Abstract
In the early 1980s, a highly contagious viral hemorrhagic fever in rabbits (Oryctolagus cuniculus) emerged, causing a very high rate of mortality in these animals. Since the initial occurrence of the rabbit hemorrhagic disease virus (RHDV), several hundred million rabbits have died after infection. The emergence of genetically-different virus variants (RHDV GI.1 and GI.2) indicated the very high variability of RHDV. Moreover, with these variants, the host range broadened to hare species (Lepus). The circulation of RHDV genotypes displays different virulences and a limited induction of cross-protective immunity. Interestingly, juvenile rabbits (<9 weeks of age) with an immature immune system display a general resistance to RHDV GI.1, and a limited resistance to RHDV GI.2 strains, whereas less than 3% of adult rabbits survive an infection by either RHDV GI.1. or GI.2. Several not-yet fully understood phenomena characterize the RHD. A very low infection dose followed by an extremely rapid viral replication could be simplified to the induction of a disseminated intravascular coagulopathy (DIC), a severe loss of lymphocytes—especially T-cells—and death within 36 to 72 h post infection. On the other hand, in animals surviving the infection or after vaccination, very high titers of RHDV-neutralizing antibodies were induced. Several studies have been conducted in order to deepen the knowledge about the virus’ genetics, epidemiology, RHDV-induced pathology, and the anti-RHDV immune responses of rabbits in order to understand the phenomenon of the juvenile resistance to this virus. Moreover, several approaches have been used to produce efficient vaccines in order to prevent an infection with RHDV. In this review, we discuss the current knowledge about anti-RHDV resistance and immunity, RHDV vaccination, and the further need to establish rationally-based RHDV vaccines.
1. Introduction
Interactions between (viral) pathogens and their hosts certainly play a key role in the development of biological diversity, following an ever-ongoing ‘arms race’ in which pathogen virulence and host resistance mechanisms co-evolve with each other, leading to a dynamic equilibrium. The Calicivirus that causes rabbit haemorrhagic disease probably evolved from low-virulence strains, and is still a huge problem in the world today. Despite the fact that the disease has been circulating in the environment for about 30 years, it still has a high mortality rate [1]. The observed innate resistance of young rabbits to infections, coupled with the high fertility of the species, causes the population to recover quickly. The evolution of the resistance of rabbits to infections was also recorded [2], which together with the phenomenon of cross-protection after infection with non-pathogenic Australian rabbit calicivirus resulted in the reduction of virulence of the virus [3,4]. In turn, the virus, in order to maintain its virulence at a high level, shows the ability to use various vectors to spread over long distances, and cases of breaking the species barrier have been observed [1,3].
In the last quarter of 1983, an extremely contagious disease for European rabbits (Oryctolagus cuniculus) emerged, killing—in the following 12 months after the first outbreak—over 140 million domestic rabbits in China [5]. The disease spread in this short time over an area of about 500,000 km2. Due to the observed pathology, with pronounced damage in the liver and the severe dysregulation of the coagulation system, the disease was described as rabbit hemorrhagic disease (RHD) [5,6]. In the following decade, the virus spread throughout the globe, and threatens the state economies that are based on the rabbit industry [5,7,8,9,10,11,12,13,14,15,16,17]. RHDV had also a negative impact in Mediterranean ecosystems, were rabbits are a food base for endangered endemic species, including the Iberian eagle (Aquila adalberti) and Iberian lynx (Lynx pardinus) [18,19]. In contrast, the killing of 90% of the ‘pest’ rabbit population in Australia in very short time by RHDV was a desired impact on the ecosystem there [11,20,21,22].
The causative agent was identified as a Calicivirus [23,24,25,26,27,28]. Until 2010, all RHD outbreaks were caused by the so-called ‘classical’ RHDV (GI.1). In 2010 [29], in France, a new RHDV strain emerged, designated as RHDV2 (GI.2), which not only infects O. cuniculus but also hare species Lepus capensis mediterraneus [30], L. corsicanus [31], L. europaeus [32,33], and L. timidus [34]. In the 1990s, the classic RHDV GI.1 strains were detected in Lepus granatensis [34]. These infections in Lepus species indicate that both RHDV GI.1 and GI.2 have the ability to break the species barrier and infect other species besides O. cuniculus [34,35]. In 2020, there were new RHDV GI.2 epidemics in the Southwestern United States and Mexico, with reported mortality rates of over 90% [36] in black-tailed jackrabbits (Lepus californicus), antelope jackrabbits (Lepus alleni), desert cottontails (Sylvilagus audubonii), and mountain cottontails (Sylvilagus nuttallii). There are concerns that RHDV infection could significantly adversely affect these already endangered North American species of lagomorphs [37]. However, the pathology of RHD is similar, and is characterized by a severe clinical course and high mortality rates from 90 to 100% as a result of progressive multi-organ failure resulting from the development of DIC syndrome [38].
The clinical course of RHD appears in three forms: subacute, acute, and chronic [39,40], with respiratory, nervous and digestive disorders as a result of pathological changes in the inner organs and pronounced damage in the liver [5,41]. One phenomenon of RHD is the resistance of juvenile rabbits against a RHDV infection until the age of 4 weeks (GІ.2 strains) and 9 weeks (GІ.1 strains), respectively, in contrast to the about 3% of adult rabbits which survive after more or less severe clinical symptoms [6].
The pathology after RHDV infection is characterized by two main processes:
- (a)
- Disturbed physiology with the severe destruction of the target organs (liver, spleen, kidney) along with an extremely rapid RHDV replication within 36–72 h post infection, with up to 214 HU related to 108 copies of viral RNA per g of liver tissue [42]. Immunohistological examinations and in-situ hybridization (ISH) studies of infected adult rabbits have demonstrated the presence of RHDV in periportal hepatocytes, and macrophages of the liver, lungs, and spleen [43,44]. The disease progression is correlated to an increased apoptosis of hepatocytes and liver endothelial cells [43]. The disturbed coagulation by increased prothrombin time, increased fibrin degradation, and decreased antithrombin III activity results in micro-thrombi, DIC, and a hypervariable condition in blood coagulation [45]. Liver enzymes significantly increase, whereas antioxidant enzymes decrease within the first 36 h post infection [46]. Respiratory acidosis, hypoglycemia, and increased creatinine kinase activity indicate multi organ dysfunction, which finally results in death.
- (b)
- Disturbed immune response with an induction of systemic apoptosis of lymphocytes, and especially T-cells, leading to a nearly complete loss of the immune effectors within 48–72 h after infection. In adult rabbits, the apoptosis of T- and B-cells in the liver and peripheral blood [47,48] with the infiltration of neutrophils [49] ends in a decrease of regulatory T-cells [50] and severe leukopenia before death [51]. After RHDV infection, an increase of pro-inflammatory cytokines in the serum, liver, and spleen of young and adult rabbits [50,52], and also anti-inflammatory cytokines in adult rabbits has been reported [53].
Due to the enormous severity of the disease and the impact on the production of rabbits worldwide, several pharmaceutical companies started to develop a vaccine against this deadly viral disease (Table 1). Unfortunately, until now, no cell culture system existed which allows an in-vitro cultivation of native RHDV strains. Therefore, the first vaccines were based on inactivated RHDV prepared from the liver of RHDV-infected rabbits (Table 1) [54,55,56]. Therefore, early after the introduction of such inactivated ‘liver’ vaccines, several recombinant vaccines containing RHDV and the RHDV-2 capsid protein VP1 expressed in different in-vitro expression systems were developed [57,58,59,60,61,62,63]. The vaccines against RHDV were demonstrated to have limited cross protection against RHDV-2, and vice versa; for this reason, the current vaccines contain one or both virus-derived antigens in their formulation [64,65,66,67,68]. To date, two different types of genomic recombination have been described: one in the RHDV VP60 sequence region, and a second between the structural and nonstructural genome regions [64]. Partially non-pathogenic GІ.1 strains were detected in the late 1990s, which provide cross-protection against circulating pathogenic RHDV [69].

Table 1.
Overview of liver-derived vaccines against RHDV, licensed in Germany (PEI, state 11.01.2021).
Therefore, an in-depth characterization of the humoral and cellular immune mechanisms after RHDV infection comparing the different regulation in juvenile and adult rabbits could provide the knowledge base for rational anti-RHDV vaccine development.
2. Lagovirus Europaeus—Epidemiological History of the RHD Causative Agent
The origin of RHDV is not fully understood. The pathogenic virus forms may have evolved from avirulent strains which circulated in European rabbits asymptomatically [66,70,71,72,73] for at least 30 years before the first outbreak of RHD in China. A possible common ancestor of both RHDV and Rabbit calicivirus-like viruses from 200 years ago was predicted, which then mutated to the virulent RHDV strains which emerged in 1983 [74]. A second hypothesis is based on spillover infections of caliciviruses found in micromammals living close to wild rabbit populations [6,29,75,76]. Similarities in the clinical course and outcome after infection with European brown hare syndrome virus (EBHSV), first detected in Denmark and Sweden in 1980 in European brown hares (L. europaeus) to RHD in rabbits indicated an evolutionary relation. However, the limited sequence homology of about 76% between RHDV and EBHSV VP1 indicated that RHDV did not evolve from EBHSV [77,78,79]. This is proven by the fact that hares which survived an EBHSV infection displayed no cross immunity against RHDV [80].
The first outbreak of RHD in Europe was detected in Italy in 1986 [81] only two years after the epidemics in China and Korea in 1984 [5,82,83]. Within ten years, RHDV became endemic in Europe, resulting in a severe reduction of wild rabbit populations, especially on the Iberian Peninsula [84,85,86]. Furthermore, in domestic rabbits, RHDV caused dramatic losses in Europe as well as in North Africa [9]. In 1988, RHDV reached the American continent, with limited outbreaks in domestic rabbits in Mexico. However, due to the absence of a susceptible wild rabbit population, it was eradicated shortly after [41]. In 2000, in North America, a limited number of RHDV outbreaks was reported, and also in geographically distant regions—such as Cuba, Uruguay, and Reunion Island—RHDV caused losses in domestic rabbits [87,88]. Nowadays, RHDV is endemic in most parts of Europe, Asia, and parts of Africa.
In 1995, despite the rigorous quarantine measures, RHDV escaped from the Wardang Island in Spencer Gulf, South Australia, where the Czech V351 strain of RHDV was introduced as a biocontrol agent in 1991 [11,20,22]. Within two years, RHDV spread all over southern Australia, reducing by up to 95% the unwanted ‘pest species’ European rabbit, which is a major threat to the endemic wildlife [21,89]. A similar impact on the population of non-native European rabbits was seen in New Zealand, where the Czech V351 RHDV strain was illegally introduced [90,91]. A new RHDV strain—designated as RHDV2—emerged in 2010 in France, and spread rapidly across Europe and worldwide soon afterward as well [32].
Lagovirus europaeus belongs to the Caliciviridae family, genus Lagovirus [92]. Le Pendu et al. [93] renamed the RHDV nomenclature, and the lagovirus strains were divided into two main gene groups associated with RHDV (GI) or with the European brown hare syndrome virus (EBHSV, GII). Both, GI (RHDV) and GII (EBHSV), are closely related with regards to clinical signs, pathological and histopathological alterations, mortality rates, virion morphology, and antigenicity, but cross-species infection and cross-species protection could not be obtained for RHDV, but was found in some cases for RHDV2 [94,95]. Despite the similarities, L. europaeus GI and GII represent distinct agents infecting different species, although they cause similar diseases [40,78,80,96,97,98].
Based on genetic variability, the RHDV GI genogroup can be distinguished into 4 genotypes: GI.1, GI.2, GI.3 and GI.4. The GI.1 genotype includes the classic RHDV strains (Lagovirus europaeus/GI.1), while the GI.2 genotype was classified as RHDV2 (Lagovirus europaeus/GI.2). According to the recently-proposed nomenclature [93], GI.1 can be further divided into different antigenic variants (GI.1 a-d) based on phylogenesis and genetic distances [93]. To date, the best-tested genetic variants of Lagovirus europaeus are the GI.1a variants [47,99,100,101,102]. The main features of GI.1, G.I.1a, and G.I.2 are shown in Table 2.
Table 2.
Comparison between the main types of Lagovirus europaeus.
The GI.3 genotype is represented by RCV-E1 (Lagovirus europaeus/GI.3), and genotype GI.4 is represented by RCV-A1 (Lagovirus europaeus/GI.4) and RCV-E2 (Lagovirus europaeus/GI.4d). The strains found in these genotypes are referred to as non-pathogenic rabbit calicivirus-like viruses (RCV-like viruses). Strains from the GI.1 and GI.2 genotypes are known infectious agents responsible for RHD according to the recently proposed nomenclature [93].
The EBHSV can be divided into two groups [103]. These two groups were classified as the GII genogroup, which contains two genotypes: GII.1, which corresponds to EBHSV, and GII.2, which includes non-pathogenic strains of lagoviruses infecting hares, which are called HaCV. Additionally, variants were described among the GII.1 genotype: GII.1a (G1/group A), GII.1b (GI.3/group B), and GII.1c (G2/group B) [93].
Lagoviridae are positive-sense single-stranded RNA viruses with a non-enveloped icosahedral capsid. All RHDV virions in all genotypes share a similar structure: the single subunit capsid protein VP1 forms the virion, the minor structural protein VP2 is responsible for stability after the incorporation of viral RNA, and the so-called VPg (viral protein genome-linked) is essential for virus replication [104]. The capsid protein VP1 is the main viral antigen, as it is exposed to the humoral response of the host due to the lack of an envelope. Furthermore, the C-terminal sequence of this protein is exposed on the external surface, and is mostly responsible for the antigenic variability within this virus family [105]. VP1 induces high titres of virus-neutralizing antibodies in surviving hosts, and displays the antigenic variability within this virus family [13,105,106,107]. Because of the high variability between the genotypes, cross protective immunity is induced at a limited level only. This was also found in several studies in which vaccines against RHDV GІ.1 strains provided only limited cross protection, and vaccines against RHDV GІ.2 provided even less [108,109,110,111]. Due to its high variability and recombination ability, the evolution within the RHDV genogroups is ongoing. In addition, it is likely that new strains with different virulence will appear. Whether or not recent vaccines will induce an immunity which will also protect against new emerging strains is at least questionable.
3. Host-Pathogen Interaction in Rabbits after Infection with RHDV (Lagovirus europaeus/GI)
Infection with RHDV causes very rapid, severe organ destructions in the inner organs, especially the liver. Moreover, an immune pathological process starting with a polyclonal activation of the lymphocytes leads to dramatic losses of these immune effectors, and finally a multi-systemic, multi-organ failure, and the death of infected host [6]. This extremely rapid disease progress is even more impressive given the fact that very low doses are sufficient to infect a naïve host [112].
The virus is transmitted mainly orally, but also by nasal, conjunctival, or parenteral routes, directly between animals and indirectly by insects, which take up the virus from the excretions of infected animals [113,114,115]. There is an extremely short time (24–60 h post infection) between infection with very low doses in peripheral mucosal surfaces and the overwhelming viral load in inner organs’ requests for an initial replication in these peripheral epithelia, and furthermore, for innate immune mechanisms which might be decisive for survival.
