
3.1. Comparison of Immunogenicity in Cattle, Sheep, and Pigs
3.2. Cross-Neutralization Analysis
3.3. Vaccine Matching Test Using Vaccinated Cattle Sera
3.4. Correlation between Antibody Titers against Each Topotype Virus in Vaccinated Animals Detected by NA-ELISA and VNT
3.5. Relationship between NA-ELISA Titers of Vaccinated Pigs and Clinical Protection against FMDV Serotype O
4. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Grubman, M.J.; Baxt, B. Foot-and-mouth disease. Clin. Microbiol. Rev. 2004, 17, 465–493. [Google Scholar] [CrossRef]
- Knight-Jones, T.J.; Rushton, J. The economic impacts of foot and mouth disease—What are they, how big are they and where do they occur? Prev. Vet. Med. 2013, 112, 161–173. [Google Scholar] [CrossRef]
- Brooksby, J.B. The virus of foot-and-mouth disease. Adv. Virus Res. 1958, 5, 1–37. [Google Scholar]
- Domingo, E.; Escarmis, C.; Baranowski, E.; Ruiz-Jarabo, C.M.; Carrillo, E.; Nunez, J.I.; Sobrino, F. Evolution of foot-and-mouth disease virus. Virus Res. 2003, 91, 47–63. [Google Scholar] [CrossRef]
- Knowles, N.J.; Wadsworth, J.; Bachanek-Bankowska, K.; King, D.P. VP1 sequencing protocol for foot and mouth disease virus molecular epidemiology. Rev. Sci. Tech. 2016, 35, 741–755. [Google Scholar] [CrossRef] [PubMed]
- Paton, D.J.; Valarcher, J.F.; Bergmann, I.; Matlho, O.G.; Zakharov, V.M.; Palma, E.L.; Thomson, G.R. Selection of foot and mouth disease vaccine strains—A review. Rev. Sci. Tech. Off. Int. Epiz. 2005, 24, 981–993. [Google Scholar] [CrossRef]
- Zhu, Z.; Yang, F.; He, J.; Li, J.; Cao, W.; Li, J.; Xia, Y.; Guo, J.; Jin, Y.; Zhang, K.; et al. First detection of foot-and-mouth disease virus O/ME-SA/Ind2001 in China. Transbound. Emerg. Dis. 2018, 65, 2027–2031. [Google Scholar] [CrossRef] [PubMed]
- Li, K.B.; Liang, J.P. Assess the effect of the foot and mouth disease type O inactivated vaccine (O/MYA98/BY/2010 strain) for pigs. Adv. Mater. Res. 2012, 468–471, 365–369. [Google Scholar] [CrossRef]
- Zheng, H.X.; He, J.J.; Guo, J.H.; Jin, Y.; Yang, F.; Lv, L.; Liu, X.T. Genetic characterization of a new pandemic Southeast Asia topotype strain of serotype O foot-and-mouth disease virus isolated in China during 2010. Virus Genes 2012, 44, 80–88. [Google Scholar] [CrossRef]
- World Organisation for Animal Health. Foot and Mouth Disease. In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals, 7th ed.; Office International des Epizooties: Paris, France, 2012; Volume 2, pp. 145–173. [Google Scholar]
- Rweyemamu, M.M.; Booth, J.C.; Head, M.; Pay, T.W. Microneutralization tests for serological typing and subtyping of foot-and-mouth disease virus strains. J. Hyg. 1978, 81, 107–123. [Google Scholar] [CrossRef] [PubMed]
- Haas, B. Application of the FMD Liquid-Phase Blocking Sandwich ELISA. Problems Encountered in Import/Export Serology and Possible Solutions; Report of the Session of the Research Group of the Standing Technical Committee of the European Commission for the Control of Foot-and-Mouth Disease; European Commission for the Control of Foot-and-Mouth Disease: Vienna, Austria, 1994; pp. 124–127. [Google Scholar]
- Mackay, D.K.; Bulut, A.N.; Rendle, T.; Davidson, F.; Ferris, N.P. A solid-phase competition ELISA for measuring antibody to foot-and-mouth disease virus. J. Virol. Methods 2001, 97, 33–48. [Google Scholar] [CrossRef]
- Cao, Y.M.; Li, K.; Wang, S.; Fu, Y.F.; Sun, P.; Li, P.H.; Bai, X.W.; Zhang, J.; Ma, X.Q.; Xing, X.C.; et al. Implication of Broadly Neutralizing Bovine Monoclonal Antibodies in the Development of an Enzyme-Linked Immunosorbent Assay for Detecting Neutralizing Antibodies against Foot-and-Mouth Disease Virus Serotype O. J. Clin. Microbiol. 2019, 57, e01030-19. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.M.; Lu, Z.J.; Sun, J.C.; Bai, X.W.; Sun, P.; Bao, H.F.; Chen, Y.L.; Guo, J.H.; Liu, X.T.; Liu, Z.X. Synthesis of empty capsid-like particles of Asia I foot-and-mouth disease virus in insect cells and their immunogenicity in guinea pigs. Vet. Microbiol. 2009, 137, 10–17. [Google Scholar] [CrossRef]
- Van Maanen, C.; Terpstra, C. Comparison of a liquid-phase blocking sandwich ELISA and a serum neutralization test to evaluate immunity in potency tests of foot-and-mouth disease vaccines. J. Immunol. Methods 1989, 124, 111–119. [Google Scholar] [CrossRef]