An important aspect of the characterization of RHDV infections (Lagovirus europaeus/GI) is its inability to cause severe disease in juvenile rabbits younger than 9 weeks (RHDV GІ.1 [113]) or younger than 4 weeks (RHDV GІ.2) [116] compared to adult rabbits. This is especially remarkable in respect to the immature immune system of juvenile rabbits, especially of the B-lymphocyte ontogeny and maturation [117], which request innate immune mechanisms preventing the initial high replication of RHDV, or which regulate adaptive immune mechanisms to block RHDV replication in the inner organs, especially the liver. The regulation towards T-effector cell-mediated immune mechanisms in juvenile rabbits might be similar to rare cases of adult rabbits surviving the RHDV infection, in which a less severe T-cell loss is seen compared to moribund rabbits [102,118]. The survival of adult rabbits seems to be positively correlated to an IFNγ-blocking of viral replication [53,55,119]. The following B-cell response ending up in high titers of RHDV neutralizing antibodies can protect against future reinfection, but is not responsible for survival—as it takes too long—but rather for long term protection [120,121]. The question is whether the innate effector and/or regulatory mechanisms in juveniles and adult rabbits surviving an RHDV infection are different. The reasons for the two different resistance patterns after RHDV infection in resistant juveniles and fully-susceptible adults were investigated based on different suggestions:
- (a)
- Missing virus receptor in peripheral mucosal surfaces.Juvenile rabbits do not express, or only express in incomplete formation, one identified attachment factor for RHDV, the so called HBGA (histo-blood group antigens) in the upper respiratory and digestive tract until the age of 9 weeks [122]. However, entry into hepatocytes is not provided by HBGA because they are not expressed at any age on hepatocytes [122]. Moreover, even in juvenile rabbits, RHDV was detected after infection in the liver [123,124]. Finally, immunosuppressed juvenile rabbits develop a full disease state as adults, indicating that immune mechanisms are involved in the juvenile resistance against RHDV [125].
- (b)
- Different replication kinetics in peripheral mucosal surfaces.Flies or direct contact transfer RHDV via the oral/oculo-nasal aerogenic route through the upper respiratory tract with an extremely low dose of possibly less than 100 viral particles [112]. However, between 24 and 48 h post-infection, RHDV replicates to very high titers in the liver of adult rabbits, whereas the amount of RHDV in the liver of juveniles remains low [124]. Whether this is due to limited virus replication in peripheral mucosa in juveniles or due to more effective type I or III IFN responses in juveniles compared to adult rabbits is not clear.
- (c)
- Different effective innate immune responses.The pathological processes (see above) leading finally to a DIC are described for both juvenile and adult rabbits [124,126]. Protective immune mechanisms preventing severe clinical processes are mostly developed in juvenile rabbits, but rarely in adults, The more robust innate immune response in juvenile rabbits was characterized as one reason for the RHDV resistance in a transcriptome analysis-based study [127]. The involvement of cellular immune mechanisms, and especially of CD8+ T-cells in adult rabbits in protective responses were elucidated [111,128].
- (d)
- Different regulation of the Th1/Th2 balance in juvenile and adult rabbits.In surviving adult rabbits, an early and strong increase of IFNγ as well as a limited loss of T-cells was determined. The course of the disease is less severe with a reduction of the viral replication in the liver, an increase of CD8+ T-cells, and an increase of IFNγ as well as type I IFNs [53]. Similar response patterns are described for juvenile rabbits where macrophages, T- and B-cells increased in the liver and spleen [52,123]. The reasons for the ‘decision’ towards Th1-based immune responses are different in adults and in juveniles. In juveniles, it might be based on their immature B-cell development [117], as the different outcome after RHDV infection is related to B1 cells contributing to innate immune responses, and not B2 cells as part of the adaptive immune response [129].
However, neither the regulation towards immune mechanisms, which provides final protection, nor the cell populations involved in such early regulatory processes are characterized yet. Consequently, whether the regulation at immune checkpoints towards protective Th1 immune mechanisms and the immune cells inducing and regulating these processes are the same in juvenile and in adult rabbits is also unclear.
3.1. Innate Immune Mechanisms Involved in the Anti-RHDV Immune Response of Juvenile and Adult Rabbits
In naïve animals which encounter a viral pathogen for the first time, an effective innate immune response is in most cases decisive for survival or severe disease and death. This is provided either by the complete blocking of viral replication, and afterwards clearance at the peripheral epithelia, or by decreasing the viral replication for a certain time to a level that—following adaptive immune mechanisms—can reach a level to clear the viral pathogen.
3.1.1. Pathogen Sensing
The immune response to virus infection is initiated when the PRRs (pathogen recognition receptors) of the host cells recognize specific non-self pathogen-associated molecular patterns (PAMPs) which introduce innate immunity by triggering intracellular signalling events [130,131].
PRRs include, among others, toll-like receptors (TLRs) located on the cell surface, as well as nod-like receptors (NLRs), type-C lectin receptors (CLRs), RIG-I-like receptors (RLRs) and Pyrin-HIN domain name receptors (PYHIN) (Figure 1). PRRs are responsible for the recognition of PAMPs, and moreover DAMPs—damage/danger-associated molecular patterns which occur through infection-induced cell losses. The activation through these sensors initiates the production of cytokines and chemokines in the host organism, which activates NFκB and IRF-3 factor, and finally leads through type І IFN and proinflammatory cytokines (IL-1, IL-2, 1L-6, or IL-12 or TNF-α) to an antiviral status [53,132,133].
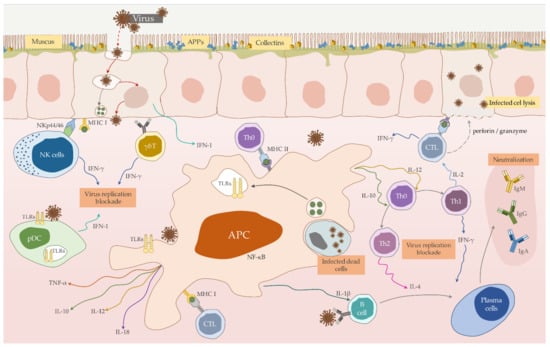
Figure 1.
Schematic representation the mutual interaction of immune cells proven or assumed to be involved in the immune response against Lagovirus europaeus infection.
In the case of RHDV, the incubation period is 1 to 3 days, i.e., very short, and rabbits usually succumb within 12 to 36 h after the onset of fever (>40 °C). The rapid and fatal course of the disease suggests that the rabbits usually die before mounting an effective adaptive immune response against RHDV infection, and moreover that innate virus replication-blocking immune mechanisms are obviously decisive for survival or death.
Concerning rabbits, the general knowledge about PRR is very limited. Only TLR1 [134], TLR2 [135], TLR3 [136], TLR4 [137], TLR8 [138], and TLR9 [139] (GenBank Accession number: KC349941, NM001082781, NM001082219, DQ250128, AY101394, JX948743, HM448400) have been identified and sequenced until now. Further putative TLR sequences of rabbits have been identified but not yet functionally characterized (TLR5 Acc. Number: HQ874605, TLR6 Acc. Number: KC349945, TLR10 Acc. Number: KC349942, TLR11 Acc. Number: KC349943, TRL12 Acc. Number: KC349944). However, the complete sequence of TLR1 is known [134], but might be of lesser importance in viral infections of rabbits, because it is known to have a more antibacterial effect. TLR3 was identified by recognizing double stranded viral RNA, and was shown to be very diverse among Oryctolagus cuniculus [140]. In total, 41 single nucleotide polymorphisms (SNPs) in the sequence of TLR3, and some substitutions, were observed within the LRR motifs in the TLR3 molecule [140]. Only TLR4 was shown to play a role in a RHDV infection. It was activated during RHDV infection through binding of the high mobility group box 1 protein (HMGB1), which is secreted by immune cells during inflammation [141]. The receptors known to recognize single stranded viral RNA, TLR7 and TLR8, were found to be absent or were previously pseudogenised [136] in rabbits. However, it was shown later that TLR8 is a functional gene, but with lower activity than in other mammals [138]. TLR9 is preferentially expressed in immune cell-rich tissues. However, the involvement in immune responses against real bacterial pathogens is not yet shown [139]. In general, the involvement of TLR pathways in the recognition of RHDV and the possible influences between resistant juveniles and fully susceptible adult rabbits are missing, and should be definitely explored further.
Among NLRs, studies are rare; only a few reports are known about the presence of NOD-like receptors in rabbits, for example rNLRX1 in the infection of rabbits with enterohemorrhagic Escherichia coli [142]; nevertheless, these kinds of receptors are also described to take part in bacterial infections more often than viral infections [130]. Interestingly, only one report exists about the diversity of RIG-I-like receptors among leporids (Oryctolagus, Sylvilagus, Lepus), which were shown to be involved in immune responses against myxomatosis [143].
3.1.2. Apoptosis and Autophagy in RHDV Infection
The major histopathological lesions found at necropsy are acute hepatitis due to liver cell loss as the result of RHDV-induced apoptosis, and splenomegaly. Haemorrhages and congestions can be seen in several organs—particularly in the lungs, heart, and kidneys—as a result of a massive disseminated intravascular coagulation (DIC), which is usually the cause of death [45].
The process of programmed cell death plays one of the key roles in the pathogenesis of RHD, and affects various cells. Apoptosis in rabbits infected with different strains of the RHD virus was first detected using histopathological methods, i.e., 40 h p.i. in hepatocytes [43]. In addition, the presence of apoptotic cells in RHDV (Lagovirus europaeus/GI.1) infection has also been confirmed in macrophages of the abdominal cavity, epithelial cells, lungs, kidneys, heart, spleen, and lymph nodes [43,49]. More detailed studies on apoptosis in immune cells after RHDV virus infection were performed using six RHDV (Lagovirus europaeus/GI.1) hemagglutinogenic strains (24V, 1447V/96, 01-04, 237/04, V-412, 05-01) and three non-hemagglutinogenic strains (Rainham, Frankfurt, and Asturias). This showed that apoptosis in the granulocytes and peripheral blood lymphocytes is activated from 4–8 h p.i. in the case of non-hemagglutinogenic strains, and from 12 h p.i. with hemagglutogenic strains [47]. This process intensifies in both cases up to 24–36 h p.i. Apoptosis was more intense in lymphocytes after infection with both hemagglutinogenic and non-hemagglutinogenic RHDV (Lagovirus europaeus/GI.1) strains [47,48]. Autophagy is also activated during RHDV (Lagovirus europaeus/GI.1) infection. This process weakens as the disease develops, which in turn is accompanied by an increase of programmed cell death. This phenomenon probably shows a protective effect of the organism in response to virus infection. Therefore, further research is necessary in order to understand the process of autophagy in RHDV (Lagovirus europaeus/GI.1) infection, and the influence of different virulent RHDV strains [126].
The first study on the phenomenon of the apoptosis of peripheral blood granulocytes and lymphocytes in rabbits infected with RHDVa (Lagovirus europaeus/GI.1a) strains—including three hemagglutinogenic strains (Triptis, Hartmannsdorf, Vt97) and two non-hemagglutinogenic strains (9905 RHDVa, Pv97)—showed that, for all of the strains tested, apoptosis is expressed through changes occurring in the percentage share of apoptotic granulocytes and lymphocytes. These changes were recorded at 4–8 h p.i. and 24–36 h p.i. [144]. Moreover, the frequency of apoptotic changes was higher in the case of apoptotic granulocytes, and the average number of these cells remained ten times lower compared to apoptotic lymphocytes [144]. A number of studies show that during RHDVa infection a different immunological picture is observed than in RHDV, and these differences relate primarily to specific cellular and humoral immunity and the process of apoptosis [47,102,145]. These studies show that the biological features of RHDV GI.1 strains (hemagglutination capacity, antigenic variability) have an impact on the immune response.
To date, studies on apoptosis and autophagy in rabbits infected with genotype RHDV GI.2 strains have not been conducted. A better understanding of the different immune processes after infection with different RHDV genotypes in relation to the different outcome of such infections in juvenile versus adult rabbits is certainly a key element for rationally-based vaccine design.
3.1.3. Apoptosis and Autophagy in RHDV Infection
An important aspect to understand the different immune responses in naïve juvenile versus adult rabbits to RHDV infections is based on the characterization of cytokine induction and regulation. Several studies have indicated that cytokines play an important role in modulating the immune response through the activation of peripheral blood leukocytes and their recruitment to the liver (Figure 1). An increase of tumor necrosis factor alpha (TNF-α), interferon alpha (IFN-α), interferon gamma (IFN-γ), transforming growth factor beta (TGF-β), and interleukin-1, -6, -8, and -10 has been observed [50,146]. In the liver, a decrease of hepatocyte growth factor (HGF) expression, which is responsible for liver regeneration, and an increase of the activity of TNF-α, TGF-β, and IL-1β and -6 was measured [46,147,148]. The analysis of cytokine patterns in peripheral blood leukocytes showed a decrease in IL-1β, -2, and -18 activity, and an increased expression of IL-6, -8, -10, TNF-α, and TGF-β genes [53,133]. It was speculated that TNF-β, similarly to TNF-α, which extends the life of infected rabbits by 8-12 h, can act in the same way [149].
Furthermore, the analysis of the gene expression pattern of key elements in the regulation of innate immunity indicated that the different resistance of young rabbits and adults after RHDV GI.1 or GI.2 infection is based on these early recognition and regulation processes [127]. In response to RHDV GI.1, particularly, genes of the major histocompatibility class I complex, as well as interferon-induced genes, were upregulated in juvenile rabbits, accompanied by an increased activity of natural killer cells and CD4+ T-cell activation. These phenomena could be important for the resistance of juveniles [127]. In contrast, after RHDV GI.2 infection, these were downregulated, which correlates with the limited resistance of juveniles older than 4 weeks of age. Interestingly, in adult rabbits, MHC (major histocompatibility complex) class II genes were upregulated, indicating an induction of a Th2 immune response known to be unable to protect naïve rabbits because of the rapid disease progress [127].
Possibly, these pathological processes impacting mainly the liver and the cytokine storm may be the direct cause of disseminated intravascular coagulopathy [126,150].
3.1.4. Innate Effector Component and Functions
In the course of RHD, several innate cellular effectors provide the body’s first and primary line of defense in contact with viral pathogens [130].
Neutrophils (polymorphonuclear cells, PMNs) are crucial in innate immune responses due to their cytotoxic, phagocytic and neutrophil extracellular trap forming (NET) activities [151] after the recognition of PAMPs by TLRs. Moreover, neutrophils produce active substances such as lysozyme (LZM), myeloperoxidase (MPO), elastase, esterase, or defensins, which provide anti-viral activities [151,152]. In two studies, the activities of PMNs were measured in the early course of RHD by a variety of parameters (neutrophil phagocytosis, PMN adherence capacity, spontaneous and stimulated metabolic activity, etc.). Most of the parameters indicating functional activation were increased as early as 4–8h p.i. [153]. Interestingly, after infection with Lagovirus europaeus/GI.1a, the same parameters decreased, starting at 8–12 h p.i. [101,152], suggesting a different regulation of innate immune mechanisms depending on the genotype of the virus which is probably responsible for the rapid deaths of rabbits in these studies 24–36 h p.i. after RHDVa infection.
Furthermore, during RHDV (Lagovirus europaeus/GI.1) infection, a polyclonal hyperactivation in immune cell populations was observed, leading to a recruitment of immature immune cells from the bone marrow within the final stage of RHD before death. Neutrophils display cytoplasmic vacuolization, toxic granules, and Doehl’s bodies as a sign of a systemic dysregulation within the hematopoietic tissues. Macrophages were present in increased amounts, showing the vacuolization of the cytoplasm and phagocytized erythrocytes, lymphocytes, and platelets [146]. In order to characterize the dysregulation of the different functional activities of the innate effector cells, further studies investigating the regulatory gene networks are needed.
The role of natural killer cells which are involved in immediate cytotoxicity against virus infected cells has not been well investigated, yet. In a gene expression study comparing the response of juvenile and adult rabbits to infection with RHDV GI.1 and GI.2 strains, the genes involved in activation and function of NK cells (PTPN22, VAV1, and ARRB2) were downregulated in GI.2.-infected juveniles, while they were upregulated in GI.1-infected juveniles which survived the infection, compared to adults infected with GI.1. [127]. Also, the natural cytotoxicity triggering receptor 3 (NCR3) was elevated. This indicated that, in naïve rabbits, the activation of innate effector cells is crucial for survival.
3.2. Antigen Presentation
Virus particles are sensed by pathogen recognition receptors (PRRs) on the surface [154,155,156] or in the endosomes [157] of myeloid cells, mainly dendritic cells (DC). The uptake of such particles activates DC, which afterwards mature. Mature DC process and present peptides derived from viral proteins loaded into major histocompatibility complex (MHC) class I or class II molecules to the T-cell receptor complex of CD8+ or CD4+ T-cells, respectively. It is worth mentioning that the CD4 gene shows an overall higher divergence in lagomorphs in comparison to primates, with the highest divergence in the D2 domain [158]. The interaction between DC and T-cells leads to an activation and proliferation followed by adaptive effector function via Th1 and/or Th2 responses. Ideally, a Th1 response limits the viral replication by cytotoxic CD8+ T-cells which lyse virus-infected cells or abrogate the viral replication through IFNγ-promoted Type I IFN responses. A Th2 response helps B cells to produce virus neutralizing antibodies which act mainly through the blocking of the viral epitopes which are necessary to interact with virus receptors [159,160].
However, in the ‘battle for survival’ between viral pathogens and vertebrate hosts, this response is often counteracted by viral proteins which are able to block the intracellular processing and presentation machinery in DC at different stages [161].
3.2.1. DC’s Interaction with RHDV
Although the immune response of naïve rabbits against RHDV infection is widely studied, only limited data are available on whether the viral replication also happens in myeloid cells (monocytes, macrophages, DC). RHDV VP1 was detected during infection in Kupffer cells in the liver, as well as in circulating monocytes, lymphocytes, and macrophages in the red and white pulp of the spleen, lung macrophages, the glomerular mesangial cells of the kidneys, and lymphocytes in the thymus and lymph nodes [162,163,164]. Whether this indicates an active replication in these cells or whether RHDV VP1 was detected after the uptake of other infected cells is not clear. However, the detection of RHDV in Kupffer cells was connected with signs of apoptosis [43].
3.2.2. Antigen Processing
Obviously, in adult rabbits, the initiated Th2 immune response after infection is counteracted by RHDV through an apoptosis induction in T-cells [6,127]. Whether DC at the peripheral mucosal surfaces or Kupffer cells in the liver are able to process and present RHDV VP1 epitopes is not clear. In juvenile rabbits, an increase of MHC class II early after the infection with RHDV GI.1 was connected with an upregulation of CD4+ gene activity, and also with an increased survival rate in contrast to RHDV GI.2-infected rabbits [127]. The extremely rapid pathological progress in the liver normally leads to death before a successful induction of adaptive Th2 immune mechanisms can occur.
However, in the rare cases of the survival of naïve RHDV-infected adult rabbits, an effective CD8+ T-cell response protects the liver from severe damage by the secretion of a high level of IFNγ. Moreover, a strong B-cell response provides high titers of RHDV neutralizing antibodies [102,118]. Both indicate that an effective processing and presentation of RHDV VP1 epitopes via MHC I and MHC II by DC occurred.
In contrast to the severe course of RHD in adults, in juvenile rabbits a completely different outcome is seen after infection with RHDV, which is not only indicated by survival but by a complete blockage of the viral replication in the inner organs [6]. An early and very effective type I IFN response might be decisive for this resistance. However, there are also differences in antigen presentation compared to adults, which are moreover different after infection with GI.1 or GI.2 RHDV strains. After GI.1 RHDV infection, the antigen presentation leading to an effective CD8+ T-cell response via MHC I as well as the MHC II pathways triggering mainly the B-cell response are upregulated. After infection with GI.2 strains which are able to infect and kill juveniles older than 4 weeks, the MHC II pathways are downregulated. This different activation of antigen presentation pathways might be the reason for the different resistance of juvenile rabbits [127].
3.3. Adaptive Immunity
The mechanisms of acquired adaptive immunity evolved in early jawless vertebrates with the appearance of variable lymphocyte receptors (VLR) comprised of leucine-rich-repeat (LRR) segments [165], and further in jawed vertebrates with the immunoglobulin receptor gene family and the recombination activation genes in B- and T-lymphocytes [166]. These effector cells create a second line of defence after the recognition of variable epitopes in pathogen-derived proteins. This second line of defence is activated when the innate immune mechanisms cannot completely clear an infection due to the high number or virulence of the invading pathogens. However, the activation of adaptive immune mechanisms towards a ‘new’ pathogen in naïve hosts needs 1–2 weeks to reach full efficacy. The disadvantage of this late first adaptive response is afterwards turned into the advantage of quicker and more effective response due to immunological memory by circulating antigen-specific antibodies and specific B- and T-memory cells [167]. The adaptive immune response evolved as a complex regulatory network between the humoral factors and immune cells of both early innate and later adaptive immune mechanisms. The responses start with the presentation of pathogen-derived peptides presented via MHC class I, mainly at peripheral mucosal epithelia, or MHC class II by antigen presenting cells (APC) at peripheral or in systemic lymphoid tissue (Section 3.2). The presentation via MHC I or MHC II determines the adaptive response through Th1 regulation towards cellular, mostly CD8+ T-cell, permitted responses or Th2 regulation towards humoral antibody secretion by activated B-cells.
In naïve animals, the survival after infection with virulent RHDV depends in the first instance on the effective innate immune mechanisms simply because of the time delay of between 3 and 7 days until adaptive immune effectors, activated B-cells, or activated cytotoxic T-cells fully functionally respond after the first pathogen recognition. After the infection of juvenile rabbits with virulent RHDV strains, a robust innate immune response prevents completely the development of RHD by limiting the viral replication. However, in adult rabbits, the innate mechanisms mainly fail, and the extremely rapid viral replication leads to severe RHD and high mortality. The different innate immune response in juvenile and adult rabbits involved in resistance and survival against RHD infection then determines a different adaptive immune response. Finally, the immunological memory can differ in response to infection or vaccination with regard to specific T- and B-lymphocytes. This is also seen in rabbits surviving a RHDV infection or after vaccination by the high immunogenicity of RHDV VP1. Overall, the development of adaptive immune responses after infection is always interfered by pathogen-induced counteraction. All of these aspects will be discussed in this paragraph.
3.3.1. Humoral Immunity to RHDV
Calicivirus infections in vertebrate hosts most widely trigger a humoral response, which is involved in infection control through the limitation of virus propagation and the systemic spread of the virus [121,161,168,169]. In rabbits, after RHDV infection, the antibody response is different either between naïve and immune rabbits (survivors of infection or after vaccination), and between juveniles with mostly immature B-cells and adults with a fully mature immune system [6,111,128]. Moreover, the structure of RHDV particles is partially different from other caliciviruses. Mature virions are composed of the VP1 protein building a spherical, non-enveloped icosahedral capsid of 32 to 44 nm. The VP1 that includes the N-terminal arm (NTA), the shell (S), and the protrusion (P) forms in this particle characteristic cup-shaped depressions [170,171,172]. A specific P2 sub-domain of RHDV displays high variability in different RHDV isolates, which contributes to HBGA binding differences. Furthermore, in the most exposed surface loop—L1 (a.a. 300–318)—of VP1, a high variability was detected between different RHDV genotypes. This region contains the neutralizing antibody inducing epitopes [106,107].
As discussed above, in naïve animals the Th2 regulated B-cell response leading to virus-neutralizing antibodies does not provide protection against primary RHDV infection, mainly due to the extremely rapid RHDV replication, the induced T-cell apoptosis, the severe liver destruction by DIC, and due to the time delay between infection and the first appearance of effective anti-RHDV antibody titers (Figure 2). On the other hand, the first RHDV-specific IgM was detectable as early as 50 to 60 h p.i., and is reliably detected at 72 h p.i. [173]. Whether RHDV specific IgM is involved in the decline of the RHDV load in the liver detected at about 50h post infection with virus-capture ELISA [11] is not clear. RHDV particles isolated from rabbit tissues 144 h p.i. were not able to infect naive rabbits, possibly due to the formation of immune complexes or the degradation of viral particles [120].
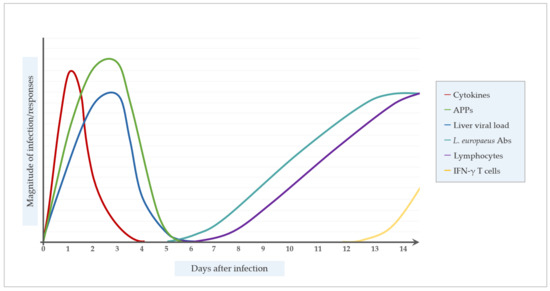
Figure 2.
Temporal dynamic of the immune response after Lagovirus europaeus infection.; APPs, acute phase proteins; L. europaeus Abs, Lagovirus europaeus antibodies.
On the other hand, in surviving rabbits, very high titers of RHDV-neutralizing antibodies not only protect from a severe clinical course and death but also prevent the viral replication in inner organs. Moreover, repeated infection always increases such titers very effectively, which presumably leads to a lifelong immunity [6,11].
The percentage of B lymphocytes (determined as CD19+) increased constantly from 8 h to 52 h post infection [99,102,145], followed by high IgM within 2 weeks.
The anti-RHDV IgG which increased slower persisted for months [111]. Interestingly, a repeated vaccination 21 days after the primary one reduced the persistence of protective IgG [111]. The virus-neutralizing capacity of anti-RHDV antibodies was proven by the passive immunization used to stop an RHD outbreak in a rabbit farm [121]. The induction of IgA titers in naïve rabbits which survived an infection was measured in field tests in Australia. Whether these antibodies already prevent infection at peripheral mucosal surfaces is not clear [120].
Three aspects in the antibody response of naïve, juvenile or adult rabbits to a RHDV infection have to be distinguished.
Can naïve young rabbits below 8 weeks of age display a high affinity antibody response despite their mainly immature B-cells? The diversification of the VDJ (variable diversity joining) repertoire seems to start after birth, and it is completely similar to an adult’s by 2–3 months of age [117]. The antibody responsiveness of rabbit kits to immunization also follows the same dynamic. The advantage or disadvantage behind this delayed diversification is not known. However, there are not many diseases that have been reported to affect rabbit kittens, and the resistance to RHDV GI.1 strains is one example of the ability of young rabbits of 4 weeks of age to mount high titers (up to 1.640) after infection [121]. These titers correlated with an increase of B-cells in the liver and spleen, and a robust pro-inflammatory cytokine response. This paradigm changed just recently, when RHDV-2 spread out. However, an involvement of the induced specific antibodies in the immediate clearance of RHDV very early after infection and the protection of the hepatocytes from being infected seems not to be reliable.
Do maternal antibodies from immune mothers influence the antibody response in their offspring? In general, maternal antibodies provide protection from a severe clinical course after infection [174]. Rabbits have a haemochorial placentation, in which maternal antibodies are transmitted from the mother to the offspring through the placenta [175]. In wild rabbit populations, maternal antibodies seem to be important for protection against RHD even after the natural juvenile resistance decline [120,176]. After a single-dose vaccination, RHDV-specific maternal antibodies protected offspring up to the fourth generation. They were mainly transmitted in gestation through trans-placental transmission, and only partly in the lactation period. These maternal antibodies passively protect offspring for at least the first 28 days of life [177]. Specific studies on whether those RHDV-specific maternal antibodies interfere with an active immunization has not yet been studied in detail. However, in general, maternal antibodies attenuate the response to vaccination; especially, the induction of virus-neutralizing antibodies can be reduced. Rabbits have two Cγ gene segments with identical nucleotide sequences, which means that they have only one IgG subclass [178,179], and so no restriction of the IgG subclass is expected during maternal transfer. The stimulation of the immune system of the offspring is not hampered by maternal antibodies, and rabbits with induced pathogen-specific antibodies (IgG + IgA) can be clearly distinguished from rabbits protected from maternal ones (IgG) [120]. The persistence of those antibodies, however, is limited, especially after vaccination [177].
Is the antibody pattern in adult rabbits surviving a RHDV infection different from that of RHDV-resistant juveniles below 8 weeks of age in which a systemic RHDV replication is prevented by a robust innate immunity? Rabbit B cell lymphopoiesis initially occurs in the fetal liver [180,181]: after birth, the bone marrow is the main resource of pre-B-cells, in which they comprise 10–20% of all hematopoietic cells. However, until 8–9 weeks of age, the number of pre-B-cells declines constantly to negligible levels. Moreover, at the age of 4 months, no immature B-cells are present in the bone marrow anymore [182]. However, this time-limited B-lymphopoiesis in bone marrow seems to have no negative impact on the ability of adult rabbits to mount an effective high-affinity antibody response. The general usage of young adult rabbits (6 months of age) for the production of highly-specific antisera indicates the potential of the B-cell repertoire. After RHDV infection, these responses are obviously different in juveniles (<9 weeks of age), with an ongoing B-cell ontogeny from adult rabbits with a fully mature immune system.
3.3.2. Cellular Response
Viral infections are not only controlled by specific virus-neutralizing antibodies in CD4+ T-cell-regulated Th2 immune responses but also by CD8+ T-cells which, after activation, either lyse virus-infected cells or induce by the secretion of IFNγ a complex network which blocks the viral replication in infected cells without cell destruction. This is very important in metabolic organs like the liver, where massive cell damages lead to immunopathological organ destruction. In RHD, such a process is involved in the fatal outcome in adult rabbits.