- Hingley, P.J.; Pay, T.W. Sources of variability in foot and mouth disease vaccine potency estimates based on serum neutralizing antibody assay. J. Biol. Stand. 1987, 15, 127–142. [Google Scholar] [CrossRef]
- OIE. Chapter 3.1.8. Foot and mouth disease (Infection with foot and mouth disease virus). In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals, 8th ed.; OIE: Paris, France, 2018; pp. 433–467. [Google Scholar]
- Mattion, N.; Goris, N.; Willems, T.; Robiolo, B.; Maradei, E.; Beascoechea, C.P.; Perez, A.; Smitsaart, E.; Fondevila, N.; Palma, E.; et al. Some guidelines for determining foot-and-mouth disease vaccine strain matching by serology. Vaccine 2009, 27, 741–747. [Google Scholar] [CrossRef]
- Brito, B.P.; Perez, A.M.; Capozzo, A.V. Accuracy of traditional and novel serology tests for predicting cross-protection in foot-and-mouth disease vaccinated cattle. Vaccine 2014, 32, 433–436. [Google Scholar] [CrossRef]
- Li, D.; Lu, Z.J.; Xie, B.X.; Sun, P.; Chen, Y.L.; Fu, Y.F.; Liu, Z.X. Alternative way to test the efficacy of swine FMD vaccines: Measurement of pigs median infected dose (PID50) and regulation of live virus challenge dose. Virol. J. 2010, 7, 215. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Liu, Z.X.; Sun, P.; Li, Y.L.; Lu, Z.J.; Tian, M.N.; Chen, Y.L.; Xie, B.X.; Bao, H.F.; Fu, Y.F.; et al. The efficacy of FMD vaccine reduced non-structural proteins with a mAb against 3B protein. Vet. Res. Commun. 2010, 34, 445–457. [Google Scholar] [CrossRef]
- Robiolo, B.; La Torre, J.; Maradei, E.; Beascoechea, C.P.; Perez, A.; Seki, C.; Smitsaart, E.; Fondevila, N.; Palma, E.; Goris, N.; et al. Confidence in indirect assessment of foot-and-mouth disease vaccine potency and vaccine matching carried out by liquid phase ELISA and virus neutralization tests. Vaccine 2010, 28, 6235–6241. [Google Scholar] [CrossRef]
- McCullough, K.C.; Crowther, J.R.; Butcher, R.N.; Carpenter, W.C.; Brocchi, E.; Capucci, L.; De Simone, F. Immune protection against foot-and-mouth disease virus studied using virus-neutralizing and non-neutralizing concentrations of monoclonal antibodies. Immunology 1986, 58, 421–428. [Google Scholar] [PubMed]
- McCullough, K.C.; Bruckner, L.; Schaffner, R.; Fraefel, W.; Muller, H.K.; Kihm, U. Relationship between the anti-FMD virus antibody reaction as measured by different assays, and protection in vivo against challenge infection. Vet. Microbiol. 1992, 30, 99–112. [Google Scholar] [CrossRef]
- McCullough, K.C.; De Simone, F.; Brocchi, E.; Capucci, L.; Crowther, J.R.; Kihm, U. Protective immune response against foot-and-mouth disease. J. Virol. 1992, 66, 1835–1840. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Animal Species | Group | Vaccine Strain | No. of Animals | Vaccination Dose |
|---|---|---|---|---|
| Pigs | A | SCGH/CHA/2016 | 10 | 2 mL |
| B | O/Mya/98 | 10 | 2 mL | |
| C | O/Tibet/99 | 10 | 2 mL | |
| D | O/XJ/CHA/2017 | 10 | 2 mL | |
| Cattle | E | SCGH/CHA/2016 | 12 | 2 mL |
| F | O/Mya/98 | 12 | 2 mL | |
| G | O/Tibet/99 | 13 | 2 mL | |
| H | O/XJ/CHA/2017 | 13 | 2 mL | |
| Sheep | I | SCGH/CHA/2016 | 12 | 2 mL |
| J | O/Mya/98 | 12 | 2 mL | |
| K | O/Tibet/99 | 13 | 2 mL | |
| L | O/XJ/CHA/2017 | 13 | 2 mL |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, Y.; Li, K.; Xing, X.; Bao, H.; Huang, N.; Zhu, G.; Bai, X.; Sun, P.; Fu, Y.; Li, P.; et al. Selection of Vaccine Candidate for Foot-and-Mouth Disease Virus Serotype O Using a Blocking Enzyme-Linked Immunosorbent Assay. Vaccines 2021, 9, 387. https://doi.org/10.3390/vaccines9040387
Cao Y, Li K, Xing X, Bao H, Huang N, Zhu G, Bai X, Sun P, Fu Y, Li P, et al. Selection of Vaccine Candidate for Foot-and-Mouth Disease Virus Serotype O Using a Blocking Enzyme-Linked Immunosorbent Assay. Vaccines. 2021; 9(4):387. https://doi.org/10.3390/vaccines9040387
Chicago/Turabian StyleCao, Yimei, Kun Li, Xiangchuan Xing, Huifang Bao, Nana Huang, Guoqiang Zhu, Xingwen Bai, Pu Sun, Yuanfang Fu, Pinghua Li, and et al. 2021. "Selection of Vaccine Candidate for Foot-and-Mouth Disease Virus Serotype O Using a Blocking Enzyme-Linked Immunosorbent Assay" Vaccines 9, no. 4: 387. https://doi.org/10.3390/vaccines9040387
APA StyleCao, Y., Li, K., Xing, X., Bao, H., Huang, N., Zhu, G., Bai, X., Sun, P., Fu, Y., Li, P., Zhang, J., Ma, X., Li, D., Liu, Z., & Lu, Z. (2021). Selection of Vaccine Candidate for Foot-and-Mouth Disease Virus Serotype O Using a Blocking Enzyme-Linked Immunosorbent Assay. Vaccines, 9(4), 387. https://doi.org/10.3390/vaccines9040387