The cellular immune responses to RHDV infections are as different as the outcome between juvenile and adult rabbits. In juveniles (<9 weeks of age) no significant changes in the number of T-cells were detected after infection with RHDV GI.1 strains. However, the immunopathology is comparable after infection with GI.2 strains with a remarkable lymphopenia, especially of T-cells [47,49,51,183]. In adults, the apoptosis of lymphocytes is one main pathological process after both RHDV GI.1 and GI.2 strains [6,184]. Especially, the number of CD4(+)Foxp3(+) regulatory T cells (Tregs) decrease in infected adults, whereas in juvenile rabbits the number and frequency of splenic Tregs did not change [50]. Interestingly, the impact on T-cells varies after infection with hemagglutinating or non-hemagglutinating strains. The percentage of all T lymphocytes (CD5+), as well as of CD4+ Thelper cells and CD8+ Tcytotoxic cells, decreases after infection with HA- strains, but increase slightly after infection with HA+, starting 4–8 h p.i. [47,48,144]. In RHDV GI.2 infection, an initial decrease in CD4+ and CD8+ T-cells is observed, followed by a progressive apoptosis in moribund rabbits, and by recovery and proliferation in surviving adult rabbits. This indicates an involvement of CD8+ T-cells in protective responses in naïve adult rabbits [111].
The recruitment of different leukocyte populations to the RHDV-infected liver has been shown to be decisive for the outcome. In adult rabbits, mostly heterophils infiltrate the liver, and appear there near hepatocytes showing severe cellular damage. In contrast, in young rabbits, lymphocytes in close membrane contacts with the cell surface of undamaged hepatocytes dominate [185]. The importance of T-cells in the protection of the liver from severe pathological processes and fatal outcomes after infection was proven in young rabbits that lost their natural resistance after immunosuppressive treatment [125].
3.4. Immune Response Pattern Protecting Naïve Rabbits—Lessons to Learn for Vaccination
To summarise the findings about protective immune responses in naïve rabbits against RHDV infection, different patterns are seen in juveniles and adults.
The robust innate immune system based on type I IFN-regulated gene networks limits the RHDV replication already in the periphery, and prevents a systemic infection [125,127,186]. The adaptive immune response, afterwards, is mainly based on neutralizing antibodies, which provide a long-term immunity. In the field, the titer of anti-RHDV antibodies increases strongly after repeated contact with the virus, leading to (presumably lifelong) immunity against homologous or closely-related RHDV strains [187,188]. However, the rapid evolution of RHDV counteracts the antibody-based immunological memory [109,189,190,191,192].
In adults, an extremely rapid RHDV replication overwhelms possible innate responses in the periphery, ending up in a rapid systemic spread and infection of inner organs. The induced Th2 immune responses are not able to reach a level which limits the RHDV-induced pathology, with a fatal outcome. However, as-yet not understood early immune regulatory events direct the immune response to Th1-based mechanisms with the activation of CD8+ T-cells, which seem to be able to control the RHDV replication in the liver without destructive cytotoxic activities. In these rare cases, the integrity of the liver is retained, and the infected rabbits survive the infection with less severe clinical signs.
A vaccine, which would trigger the immunoregulatory mechanisms behind these two patterns, might help to eradicate RHDV, because the neutralizing antibody-induced viral evolution is accompanied by cellular immune mechanisms which strictly limit the release of new RHDV variants.
4. Vaccines—From Inactivated RHDV-Infected Liver Preparation to Recombinant Vaccines
The extremely high mortality in naïve rabbits after RHDV infection, along with the enormous impact on commercial farmed rabbits and pet rabbits, and finally on wild rabbits in ecosystems lead to high activities in vaccine development. Unfortunately, all of the attempts to establish an in-vitro cell culture system to produce RHDV vaccines failed. Moreover, the emergence of many RHD epizootics and the variability of RHDV increased the need to develop an effective vaccine to limit the expansion of the virus.
4.1. Conventional Vaccines against RHDV
Shortly after the emergence of RHDV GI.1, several inactivated RHDV vaccines based on preparations of liver material from RHDV-infected rabbits were developed [54,55,56]. So far, no standard RHDV strain for vaccine development has been established. Therefore, usually the most virulent, local RHDV strains are selected as vaccine strains. All of these inactivated vaccine candidates were tested in ‘classical vaccination-challenge trials’, and the high immunogenicity of inactivated RHDV was proven. In these studies, the induction of virus-neutralizing antibodies was shown, lasting mostly for more than 12 months post vaccination. All of these vaccines not only prevent severe clinical courses of RHD and high mortality but also inhibit RHDV replication [54,56]. However, after the emergence of RHDV2 in 2010, new vaccines had to be developed, as these classical vaccines were shown to be very effective in inducing long term protection against homologous RHDV GI.1 strains, but provided only limited cross protection against GI.2 genogroup strains [193]. The shortly-thereafter developed liver-derived RHDV2 vaccines (FILAVAC VHD K C+V, Filavie S.A.S; ERAVAC, Hipra) also conveyed only limited cross protection to other RHDV strains, so that rabbits need to be vaccinated either with two different vaccines or with polyvalent vaccines, leading to higher economical efforts for breeders. Moreover, the observed variability of RHDV with the potential emergence of further virus variants showed the limitations of a vaccine development strategy based on liver preparations from infected rabbits.
4.2. Recombinant Vaccines against RHDV
In order to improve vaccine production methods using the liver material of infected rabbits, numerous alternative approaches were developed in the last few years. However, the induction of general protection against RHDV is not the sole focus. Several claims have been made in regard to a recombinant vaccine. A vaccine should be ethically justifiable, induce protection shortly after administration, induce long term protection, convey cross protection against several RHDV strains, be economical, and ideally be administered only once during a rabbit’s life.
In order to achieve some or all of these goals, since the 1990s, many recombinant vaccines against RHDV have been developed [58,59,194]. In most cases, the focus was on the expression of the capsid protein VP1, as it was shown to be very immunogenic [58,105]. Recombinant VP1 was expressed using different expression vectors, such as baculovirus [58,111,128,194], adenovirus [195], myxoma virus [60], yeast [196], Escherichia coli [197], vaccinia virus [61], insect cells [59], or plants [198].
One desired effect of VP1 expression is the spontaneous formation of virus-like particles (VLP). Because of their similarity to infectious virus particles, they have more immunogenic potential than non-assembled VP1, and therefore can elicit a strong humoral and cellular immune response [199,200,201]. VLP are taken up by antigen-presenting cells, and are presented by MHC class II molecules, leading to the activation of DC, cytokine release, and CD4+ T-cell activation, as well as by MHC class I molecules, leading to CD8+ T-cell activation [200,202]. Due to their small size, VLP can spread to lymph nodes to interact with even more CD4+ T-cells. In addition to a potent cellular immune response, a humoral immune response is also triggered by the activation of B-cells, leading to high antibody titers and the generation of memory B-cells [200]. In the few last years, vaccines based on VP1 VLP have therefore been brought into focus [111,128,197,203].
So far, to our knowledge, only two recombinant vaccines have come to market (NOBIVAC® Myxo-RHD PLUS + NOBIVAC® Myxo-RHD, MSD Animal Health), with only one conveying protection also against RHDV2. Both are based on a myxoma virus vector expressing RHDV/RHDV2 VP1.
4.3. New Approaches in Vaccine Design
Classical vaccine development approaches concentrate on the administration of antigens, which are ideally very immunogenic, in order to induce a specific and hopefully long-lasting immune response and therefore protection against the pathogen. With many vaccines, however, the duration of protection is often limited to only a few years at most. Additionally, in order to prolong protection, adjuvants are needed for the enhancement of the immune response. Furthermore, vaccines need to be boosted over the years in order to maintain protection. Classical vaccines are often developed against ‘easy pathogens’ based on the use of the structural proteins of those pathogens, which are known for high immunogenicity. However, the challenges of modern vaccine development lie more with pathogens that hide from the immune system, or which change and mutate constantly, such as HIV, Malaria, and Ebola, etc. [204]. In order to gather more potential immunomodulatory protein candidates, a novel process—reverse vaccinology—was developed 20 years ago. Whole virus and bacteria genomes are scanned, and immunological epitopes are predicted. Furthermore, conserved epitopes in core proteins are predicted to develop vaccines against many related strains of pathogens [205,206]. One example for reverse vaccinology is the development of a vaccine candidate against Ebola. With epitope prediction tools, proteins containing epitopes which present to human leukocyte antigen molecules and induce IFNγ in peripheral blood mononuclear cells were found [207]. In the last decade, a new discipline came into focus: systems vaccinology. It combines reverse vaccinology, which studies the role of immunogens, with additional factors like adjuvants and antigen delivery. With certain data-sets gained from systems-based studies, mainly by -omics techniques, it is possible to develop better vaccines based on improved insight into the immune response induced by the vaccine candidate, and to detect correlates of protection [204].
As for RHDV vaccine development, the fact that, in recent years, another highly-pathogenic virus variant emerged, which left the classic vaccines useless, demonstrated that it is important to constantly improve vaccines by, for example, considering the antigenicity of the core proteins of different virus strains to protect against possible new emerging strains, and to strive for not only B-cell mediated immunity but also adapted cellular and trained immunity, so that vaccines induce a full, long lasting protection.
Author Contributions
Conceptualization, P.N.-R., B.K., C.M.; methodology, R.H., D.B., M.B., J.M.; software, R.H., D.B.; validation, P.N.-R., B.K., C.M.; formal analysis, P.N.-R., B.K., C.M.; investigation, D.B., R.H., M.B., J.M., resources, P.N.-R., B.K.; data curation, C.M., R.H., D.B.; writing—original draft preparation, D.B., R.H., B.K., C.M., P.N.-R.; writing—review and editing, B.K., P.N.-R., J.M., M.B.; visualization, R.H., D.B., C.M.; supervision, P.N.-R., B.K.; project administration, P.N.-R.; funding acquisition, P.N.-R. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Di Giallonardo, F.; Holmes, EC. Viral biocontrol: Grand experiments in disease emergence and evolution. Trends Microbiol. 2015, 23, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Elsworth, P.G.; Kovaliski, J.; Cooke, B. Rabbit Haemorrhagic Disease: Are Australian rabbits (Oryctolagus cuniculus) evolving resistance to infection with Czech CAPM 351 RHDV? Epidemiol. Infect. 2012, 140, 1972–1981. [Google Scholar] [CrossRef]
- Elsworth, P.; Cooke, B.D.; Kovaliski, J.; Sinclair, R.; Holmes, E.C.; Strive, T. Increased virulence of rabbit haemorrhagic disease virus associated with genetic resistance in wild Australian rabbits (Oryctolagus cuniculus). Virology 2014, 464–465, 415–423. [Google Scholar] [CrossRef]
- Strive, T.; Elsworth, P.; Liu, J.; Wright, J.D.; Kovaliski, J.; Capucci, L. The non-pathogenic Australian rabbit calicivirus RCV-A1 provides temporal and partial cross protection to lethal Rabbit Haemorrhagic Disease Virus infection which is not dependent on antibody titres. Vet. Res. 2013, 44, 51. [Google Scholar] [CrossRef]
- Liu, S.J.; Xue, H.P.; Pu, B.Q.; Qian, N.H. A New Viral Disease in Rabbits. Anim. Hus. Vet. Med. 1984, 16, 253–255. [Google Scholar]
- Abrantes, J.; Van der Loo, W.; Le Pendu, J.; Esteves, P.J. Rabbit haemorrhagic disease (RHD) and rabbit haemorrhagic disease virus (RHDV): A review. Vet. Res. 2012, 43, 12. [Google Scholar] [CrossRef]
- Górski, J.; Mizak, B.; Mizak, Z.; Komorowski, A. Obraz kliniczny oraz zmiany anatomopatologiczne w przebiegu pomoru królików (wirusowej krwiotocznej bronchopneumonii królików). Życie Wet. 1988, 63, 266–269. [Google Scholar]
- Löliger, H.C.; Eskens, U. Incidence, epizootiology and control of viral haemorrhagic disease of rabbits and the European brown hare syndrome in Germany. Rev. Sci. Tech. 1991, 10, 423–434. [Google Scholar] [CrossRef] [PubMed]
- Morisse, J.P.; le Gall, G.; Boilletot, E. Hepatitis of Viral Origin in Leporidae: Introduction and Aetiological Hypotheses. Rev. Sci. Tech. 1991, 10, 269–310. [Google Scholar] [CrossRef]
- Le Gall, G.; Arnauld, C.; Boilletot, E.; Morisse, J.P.; Rasschaert, D. Molecular Epidemiology of Rabbit Haemorrhagic Disease Virus Outbreaks in France during 1988 to 1995. J. Gen. Virol. 1998, 79, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Cooke, B.D. Rabbit Haemorrhagic Disease: Field Epidemiology and the Management of Wild Rabbit Populations. OIE Rev. Sci. Tech. 2002, 21, 347–358. [Google Scholar] [CrossRef] [PubMed]
- Alda, F.; Gaitero, T.; Suárez, M.; Merchán, T.; Rocha, G.; Doadrio, I. Evolutionary History and Molecular Epidemiology of Rabbit Haemorrhagic Disease Virus in the Iberian Peninsula and Western Europe. BMC Evol. Biol. 2010, 10, 347. [Google Scholar] [CrossRef] [PubMed]
- Kinnear, M.; Linde, C.C. Capsid Gene Divergence in Rabbit Hemorrhagic Disease Virus. J. Gen. Virol. 2010, 91, 174–181. [Google Scholar] [CrossRef]
- Abrantes, J.; Lopes, A.M.; Dalton, K.P.; Melo, P.; Correia, J.J.; Ramada, M.; Alves, P.C.; Parra, F.; Esteves, P.J. New Variant of Rabbit Hemorrhagic Disease Virus, Portugal, 2012–2013. Emerg. Infect. Dis. 2013, 19, 1900–1902. [Google Scholar] [CrossRef] [PubMed]
- Abrantes, J.; Lopes, A.M.; Dalton, K.P.; Parra, F.; Esteves, P.J. Detection of RHDVa on the Iberian Peninsula: Isolation of an RHDVa Strain from a Spanish Rabbitry. Arch. Virol. 2014, 159, 321–326. [Google Scholar] [CrossRef]
- Westcott, D.G.; Frossard, J.P.; Everest, D.; Dastjerdi, A.; Duff, J.P.; Steinbach, F.; Choudhury, B. Incursion of RHDV2-like Variant in Great Britain. Vet. Rec. 2014, 174, 333. [Google Scholar] [CrossRef]
- Bárcena, J.; Guerra, B.; Angulo, I.; González, J.; Valcárcel, F.; Mata, C.P.; Castón, J.R.; Blanco, E.; Alejo, A. Comparative Analysis of Rabbit Hemorrhagic Disease Virus (RHDV) and New RHDV2 Virus Antigenicity, Using Specific Virus-like Particles. Vet. Res. 2015, 46. [Google Scholar] [CrossRef] [PubMed]
- Monterroso, P.; Garrote, G.; Serronha, A.; Santos, E.; Delibes-Mateos, M.; Abrantes, J.; de Ayala, R.P.; Silvestre, F.; Carvalho, J.; Vasco, I.; et al. Disease-Mediated Bottom-up Regulation: An Emergent Virus Affects a Keystone Prey, and Alters the Dynamics of Trophic Webs. Sci. Rep. 2016, 6, 1–9. [Google Scholar] [CrossRef]
- Rouco, C.; Abrantes, J.; Serronha, A.; Lopes, A.M.; Maio, E.; Magalhães, M.J.; Blanco, E.; Bárcena, J.; Esteves, P.J.; Santos, N.; et al. Epidemiology of RHDV2 (Lagovirus Europaeus/GI.2) in Free-Living Wild European Rabbits in Portugal. Transbound. Emerg. Dis. 2018, 65, e373–e382. [Google Scholar] [CrossRef] [PubMed]
- Gibb, J.A.; Williams, J.M. The rabbit in New Zealand. In The European Rabbit: History and Biology of a Successful Colonizer; Thompson, H.V., King, C.M., Eds.; Oxford University Press: Oxford, UK, 1994; pp. 158–200. [Google Scholar]
- Mutze, G.; Cooke, B.; Alexander, P. The Initial Impact of Rabbit Hemorrhagic Disease on European Rabbit Populations in South Australia. J. Wildl. Dis. 1998, 34, 221–227. [Google Scholar] [CrossRef]
- Fenner, F. Deliberate Introduction of the European Rabbit, Oryctolagus Cuniculus, into Australia. OIE Rev. Sci. Tech. 2010, 29, 103–111. [Google Scholar] [CrossRef]
- Granzow, H.; Schirrmeier, H.; Tews, G. Hämorrhagische Septikämie Der Kaninchen—Erregernachweis Und Erste Elektronenmikroskopische Charakterisierung. Mh. Vet. Med. 1989, 44, 379–380. [Google Scholar]
- Ohlinger, V.F.; Haas, B.; Ahl, R.; Weiland, F. Rabbit haemorrhagic disease—A contagious disease caused by Calicivirus. Tierärztliche Umschau. 1989, 44, 284–294. [Google Scholar]
- Ohlinger, V.F.; Haas, B.; Meyers, G.; Weiland, F.; Thiel, H.J. Identification and Characterization of the Virus Causing Rabbit Hemorrhagic Disease. J. Virol. 1990, 64, 3331–3336. [Google Scholar] [CrossRef] [PubMed]
- Parra, F.; Prieto, M. Purification and Characterization of a Calicivirus as the Causative Agent of a Lethal Hemorrhagic Disease in Rabbits. J. Virol. 1990, 64, 4013–4015. [Google Scholar] [CrossRef]
- Meyers, G.; Wirblich, C.; Thiel, H.J. Rabbit Hemorrhagic Disease Virus-Molecular Cloning and Nucleotide Sequencing of a Calicivirus Genome. Virology 1991, 184, 664–676. [Google Scholar] [CrossRef]
- Moussa, A.; Chasey, D.; Lavazza, A.; Capucci, L.; Šmid, B.; Meyers, G.; Rossi, C.; Thiel, H.J.; Vlásak, R.; Rønsholt, L.; et al. Haemorrhagic Disease of Lagomorphs: Evidence for a Calicivirus. Vet. Microbiol. 1992, 33, 375–381. [Google Scholar] [CrossRef]
- Le Gall-Reculé, G.; Lavazza, A.; Marchandeau, S.; Bertagnoli, S.; Zwingelstein, F.; Cavadini, P.; Martinelli, N.; Lombardi, G.; Guérin, J.L.; Lemaitre, E.; et al. Emergence of a New Lagovirus Related to Rabbit Haemorrhagic Disease Virus. Vet. Res. 2013, 44, 81. [Google Scholar] [CrossRef]
- Puggioni, G.; Cavadini, P.; Maestrale, C.; Scivoli, R.; Botti, G.; Ligios, C.; le Gall-Reculé, G.; Lavazza, A.; Capucci, L. The New French 2010 Rabbit Hemorrhagic Disease Virus Causes an RHD-like Disease in the Sardinian Cape Hare (Lepus Capensis Mediterraneus). Vet. Res. 2013, 44, 96. [Google Scholar] [CrossRef] [PubMed]
- Camarda, A.; Pugliese, N.; Cavadini, P.; Circella, E.; Capucci, L.; Caroli, A.; Legretto, M.; Mallia, E.; Lavazza, A. Detection of the New Emerging Rabbit Haemorrhagic Disease Type 2 Virus (RHDV2) in Sicily from Rabbit (Oryctolagus Cuniculus) and Italian Hare (Lepus Corsicanus). Res. Vet. Sci. 2014, 97, 642–645. [Google Scholar] [CrossRef] [PubMed]
- Rouco, C.; Aguayo-Adán, J.A.; Santoro, S.; Abrantes, J.; Delibes-Mateos, M. Worldwide Rapid Spread of the Novel Rabbit Haemorrhagic Disease Virus (GI.2/RHDV2/b). Transbound. Emerg. Dis. 2019, 66, 1762–1764. [Google Scholar] [CrossRef]
- Le Gall-Reculé, G.; Lemaitre, E.; Bertagnoli, S.; Hubert, C.; Top, S.; Decors, A.; Marchandeau, S.; Guitton, J.S. Large-Scale Lagovirus Disease Outbreaks in European Brown Hares (Lepus Europaeus) in France Caused by RHDV2 Strains Spatially Shared with Rabbits (Oryctolagus Cuniculus). Vet. Res. 2017, 48, 70. [Google Scholar] [CrossRef]
- Neimanis, A.S.; Ahola, H.; Larsson Pettersson, U.; Lopes, A.M.; Abrantes, J.; Zohari, S.; Esteves, P.J.; Gavier-Widén, D. Overcoming Species Barriers: An Outbreak of Lagovirus Europaeus GI.2/RHDV2 in an Isolated Population of Mountain Hares (Lepus Timidus). BMC Vet. Res. 2018, 14, 1–12. [Google Scholar] [CrossRef]
- Lopes, A.M.; Marques, S.; Silva, E.; Magalhães, M.J.; Pinheiro, A.; Alves, P.C.; le Pendu, J.; Esteves, P.J.; Thompson, G.; Abrantes, J. Detection of RHDV Strains in the Iberian Hare (Lepus Granatensis): Earliest Evidence of Rabbit Lagovirus Cross-Species Infection. Vet. Res. 2014, 45, 94. [Google Scholar] [CrossRef] [PubMed][Green Version]
- House Rabbit Society. Rabbit Hemorrhagic Disease Virus (RHDV). Available online: https://rabbit.org/rhdv/#southwest (accessed on 22 February 2021).
- USGS Science for a Changing World. An Update on NWHC Support to Wildlife Managers and Response to RHDV2. Available online: https://www.usgs.gov/media/files/update-nwhc-support-wildlife-managers-and-response-rhdv2 (accessed on 22 February 2021).
- Trzeciak-Ryczek, A.; Tokarz-Deptuła, B.; Deptuła, W. The Importance of Liver Lesions and Changes to Biochemical and Coagulation Factors in the Pathogenesis of RHD. Acta Biochim. Pol. 2015, 62, 169–171. [Google Scholar] [CrossRef]
- Nowotny, N.; Fuchs, A.; Schilcher, F.; Loupal, G. Zum Auftreten der Rabbit Haemorrhagic Disease (RHD) in Österreich: I Pathomorphologische und virologische Untersuchungen. Wien. Tierärztl. Mschr. 1990, 77, 19–23. [Google Scholar]
- Chasey, D.; Lucas, M.; Westcott, D.; Williams, M. European Brown Hare Syndrome in the U.K.; a Calicivirus Related to but Distinct from That of Viral Haemorrhagic Disease in Rabbits. Arch. Virol. 1992, 124, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Gregg, D.A.; House, C.; Meyer, R.; Berninger, M. Viral Haemorrhagic Disease of Rabbits in Mexico: Epidemiology and Viral Characterization. Rev. Sci. Tech. 1991, 10, 435–451. [Google Scholar] [CrossRef]
- Gall, A.; Hoffmann, B.; Teifke, J.P.; Lange, B.; Schirrmeier, H. Persistence of Viral RNA in Rabbits Which Overcome an Experimental RHDV Infection Detected by a Highly Sensitive Multiplex Real-Time RT-PCR. Vet. Microbiol. 2007, 120, 17–32. [Google Scholar] [CrossRef]
- Alonso, C.; Oviedo, J.M.; Martín-Alonso, J.M.; Díaz, E.; Boga, J.A.; Parra, F. Programmed Cell Death in the Pathogenesis of Rabbit Hemorrhagic Disease. Arch. Virol. 1998, 143, 321–332. [Google Scholar] [CrossRef] [PubMed]
- Teifke, J.P.; Reimann, I.; Schirrmeier, H. Subacute Liver Necrosis after Experimental Infection with Rabbit Haemorrhagic Disease Virus (RHDV). J. Comp. Pathol. 2002, 126, 231–234. [Google Scholar] [CrossRef] [PubMed]
- Ueda, K.; Park, J.H.; Ochiai, K.; Itakura, C. Disseminated intravascular coagulation (DIC) in rabbit haemorrhagic disease. Jpn. J. Vet. Res. 1992, 40, 133–141. [Google Scholar]
- Sanchez-Campos, S.; Alvarez, M.; Culebras, J.M.; Gonzalez-Gallego, J.; Tunon, M.J. Pathogenic Molecular Mechanisms in an Animal Model of Fulminant Hepatic Failure: Rabbit Hemorrhagic Viral Disease. J. Lab. Clin. Med. 2004, 144, 215–222. [Google Scholar] [CrossRef]
- Niedźwiedzka-Rystwej, P.; Deptuła, W. Apoptosis of Peripheral Blood Leukocytes from Rabbits Infected with Non-Haemagglutinating Strains of Rabbit Haemorrhagic Disease Virus (RHDV). Vet. Immunol. Immunopathol. 2012, 149, 54–57. [Google Scholar] [CrossRef] [PubMed]
- Niedźwiedzka-Rystwej, P.; Hukowska-Szematowicz, B.; Tokarz-Deptuła, B.; Trzeciak-Ryczek, A.; Działo, J.; Deptuła, W. Apoptosis of peripheral blood leucocytes in rabbits infected with different strains of rabbit haemorrhagic disease virus. Acta Biochim. Pol. 2013, 60, 65–69. [Google Scholar] [CrossRef]
- Marques, R.M.; Costa-E-Silva, A.; Águas, A.P.; Teixeira, L.; Ferreira, P.G. Early Acute Depletion of Lymphocytes in Calicivirus-Infected Adult Rabbits. Vet. Res. Commun. 2010, 34, 659–668. [Google Scholar] [CrossRef]
- Teixeira, L.; Marques, R.M.; Águas, A.P.; Ferreira, P.G. Regulatory T Cells Are Decreased in Acute RHDV Lethal Infection of Adult Rabbits. Vet. Immunol. Immunopathol. 2012, 148, 343–347. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, P.G.; Costa-e-Silva, A.; Oliveira, M.J.R.; Monteiro, E.; Cunha, E.M.; Águas, A.P. Severe Leukopenia and Liver Biochemistry Changes in Adult Rabbits after Calicivirus Infection. Res. Vet. Sci. 2006, 80, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Marques, R.M.; Costa-E-Silva, A.; Águas, A.P.; Teixeira, L.; Ferreira, P.G. Early Inflammatory Response of Young Rabbits Attending Natural Resistance to Calicivirus (RHDV) Infection. Vet. Immunol. Immunopathol. 2012, 150, 181–188. [Google Scholar] [CrossRef]
- Trzeciak-Ryczek, A.; Tokarz-Deptuła, B.; Deptuła, W. Expression of IL-1Ra, IL-6, IL-8, IL-18, TNF-α and IFN-γ Genes in Peripheral Blood Leukocytes of Rabbits Infected with RHDV (Rabbit Haemorrhagic Disease Virus). Dev. Comp. Immunol. 2017, 76, 310–315. [Google Scholar] [CrossRef] [PubMed]
- Argüello Villares, J.L. Viral Haemorrhagic Disease of Rabbits: Vaccination and Immune Response. Rev. Sci. Tech. 1991, 10, 459–480. [Google Scholar] [CrossRef]
- Huang, H.B. Vaccination against and Immune Response to Viral Haemorrhagic Disease of Rabbits: A Review of Research in the People’s Republic of China. Rev. Sci. Tech. Off. Int. Epiz. 1991, 10, 481–498. [Google Scholar]
- Šmíd, B.; Valíček, L.; Rodák, L.; Štěpánek, J.; Jurák, E. Rabbit Haemorrhagic Disease: An Investigation of Some Properties of the Virus and Evaluation of an Inactivated Vaccine. Vet. Microbiol. 1991, 26, 77–85. [Google Scholar] [CrossRef]
- Vautherot, J.F.; Laurent, S.; le Gall, G.; Morisse, J.P.; Rasschaert, D. Recombinant vaccine against rabbit hemorrhagic disease virus. Vet. Res. 1995, 26, 206–207. [Google Scholar]
- Soledad Marín, M.; Martín Alonso, J.M.; Pérez Ordoyo García, L.I.; Antonio Boga, J.; Argüello-Villares, J.L.; Casais, R.; Venugopal, K.; Jiang, W.; Gould, E.A.; Parra, F. Immunogenic Properties of Rabbit Haemorrhagic Disease Virus Structural Protein VP60 Expressed by a Recombinant Baculovirus: An Efficient Vaccine. Virus Res. 1995, 39, 119–128. [Google Scholar] [CrossRef]
- Plana-Duran, J.; Bastons, M.; Rodriguez, M.J.; Climent, I.; Cortés, E.; Vela, C.; Casal, I. Oral Immunization of Rabbits with VP60 Particles Confers Protection against Rabbit Hemorrhagic Disease. Arch. Virol. 1996, 141, 1423–1436. [Google Scholar] [CrossRef] [PubMed]
- Bertagnoli, S.; Gelfi, J.; le Gall, G.; Boilletot, E.; Vautherot, J.F.; Rasschaert, D.; Laurent, S.; Petit, F.; Boucraut-Baralon, C.; Milon, A. Protection against Myxomatosis and Rabbit Viral Hemorrhagic Disease with Recombinant Myxoma Viruses Expressing Rabbit Hemorrhagic Disease Virus Capsid Protein. J. Virol. 1996, 70, 5061–5066. [Google Scholar] [CrossRef] [PubMed]
- Bertagnoli, S.; Gelfi, J.; Petit, F.; Vautherot, J.F.; Rasschaert, D.; Laurent, S.; le Gall, G.; Boilletot, E.; Chantal, J.; Boucraut-Baralon, C. Protection of Rabbits against Rabbit Viral Haemorrhagic Disease with a Vaccinia-RHDV Recombinant Virus. Vaccine 1996, 14, 506–510. [Google Scholar] [CrossRef]
- Fischer, L.; le Gros, F.X.; Mason, P.W.; Paoletti, E. A Recombinant Canarypox Virus Protects Rabbits against a Lethal Rabbit Hemorrhagic Disease Virus (RHDV) Challenge. Vaccine 1997, 15, 90–96. [Google Scholar] [CrossRef]
- Castañón, S.; Marín, M.S.; Martín-Alonso, J.M.; Boga, J.A.; Casais, R.; Humara, J.M.; Ordás, R.J.; Parra, F. Immunization with Potato Plants Expressing VP60 Protein Protects against Rabbit Hemorrhagic Disease Virus. J. Virol. 1999, 73, 4452–4455. [Google Scholar] [CrossRef]
- Abrantes, J.; Droillard, C.; Lopes, A.M.; Lemaitre, E.; Lucas, P.; Blanchard, Y.; Marchandeau, S.; Esteves, P.J.; Le Gall-Reculé, G. Recombination at the Emergence of the Pathogenic Rabbit Haemorrhagic Disease Virus Lagovirus Europaeus/GI.2. Sci. Rep. 2020, 10, 14502. [Google Scholar] [CrossRef]
- Abrantes, J.; Esteves, P.J.; van der Loo, W. Evidence for Recombination in the Major Capsid Gene VP60 of the Rabbit Haemorrhagic Disease Virus (RHDV). Arch. Virol. 2008, 153, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Forrester, N.L.; Trout, R.C.; Gould, E.A. Benign Circulation of Rabbit Haemorrhagic Disease Virus on Lambay Island, Eire. Virology 2007, 358, 18–22. [Google Scholar] [CrossRef]
- Hu, B.; Wang, F.; Fan, Z.; Song, Y.; Abrantes, J.; Zuo, Y.; Esteves, P.J. Recombination between G2 and G6 Strains of Rabbit Hemorrhagic Disease Virus (RHDV) in China. Arch. Virol. 2017, 162, 269–272. [Google Scholar] [CrossRef]
- Lopes, A.M.; Dalton, K.P.; Magalhães, M.J.; Parra, F.; Esteves, P.J.; Holmes, E.C.; Abrantes, J. Full Genomic Analysis of New Variant Rabbit Hemorrhagic Disease Virus Revealed Multiple Recombination Events. J. Gen. Virol. 2015, 96, 1309–1319. [Google Scholar] [CrossRef]
- White, P.J.; Trout, R.C.; Moss, S.R.; Desai, A.; Armesto, M.; Forrester, N.L.; Gould, E.A.; Hudson, P.J. Epidemiology of Rabbit Haemorrhagic Disease Virus in the United Kingdom: Evidence for Seasonal Transmission by Both Virulent and Avirulent Modes of Infection. Epidemiol. Infect. 2004, 132, 555–567. [Google Scholar] [CrossRef]
- Capucci, L.; Fusi, P.; Lavazza, A.; Pacciarini, M.L.; Rossi, C. Detection and Preliminary Characterization of a New Rabbit Calicivirus Related to Rabbit Hemorrhagic Disease Virus but Nonpathogenic. J. Virol. 1996, 70, 8614–8623. [Google Scholar] [CrossRef] [PubMed]
- Moss, S.R.; Turner, S.L.; Trout, R.C.; White, P.J.; Hudson, P.J.; Desai, A.; Armesto, M.; Forrester, N.L.; Gould, E.A. Molecular Epidemiology of Rabbit Haemorrhagic Disease Virus. J. Gen. Virol. 2002, 83, 2461–2467. [Google Scholar] [CrossRef]
- Forrester, N.L.; Abubakr, M.I.; Abu Elzein, E.M.E.; Al-Afaleq, A.I.; Housawi, F.M.T.; Moss, S.R.; Turner, S.L.; Gould, E.A. Phylogenetic Analysis of Rabbit Haemorrhagic Disease Virus Strains from the Arabian Peninsula: Did RHDV Emerge Simultaneously in Europe and Asia? Virology 2006, 344, 277–282. [Google Scholar] [CrossRef]
- Strive, T.; Wright, J.; Kovaliski, J.; Botti, G.; Capucci, L. The Non-Pathogenic Australian Lagovirus RCV-A1 Causes a Prolonged Infection and Elicits Partial Cross-Protection to Rabbit Haemorrhagic Disease Virus. Virology 2010, 398, 125–134. [Google Scholar] [CrossRef]
- Kerr, P.J.; Kitchen, A.; Holmes, E.C. Origin and Phylodynamics of Rabbit Hemorrhagic Disease Virus. J. Virol. 2009, 83, 12129–12138. [Google Scholar] [CrossRef] [PubMed]
- Esteves, P.J.; Abrantes, J.; Bertagnoli, S.; Cavadini, P.; Gavier-Widén, D.; Guitton, J.-S.; Lavazza, A.; Lemaitre, E.; Letty, J.; Lopes, A.M.; et al. Emergence of Pathogenicity in Lagoviruses: Evolution from Pre-Existing Nonpathogenic Strains or through a Species Jump? PLoS Pathog. 2015, 11, e1005087. [Google Scholar] [CrossRef]
- Merchán, T.; Rocha, G.; Alda, F.; Silva, E.; Thompson, G.; de Trucios, S.H.; Pagés, A. Detection of Rabbit Haemorrhagic Disease Virus (RHDV) in Nonspecific Vertebrate Hosts Sympatric to the European Wild Rabbit (Oryctolagus Cuniculus). Infect. Genet. Evol. 2011, 11, 1469–1474. [Google Scholar] [CrossRef]
- Gavier-Widén, D.; Mörner, T. Epidemiology and Diagnosis of the European Brown Hare Syndrome in Scandinavian Countries: A Review. Rev. Sci. Tech. 1991, 10, 453–458. [Google Scholar] [CrossRef] [PubMed]
- Wirblich, C.; Meyers, G.; Ohlinger, V.F.; Capucci, L.; Eskens, U.; Haas, B.; Thiel, H.J. European Brown Hare Syndrome Virus: Relationship to Rabbit Hemorrhagic Disease Virus and Other Caliciviruses. J. Virol. 1994, 68, 5164–5173. [Google Scholar] [CrossRef]
- Nowotny, N.; Ros Bascuñana, C.; Ballagi-Pordány, A.; Gavier-Widén, D.; Uhlén, M.; Belák, S. Phylogenetic Analysis of Rabbit Haemorrhagic Disease and European Brown Hare Syndrome Viruses by Comparison of Sequences from the Capsid Protein Gene. Arch. Virol. 1997, 142, 657–673. [Google Scholar] [CrossRef]
- Lavazza, A.; Scicluna, M.T.; Capucci, L. Susceptibility of Hares and Rabbits to the European Brown Hare Syndrome Virus (EBHSV) and Rabbit Haemorrhagic Disease Virus (RHDV) under Experimental Conditions. Zentralbl. Veterinarmed. B 1996, 43, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Cancellotti, F.M.; Renzi, M. Epidemiology and Current Situation of Viral Haemorrhagic Disease of Rabbits and the European Brown Hare Syndrome in Italy. Rev. Sci. Tech. 1991, 10, 409–422. [Google Scholar] [CrossRef] [PubMed]
- Park, N.Y.; Chong, C.Y.; Kim, J.H.; Cho, S.M.; Cha, Y.H.; Jung, B.T.; Kim, D.S.; Yoon, J.B. An Outbreak of Viral Haemorrhagic Pneumonia (Tentative Name) of Rabbits in Korea. J. Korean Vet. Med. Assoc. 1987, 23, 603–610. [Google Scholar]
- Xu, W.Y. Viral Haemorrhagic Disease of Rabbits in the People’s Republic of China: Epidemiology and Virus Characterisation. Rev. Sci. Tech. 1991, 10, 393–408. [Google Scholar] [PubMed]
- Argüello Villares, J.L.; Llanos Pellitero, A.; Pérez-Ordoyo Garcia, L.I. Enfermedad vírica hemorragica del canejo. Rev. Med. Vet. 1988, 5, 645–650. [Google Scholar]
- Villafuerte, R.; Calvete, C.; Blanco, J.C.; Lucientes, J. Incidence of Viral Hemorrhagic Disease in Wild Rabbit Populations in Spain. Mammalia 1995, 59, 651–660. [Google Scholar] [CrossRef]
- Delibes-Mateos, M.; Redpath, S.M.; Angulo, E.; Ferreras, P.; Villafuerte, R. Rabbits as a Keystone Species in Southern Europe. Biol. Conserv. 2007, 137, 149–156. [Google Scholar] [CrossRef]
- Le Gall-Reculé, G.; Zwingelstein, F.; Laurent, S.; de Boisséson, C.; Portejoie, Y.; Rasschaert, D. Phylogenetic Analysis of Rabbit Haemorrhagic Disease Virus in France between 1993 and 2000, and the Characterisation of RHDV Antigenic Variants. Arch. Virol. 2003, 148, 65–81. [Google Scholar] [CrossRef] [PubMed]
- Farnós, O.; Rodríguez, D.; Valdés, O.; Chiong, M.; Parra, F.; Toledo, J.R.; Fernández, E.; Lleonart, R.; Suárez, M. Molecular and Antigenic Characterization of Rabbit Hemorrhagic Disease Virus Isolated in Cuba Indicates a Distinct Antigenic Subtype. Arch. Virol. 2007, 152, 1215–1221. [Google Scholar] [CrossRef]
- Cooke, B.D.; Fenner, F. Rabbit Haemorrhagic Disease and the Biological Control of Wild Rabbits, Oryctolagus Cuniculus, in Australia and New Zealand. Wildl. Res. 2002, 29, 689–706. [Google Scholar] [CrossRef]
- Thompson, J.; Clark, G. Rabbit Calicivirus Disease Now Established in New Zealand. Surveillance 1997, 24, 5–6. [Google Scholar]
- O’Keefe, J.S.; Tempero, J.; Atkinson, P.H.; Pacciarini, L.; Fallacara, F.; Horner, G.W.; Motha, J. Typing of Rabbit Haemorrhagic Disease Virus from New Zealand Wild Rabbits. N. Z. Vet. J. 1998, 46, 42–43. [Google Scholar] [CrossRef]
- Vinjé, J.; Estes, M.K.; Esteves, P.; Green, K.Y.; Katayama, K.; Knowles, N.J.; L’Homme, Y.; Martella, V.; Vennema, H.; White, P.A.; et al. ICTV Virus Taxonomy Profile: Caliciviridae. J. Gen. Virol. 2019, 100, 1469–1470. [Google Scholar] [CrossRef] [PubMed]
- Le Pendu, J.; Abrantes, J.; Bertagnoli, S.; Guitton, J.S.; le Gall-Reculé, G.; Lopes, A.M.; Marchandeau, S.; Alda, F.; Almeida, T.; Célio, A.P.; et al. Proposal for a Unified Classification System and Nomenclature of Lagoviruses. J. Gen. Virol. 2017, 98, 1658–1666. [Google Scholar] [CrossRef]
- Velarde, R.; Cavadini, P.; Neimanis, A.; Cabezón, O.; Chiari, M.; Gaffuri, A.; Lavín, S.; Grilli, G.; Gavier-Widén, D.; Lavazza, A.; et al. Spillover Events of Infection of Brown Hares (Lepus Europaeus) with Rabbit Haemorrhagic Disease Type 2 Virus (RHDV2) Caused Sporadic Cases of an European Brown Hare Syndrome-Like Disease in Italy and Spain. Transbound. Emerg. Dis. 2017, 64, 1750–1761. [Google Scholar] [CrossRef]
- Hall, R.N.; Peacock, D.E.; Kovaliski, J.; Mahar, J.E.; Mourant, R.; Piper, M.; Strive, T. Detection of RHDV2 in European Brown Hares (Lepus Europaeus) in Australia. Vet. Rec. 2017, 180, 121. [Google Scholar] [CrossRef]
- Capucci, L.; Scicluna, M.T.; Lavazza, A. Diagnosis of Viral Haemorrhagic Disease of Rabbits and the European Brown Hare Syndrome. Rev. Sci. Tech. 1991, 10, 347–370. [Google Scholar] [CrossRef]
- Marcato, P.S.; Benazzi, C.; Vecchi, G.; Galeotti, M.; della Salda, L.; Sarli, G.; Lucidi, P. Clinical and Pathological Features of Viral Haemorrhagic Disease of Rabbits and the European Brown Hare Syndrome. Rev. Sci. Tech. 1991, 10, 371–392. [Google Scholar] [CrossRef]
- Fuchs, A.; Weissenböck, H. Comparative histopathological study of rabbit haemorrhagic disease (RHD) and European brown hare syndrome (EBHS). Comp. Study J. Comp. Pathol. 1992, 107, 103–113. [Google Scholar] [CrossRef]
- Niedźwiedzka-Rystwej, P.; Deptuła, W. Cytometric analysis of lymphocytes T and B in rabbits infected with non-haemagglutinogenic strains of RHD virus (rabbit haemorrhagic disease)—Rainham, Frankfurt and Asturias. Pol. J. Vet. Sci. 2010, 13, 157–162. [Google Scholar]
- Bębnowska, D.; Niedźwiedzka-Rystwej, P. Characteristics of a New Variant of Rabbit Haemorrhagic Disease Virus—RHDV2. Acta Biolog. 2019, 26, 83–97. [Google Scholar] [CrossRef]
- Niedźwiedzka-Rystwej, P.; Tokarz-Deptuła, B.; Hukowska-Szematowicz, B.; Działo, J.; Deptuła, W. Indices of Non-Specific Immunity: An Element of Natural Immunity in Rabbits Infected with RHD (Rabbit Haemorrhagic Disease) Virus. Cent. Eur. J. Immunol. 2013, 38, 231–236. [Google Scholar] [CrossRef]
- Niedźwiedzka-Rystwej, P.; Tokarz-Deptuła, B.; Deptuła, W. Lymphocytes T and B in rabbits infected with RHD virus. Pol. J. Vet. Sci. 2013, 16, 563–565. [Google Scholar] [CrossRef] [PubMed]
- Lopes, A.M.; Capucci, L.; Gavier-Widén, D.; le Gall-Reculé, G.; Brocchi, E.; Barbieri, I.; Quéméner, A.; le Pendu, J.; Geoghegan, J.L.; Holmes, E.C.; et al. Molecular Evolution and Antigenic Variation of European Brown Hare Syndrome Virus (EBHSV). Virology 2014, 468–470, 104–112. [Google Scholar] [CrossRef]
- International Committee on Taxonomy of Viruses (ICTV). Genus: Lagovirus. Available online: https://talk.ictvonline.org/ictv-reports/ictv_online_report/positive-sense-rna-viruses/w/caliciviridae/1163/genus-lagovirus (accessed on 10 January 2021).
- Capucci, L.; Frigoli, G.; Rønshold, L.; Lavazza, A.; Brocchi, E.; Rossi, C. Antigenicity of the Rabbit Hemorrhagic Disease Virus Studied by Its Reactivity with Monoclonal Antibodies. Virus Res. 1995, 37, 221–238. [Google Scholar] [CrossRef]
- Capucci, L.; Fallacara, F.; Grazioli, S.; Lavazza, A.; Pacciarini, M.L.; Brocchi, E. A Further Step in the Evolution of Rabbit Hemorrhagic Disease Virus: The Appearance of the First Consistent Antigenic Variant. Virus Res. 1998, 58, 115–126. [Google Scholar] [CrossRef]
- Schirrmeier, H.; Reimann, I.; Köllner, B.; Granzow, H. Pathogenic, Antigenic and Molecular Properties of Rabbit Haemorrhagic Disease Virus (RHDV) Isolated from Vaccinated Rabbits: Detection and Characterization of Antigenic Variants. Arch. Virol. 1999, 144, 719–735. [Google Scholar] [CrossRef] [PubMed]
- Dalton, K.P.; Nicieza, I.; Abrantes, J.; Esteves, P.J.; Parra, F. Spread of New Variant RHDV in Domestic Rabbits on the Iberian Peninsula. Vet. Microbiol. 2014, 169, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Calvete, C.; Mendoza, M.; Alcaraz, A.; Sarto, M.P.; Jiménez-de-Bagüéss, M.P.; Calvo, A.J.; Monroy, F.; Calvo, J.H. Rabbit Haemorrhagic Disease: Cross-Protection and Comparative Pathogenicity of GI.2/RHDV2/b and GI.1b/RHDV Lagoviruses in a Challenge Trial. Vet. Microbiol. 2018, 219, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Rahali, N.; Sghaier, S.; Kbaier, H.; Zanati, A.; Bahloul, C. Genetic Characterization and Phylogenetic Analysis of Rabbit Hemorrhagic Disease Virus Isolated in Tunisia from 2015 to 2018. Arch. Virol. 2019, 164, 2327–2332. [Google Scholar] [CrossRef]
- Müller, C.; Ulrich, R.; Schinköthe, J.; Müller, M.; Köllner, B. Characterization of Protective Humoral and Cellular Immune Responses against RHDV2 Induced by a New Vaccine Based on Recombinant Baculovirus. Vaccine 2019, 37, 4195–4203. [Google Scholar] [CrossRef] [PubMed]
- Gehrmann, B.; Kretzschmar, C. Ein experimenteller Beitrag zur Epizootiologie der Viralen Hämorrhagischen Septikämie der Kaninchen (Rabbit Haemorrhagic Disease, RHD) Übertragung durch Fliegen. Berl. Munch. Tierarztl. Wschr. 1991, 104, 192–194. [Google Scholar]
- Xu, Z.J.; Chen, W.X. Viral Haemorrhagic Disease in Rabbits: A Review. Vet. Res. Commun. 1989, 13, 205–212. [Google Scholar] [CrossRef]
- Ohlinger, V.F.; Haas, B.; Thiel, H.J. Rabbit hemorrhagic disease (RHD): Characterization of the causative calicivirus. Vet. Res. 1993, 24, 103–116. [Google Scholar] [PubMed]
- Asgari, S.; Hardy, J.R.E.; Sinclair, R.G.; Cooke, B.D. Field Evidence for Mechanical Transmission of Rabbit Haemorrhagic Disease Virus (RHDV) by Flies (Diptera: Calliphoridae) among Wild Rabbits in Australia. Virus Res. 1998, 54, 123–132. [Google Scholar] [CrossRef]
- Dalton, K.P.; Nicieza, I.; Balseiro, A.; Muguerza, M.A.; Rosell, J.M.; Casais, R.; Álvarez, Á.L.; Parra, F. Variant Rabbit Hemorrhagic Disease Virus in Young Rabbits, Spain. Emerg. Infect. Dis. 2012, 18, 2009–2012. [Google Scholar] [CrossRef]
- Knight, K.L.; Winstead, C.R. Generation of Antibody Diversity in Rabbits. Curr. Opin. Immunol. 1997, 9, 228–232. [Google Scholar] [CrossRef]
- Crisci, E.; Almanza, H.; Mena, I.; Córdoba, L.; Gómez-Casado, E.; Castón, J.R.; Fraile, L.; Bárcena, J.; Montoya, M. Chimeric Calicivirus-like Particles Elicit Protective Anti-Viral Cytotoxic Responses without Adjuvant. Virology 2009, 387, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Li, J.N. Studies on interferon anti RHDV activities in rabbits. J. Anim. Quar. 1990, 3, 19–22. [Google Scholar]
- Cooke, B.D.; Robinson, A.J.; Merchant, J.C.; Nardin, A.; Capucci, L. Use of ELISAs in Field Studies of Rabbit Haemorrhagic Disease (RHD) in Australia. Epidemiol. Infect. 2000, 124, 563–576. [Google Scholar] [CrossRef]
- Ferreira, P.G.; Dinís, M.; Costa-e-Silva, A.; Águas, A.P. Adult Rabbits Acquire Resistance to Lethal Calicivirus Infection by Adoptive Transfer of Sera from Infected Young Rabbits. Vet. Immunol. Immunopathol. 2008, 121, 364–369. [Google Scholar] [CrossRef]
- Ruvoën-Clouet, N.; Ganière, J.P.; André-Fontaine, G.; Blanchard, D.; le Pendu, J. Binding of Rabbit Hemorrhagic Disease Virus to Antigens of the ABH Histo-Blood Group Family. J. Virol. 2000, 74, 11950–11954. [Google Scholar] [CrossRef]
- Mikami, O.; Park, J.H.; Kimura, T.; Ochiai, K.; Itakura, C. Hepatic Lesions in Young Rabbits Experimentally Infected with Rabbit Haemorrhagic Disease Virus. Res. Vet. Sci. 1999, 66, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, P.G.; Costa-e-Silva, A.; Monteiro, E.; Oliveira, M.J.R.; Águas, A.P. Transient Decrease in Blood Heterophils and Sustained Liver Damage Caused by Calicivirus Infection of Young Rabbits That Are Naturally Resistant to Rabbit Haemorrhagic Disease. Res. Vet. Sci. 2004, 76, 83–94. [Google Scholar] [CrossRef]
- Marques, R.M.; Teixeira, L.; Águas, A.P.; Ribeiro, J.C.; Costa-E-Silva, A.; Ferreira, P.G. Immunosuppression Abrogates Resistance of Young Rabbits to Rabbit Haemorrhagic Disease (RHD). Vet. Res. 2014, 45, 14. [Google Scholar] [CrossRef]
- Vallejo, D.; Crespo, I.; San-Miguel, B.; Álvarez, M.; Prieto, J.; Tuñón, M.J.; González-Gallego, J. Autophagic Response in the Rabbit Hemorrhagic Disease, an Animal Model of Virally-Induced Fulminant Hepatic Failure. Vet. Res. 2014, 45. [Google Scholar] [CrossRef]
- Neave, M.J.; Hall, R.N.; Huang, N.; McColl, K.A.; Kerr, P.; Hoehn, M.; Taylor, J.; Strive, T. Robust Innate Immunity of Young Rabbits Mediates Resistance to Rabbit Hemorrhagic Disease Caused by Lagovirus Europaeus GI.1 but Not GI.2. Viruses 2018, 10, 512. [Google Scholar] [CrossRef]
- Müller, C.; Ulrich, R.; Franzke, K.; Müller, M.; Köllner, B. Crude Extracts of Recombinant Baculovirus Expressing Rabbit Hemorrhagic Disease Virus 2 VLPs from Both Insect and Rabbit Cells Protect Rabbits from Rabbit Hemorrhagic Disease Caused by RHDV2. Arch. Virol. 2019, 164, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Prieto, J.M.B.; Felippe, M.J.B. Development, Phenotype, and Function of Non-Conventional B Cells. Comp. Immunol. Microbiol. Infect. Dis. 2017, 54, 38–44. [Google Scholar] [CrossRef]
- Wilkins, C.; Gale, M. Recognition of Viruses by Cytoplasmic Sensors. Curr. Opin. Immunol. 2010, 22, 41–47. [Google Scholar] [CrossRef]
- Braciale, T.J.; Hahn, Y.S. Immunity to Viruses. Immunol. Rev. 2013, 255, 5–12. [Google Scholar] [CrossRef]
- Makhluf, H.; Shresta, S. Innate Antiviral Immunity against Dengue Virus. Crit. Rev. Immunol. 2015, 35, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Trzeciak-Ryczek, A.; Tokarz-Deptuła, B.; Deptuła, W. Expression of IL-1β, IL-2, IL-10, TNF-β and GM-CSF in Peripheral Blood Leukocytes of Rabbits Experimentally Infected with Rabbit Haemorrhagic Disease Virus. Vet. Microbiol. 2016, 186, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Elfeil, W.M.K.; Algammal, A.M.; Abouelmaatti, R.R.; GErdouh, A.; Abdel-Daim, M.M. Molecular Characterization and Analysis of TLR-1 in Rabbit Tissues. Cent. Eur. J. Immunol. 2016, 41, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Marino, A.; Pergolizzi, S.; Cimino, F.; Lauriano, E.R.; Speciale, A.; D’Angelo, V.; Sicurella, M.; Argnani, R.; Manservigi, R.; Marconi, P. Role of Herpes Simplex Envelope Glycoprotein B and Toll-Like Receptor 2 in Ocular Inflammation: An Ex Vivo Organotypic Rabbit Corneal Model. Viruses 2019, 11, 819. [Google Scholar] [CrossRef]
- Astakhova, N.M.; Perelygin, A.A.; Zharkikh, A.A.; Lear, T.L.; Coleman, S.J.; MacLeod, J.N.; Brinton, M.A. Characterization of Equine and Other Vertebrate TLR3, TLR7, and TLR8 Genes. Immunogenetics 2009, 61, 529–539. [Google Scholar] [CrossRef]
- Kajikawa, O.; Frevert, C.W.; Lin, S.M.; Goodman, R.B.; Mongovin, S.M.; Wong, V.; Ballman, K.; Daubeuf, B.; Elson, G.; Martin, T.R. Gene Expression of Toll-like Receptor-2, Toll-like Receptor-4, and MD2 Is Differentially Regulated in Rabbits with Escherichia Coli Pneumonia. Gene 2005, 344, 193–202. [Google Scholar] [CrossRef]
- Lai, C.Y.; Liu, Y.L.; Yu, G.Y.; Maa, M.C.; Leu, T.H.; Xu, C.; Luo, Y.; Xiang, R.; Chuang, T.H. TLR7/8 Agonists Activate a Mild Immune Response in Rabbits through TLR8 but Not TLR7. Vaccine 2014, 32, 5593–5599. [Google Scholar] [CrossRef]
- Liu, J.; Xu, C.; Liu, Y.L.; Matsuo, H.; Pe-feng Hsieh, R.; Lo, J.F.; Tseng, P.H.; Yuan, C.J.; Luo, Y.; Xiang, R.; et al. Activation of Rabbit TLR9 by Different CpG-ODN Optimized for Mouse and Human TLR9. Comp. Immunol. Microbiol. Infect. Dis. 2012, 35, 443–451. [Google Scholar] [CrossRef]
- Abrantes, J.; Areal, H.; Esteves, P.J. Insights into the European Rabbit (Oryctolagus Cuniculus) Innate Immune System: Genetic Diversity of the Toll-like Receptor 3 (TLR3) in Wild Populations and Domestic Breeds. BMC Genet. 2013, 14. [Google Scholar] [CrossRef]
- Laliena, A.; Miguel, B.S.; Crespo, I.; Alvarez, M.; González-Gallego, J.; Tuñón, M.J. Melatonin Attenuates Inflammation and Promotes Regeneration in Rabbits with Fulminant Hepatitis of Viral Origin. J. Pineal. Res. 2012, 53, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Zhang, C.; Zhang, C.; Zhang, X.; Wu, Y. Functional Characterization of NLRX1 in Rabbit during Enterohemorrhagic Escherichia Coli Infection. Dev. Comp. Immunol. 2020, 106. [Google Scholar] [CrossRef] [PubMed]
- Lemos De Matos, A.; McFadden, G.; Esteves, P.J. Evolution of Viral Sensing RIG-I-like Receptor Genes in Leporidae Genera Oryctolagus, Sylvilagus, and Lepus. Immunogenetics 2014, 66, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Niedźwiedzka-Rystwej, P.; Tokarz-Deptuła, B.; Deptuła, W. Apoptosis of Granulocytes and Lymphocytes in Peripheral Blood in Rabbits Infected with Haemagglutinating and Non-Haemagglutinating Antigenic Variants of the RHD (Rabbit Haemorrhagic Disease) Virus. Pol. J. Vet. Sci. 2013, 16, 223–229. [Google Scholar] [CrossRef]
- Niedzwiedzka-Rystwej, P.; Tokarz-Deptula, B.; Hukowska-Szematowicz, B.; Dzialo, J.; Trzeciak-Ryczek, A.; Deptula, W. Peripheral Blood Lymphocytes B and T and Their Subpopulations in Rabbits Infected with Rabbit Haemorrhagic Disease Virus (RHDV). Med. Weter. 2013, 69, 484–487. [Google Scholar]
- Semerjyan, A.B.; Sargsyan, M.A.; Arzumanyan, H.H.; Hakobyan, L.H.; Abroyan, L.O.; Semerjyan, Z.B.; Avetisyan, A.S.; Karalova, E.M.; Manukyan, D.M.; Matevosyan, H.S.; et al. Immune Cell Pathology in Rabbit Hemorrhagic Disease. Vet. World 2019, 12, 1332–1340. [Google Scholar] [CrossRef] [PubMed]
- García-Lastra, R.; San-Miguel, B.; Crespo, I.; Jorquera, F.; Alvarez, M.; González-Gallego, J.; Tuñón, M.J. Signaling Pathways Involved in Liver Injury and Regeneration in Rabbit Hemorrhagic Disease, an Animal Model of Virally-Induced Fulminant Hepatic Failure. Vet. Res. 2010, 41. [Google Scholar] [CrossRef]
- Tuñon, M.J.; San Miguel, B.; Crespo, I.; Riezu-Boj, J.I.; Larrea, E.; Alvarez, M.; Gonzalez, I.; Bustos, M.; Gonzalez-Gallego, J.; Prieto, J. Cardiotrophin-1 Promotes a High Survival Rate in Rabbits with Lethal Fulminant Hepatitis of Viral Origin. J. Virol. 2011, 85, 13124–13132. [Google Scholar] [CrossRef] [PubMed]
- Tokarz-Deptuła, B. Immunity phenomena in rabbits infected with the RHD (rabbit haemorrhagic disease) virus. Pol. J. Environ. Stud. 2009, 7, 1–81. [Google Scholar]
- Bergin, I.L.; Wise, A.G.; Bolin, S.R.; Mullaney, T.P.; Kiupel, M.; Maes, R.K. Novel Calicivirus Identified in Rabbits, Michigan, USA. Emerg. Inf. Dis. 2009, 15, 1955–1962. [Google Scholar] [CrossRef] [PubMed]
- Kolaczkowska, E.; Kubes, P. Neutrophil Recruitment and Function in Health and Inflammation. Nat. Rev. Immunol. 2013, 13, 159–175. [Google Scholar] [CrossRef] [PubMed]
- Hrynkiewicz, R.; Bębnowska, D.; Niedźwiedzka-Rystwej, P. Myeloperoxidase and Lysozymes as a Pivotal Hallmark of Immunity Status in Rabbits. Animals 2020, 10, 1581. [Google Scholar] [CrossRef]
- Niedźwiedzka-Rystwej, P.; Deptuła, W. Non-Specific Immunity in Rabbits Infected with 10 Strains of the Rabbit Haemorrhagic Disease Virus with Different Biological Properties. Cent. Eur. J. Biol. 2010, 5, 613–632. [Google Scholar] [CrossRef]
- Zepeda-Cervantes, J.; Ramírez-Jarquín, J.O.; Vaca, L. Interaction Between Virus-Like Particles (VLPs) and Pattern Recognition Receptors (PRRs) From Dendritic Cells (DCs): Toward Better Engineering of VLPs. Front. Immunol. 2020, 11, 1100. [Google Scholar] [CrossRef]
- Ulsenheimer, A.; Gerlach, J.T.; Jung, M.C.; Gruener, N.; Wächtler, M.; Backmund, M.; Santantonio, T.; Schraut, W.; Heeg, M.H.J.; Schirren, C.A.; et al. Plasmacytoid Dendritic Cells in Acute and Chronic Hepatitis C Virus Infection. Hepatology 2005, 41, 643–651. [Google Scholar] [CrossRef]
- Bousarghin, L.; Hubert, P.; Franzen, E.; Jacobs, N.; Boniver, J.; Delvenne, P. Human Papillomavirus 16 Virus-like Particles Use Heparan Sulfates to Bind Dendritic Cells and Colocalize with Langerin in Langerhans Cells. J. Gen. Virol. 2005, 86, 1297–1305. [Google Scholar] [CrossRef] [PubMed]
- Mohsen, M.O.; Zha, L.; Cabral-Miranda, G.; Bachmann, M.F. Major Findings and Recent Advances in Virus–like Particle (VLP)-Based Vaccines. Semin. Immunol. 2017, 34, 123–132. [Google Scholar] [CrossRef] [PubMed]
- De Sousa-Pereira, P.; Abrantes, J.; Baldauf, H.-M.; Keppler, O.T.; Esteves, P.J. Evolutionary study of leporid CD4 reveals a hotspot of a genetic variability within the D2 domain. Immunogenetics 2016, 68, 477–482. [Google Scholar] [CrossRef]
- Bachmann, M.F.; Jennings, G.T. Vaccine Delivery: A Matter of Size, Geometry, Kinetics and Molecular Patterns. Nat. Rev. Immunol. 2010, 10, 787–796. [Google Scholar] [CrossRef] [PubMed]
- Serradell, M.C.; Rupil, L.L.; Martino, R.A.; Prucca, C.G.; Carranza, P.G.; Saura, A.; Fernández, E.A.; Gargantini, P.R.; Tenaglia, A.H.; Petiti, J.P.; et al. Efficient Oral Vaccination by Bioengineering Virus-like Particles with Protozoan Surface Proteins. Nat. Commun. 2019, 10. [Google Scholar] [CrossRef]
- Peñaflor-Téllez, Y.; Trujillo-Uscanga, A.; Escobar-Almazán, J.A.; Gutiérrez-Escolano, A.L. Immune Response Modulation by Caliciviruses. Front. Immunol. 2019, 10, 2334. [Google Scholar] [CrossRef] [PubMed]
- Ramiro-Ibáñez, F.; Martín-Alonso, J.M.; García Palencia, P.; Parra, F.; Alonso, C. Macrophage Tropism of Rabbit Hemorrhagic Disease Virus Is Associated with Vascular Pathology. Virus Res. 1999, 60, 21–28. [Google Scholar] [CrossRef]
- Prieto, J.M.; Fernandez, F.; Alvarez, V.; Espi, A.; García Marín, J.F.; Alvarez, M.; Martín, J.M.; Parra, F. Immunohistochemical Localisation of Rabbit Haemorrhagic Disease Virus VP-60 Antigen in Early Infection of Young and Adult Rabbits. Res. Vet. Sci. 2000, 68, 181–187. [Google Scholar] [CrossRef]
- Kimura, T.; Mitsui, I.; Okada, Y.; Furuya, T.; Ochiai, K.; Umemura, T.; Itakura, C. Distribution of Rabbit Haemorrhagic Disease Virus RNA in Experimentally Infected Rabbits. J. Comp. Pathol. 2001, 124, 134–141. [Google Scholar] [CrossRef]
- Boehm, T.; McCurley, N.; Sutoh, Y.; Schorpp, M.; Kasahara, M.; Cooper, M.D. VLR-Based Adaptive Immunity. Annu. Rev. Immunol. 2012, 30, 203–220. [Google Scholar] [CrossRef] [PubMed]
- Danilova, N. The Evolution of Adaptive Immunity. Adv. Exp. Med. Biol. 2012, 738, 218–235. [Google Scholar] [CrossRef]
- Farber, D.L.; Netea, M.G.; Radbruch, A.; Rajewsky, K.; Zinkernagel, R.M. Immunological Memory: Lessons from the Past and a Look to the Future. Nat. Rev. Immunol. 2016, 16, 124–128. [Google Scholar] [CrossRef] [PubMed]
- Chachu, K.A.; Strong, D.W.; LoBue, A.D.; Wobus, C.E.; Baric, R.S.; Virgin, H.W. Antibody Is Critical for the Clearance of Murine Norovirus Infection. J. Virol. 2008, 82, 6610–6617. [Google Scholar] [CrossRef] [PubMed]
- Lindesmith, L.C.; McDaniel, J.R.; Changela, A.; Verardi, R.; Kerr, S.A.; Costantini, V.; Brewer-Jensen, P.D.; Mallory, M.L.; Voss, W.N.; Boutz, D.R.; et al. Sera Antibody Repertoire Analyses Reveal Mechanisms of Broad and Pandemic Strain Neutralizing Responses after Human Norovirus Vaccination. Immunity 2019, 50, 1530–1541.e8. [Google Scholar] [CrossRef]
- Valíček, L.; Šmíd, B.; Rodák, L.; Kudrna, J. Electron and Immunoelectron Microscopy of Rabbit Haemorrhagic Disease Virus (RHDV). Arch. Virol. 1990, 112, 271–275. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Tian, X.; Zhai, Y.; Xu, W.; Zheng, D.; Sun, F. Cryo-electron microscopy reconstructions of two types of wild rabbit hemorrhagic disease viruses characterized the structural features of Lagovirus. Protein Cell 2010, 1, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Xu, F.; Liu, J.; Gao, B.; Liu, Y.; Zhai, Y.; Ma, J.; Zhang, K.; Baker, T.S.; Schulten, K.; et al. Atomic Model of Rabbit Hemorrhagic Disease Virus by Cryo-Electron Microscopy and Crystallography. PLoS Pathog. 2013, 9, e1003132. [Google Scholar] [CrossRef]
- Collins, B.J.; White, J.R.; Lenghaus, C.; Boyd, V.; Westbury, H.A. A Competition ELISA for the Detection of Antibodies to Rabbit Haemorrhagic Disease Virus. Vet. Microbiol. 1995, 43, 85–96. [Google Scholar] [CrossRef]
- Jennewein, M.F.; Abu-Raya, B.; Jiang, Y.; Alter, G.; Marchant, A. Transfer of Maternal Immunity and Programming of the Newborn Immune System. Semin. Immunopathol. 2017, 39, 605–613. [Google Scholar] [CrossRef]
- Furukawa, S.; Kuroda, Y.; Sugiyama, A. A Comparison of the Histological Structure of the Placenta in Experimental Animals. J. Toxicol. Pathol. 2014, 27, 11–18. [Google Scholar] [CrossRef]
- Marchandeau, S.; Chaval, Y.; Goff, E. le Prolonged Decline in the Abundance of Wild European Rabbits Oryctolagus Cuniculus and High Immunity Level over Three Years Following the Arrival of Rabbit Haemorrhagic Disease. Wildlife Biol. 2000, 6, 141–147. [Google Scholar] [CrossRef]
- Baratelli, M.; Molist-Badiola, J.; Puigredon-Fontanet, A.; Pascual, M.; Boix, O.; Mora-Igual, F.X.; Woodward, M.; Lavazza, A.; Capucci, L. Characterization of the Maternally Derived Antibody Immunity against Rhdv-2 after Administration in Breeding Does of an Inactivated Vaccine. Vaccines 2020, 8, 484. [Google Scholar] [CrossRef]
- Martens, C.L.; Currier, S.J.; Knight, K.L. Molecular genetic analysis of genes encoding the heavy chains of rabbit IgG. J. Immunol. 1984, 133, 1022–1027. [Google Scholar] [PubMed]
- Lavinder, J.J.; Hoi, K.H.; Reddy, S.T.; Wine, Y.; Georgiou, G. Systematic Characterization and Comparative Analysis of the Rabbit Immunoglobulin Repertoire. PLoS ONE 2014, 9, e101322. [Google Scholar] [CrossRef]
- Hayward, A.R.; Simons, M.A.; Lawton, A.R.; Mage, R.G.; Cooper, M.D. Pre-B and B Cells in Rabbits: Ontogeny and Allelic Exclusion of Kappa Light Chain Genes. J. Exp. Med. 1978, 148, 1367–1377. [Google Scholar] [CrossRef] [PubMed]
- McElroy, P.J.; Willcox, N.; Catty, D. Early Precursors of B Lymphocytes I. Rabbit/Mouse Species Differences in the Physical Properties and Surface Phenotype of Pre-B Cells, and in the Maturation Sequence of Early B Cells. Eur. J. Immunol. 1981, 11, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Jasper, P.J.; Zhai, S.-K.; Kalis, S.L.; Kingzette, M.; Knight, K.L. B Lymphocyte Development in Rabbit: Progenitor B Cells and Waning of B Lymphopoiesis. J. Immunol. 2003, 171, 6372–6380. [Google Scholar] [CrossRef]
- Neimanis, A.S.; Ahola, H.; Zohari, S.; Larsson Pettersson, U.; Bröjer, C.; Capucci, L.; Gavier-Widén, D. Arrival of Rabbit Haemorrhagic Disease Virus 2 to Northern Europe: Emergence and Outbreaks in Wild and Domestic Rabbits (Oryctolagus Cuniculus) in Sweden. Transbound. Emerg. Dis. 2018, 65, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.Y.; Lee, B.J.; Tai, J.H.; Park, J.H.; Lee, Y.S. Apoptosis in Rabbit Haemorrhagic Disease. J. Comp. Pathol. 2000, 123, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, P.G.; Costa-E-Silva, A.; Oliveira, M.J.R.; Monteiro, E.; Águas, A.P. Leukocyte-Hepatocyte Interaction in Calicivirus Infection: Differences between Rabbits That Are Resistant or Susceptible to Rabbit Haemorrhagic Disease (RHD). Vet. Immunol. Immunopathol. 2005, 103, 217–221. [Google Scholar] [CrossRef]
- Schwensow, N.I.; Detering, H.; Pederson, S.; Mazzoni, C.; Sinclair, R.; Peacock, D.; Kovaliski, J.; Cooke, B.; Fickel, J.; Sommer, S. Resistance to RHD Virus in Wild Australian Rabbits: Comparison of Susceptible and Resistant Individuals Using a Genomewide Approach. Mol. Ecol. 2017, 26, 4551–4561. [Google Scholar] [CrossRef]
- Fitzner, A.; Niedbalski, W. Serological Survey for RHD Antibodies in Rabbits from Two Types of Rabbit Breeding Farms. Pol. J. Vet. Sci. 2016, 19, 597–607. [Google Scholar] [CrossRef]
- Cox, T.E.; Liu, J.; van de Ven, R.; Strive, T. Different Serological Profiles to Co-Occurring Pathogenic and Nonpathogenic Caliciviruses in Wild European Rabbits (Oryctolagus Cuniculus) across Australia. J. Wildl. Dis. 2017, 53, 472–481. [Google Scholar] [CrossRef]
- Forrester, N.L.; Boag, B.; Buckley, A.; Moureau, G.; Gould, E.A. Co-Circulation of Widely Disparate Strains of Rabbit Haemorrhagic Disease Virus Could Explain Localised Epidemicity in the United Kingdom. Virology 2009, 393, 42–48. [Google Scholar] [CrossRef]
- Hall, R.N.; Capucci, L.; Matthaei, M.; Esposito, S.; Kerr, P.J.; Frese, M.; Strive, T. An in Vivo System for Directed Experimental Evolution of Rabbit Haemorrhagic Disease Virus. PLoS ONE 2017, 12, e0173727. [Google Scholar] [CrossRef]
- Abade dos Santos, F.A.; Magro, C.; Carvalho, C.L.; Ruivo, P.; Duarte, M.D.; Peleteiro, M.C. A Potential Atypical Case of Rabbit Haemorrhagic Disease in a Dwarf Rabbit. Animals 2021, 11, 40. [Google Scholar] [CrossRef]
- Le Gall-Reculé, G.; Zwingelstein, F.; Fages, M.P.; Bertagnoli, S.; Gelfi, J.; Aubineau, J.; Roobrouck, A.; Botti, G.; Lavazza, A.; Marchandeau, S. Characterisation of a Non-Pathogenic and Non-Protective Infectious Rabbit Lagovirus Related to RHDV. Virology 2011, 410, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Le Gall-Reculé, G.; Zwingelstein, F.; Boucher, S.; le Normand, B.; Plassiart, G.; Portejoie, Y.; Decors, A.; Bertagnoli, S.; Guérin, J.L.; Marchandeau, S. Virology: Detection of a New Variant of Rabbit Haemorrhagic Disease Virus in France. Vet. Rec. 2011, 168, 137–138. [Google Scholar] [CrossRef] [PubMed]
- Nagesha, H.S.; Wang, L.F.; Hyatt, A.D.; Morrissy, C.J.; Lenghaus, C.; Westbury, H.A. Self-Assembly, Antigenicity, and Immunogenicity of the Rabbit Haemorrhagic Disease Virus (Czechoslovakian Strain V-351) Capsid Protein Expressed in Baculovirus. Arch. Virol. 1995, 140, 1095–1108. [Google Scholar] [CrossRef]
- Fernández, E.; Toledo, J.R.; Chiong, M.; Parra, F.; Rodríguez, E.; Montero, C.; Méndez, L.; Capucci, L.; Farnós, O. Single Dose Adenovirus Vectored Vaccine Induces a Potent and Long-Lasting Immune Response against Rabbit Hemorrhagic Disease Virus after Parenteral or Mucosal Administration. Vet. Immunol. Immunopathol. 2011, 142, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Fernández, E.; Toledo, J.R.; Méndez, L.; González, N.; Parra, F.; Martín-Alonso, J.M.; Limonta, M.; Sánchez, K.; Cabrales, A.; Estrada, M.P.; et al. Conformational and Thermal Stability Improvements for the Large-Scale Production of Yeast-Derived Rabbit Hemorrhagic Disease Virus-Like Particles as Multipurpose Vaccine. PLoS ONE 2013, 8, e56417. [Google Scholar] [CrossRef]
- Guo, H.; Zhu, J.; Tan, Y.; Li, C.; Chen, Z.; Sun, S.; Liu, G. Self-Assembly of Virus-like Particles of Rabbit Hemorrhagic Disease Virus Capsid Protein Expressed in Escherichia Coli and Their Immunogenicity in Rabbits. Antiviral Res. 2016, 131, 85–91. [Google Scholar] [CrossRef]
- Mikschofsky, H.; Schirrmeier, H.; Keil, G.M.; Lange, B.; Polowick, P.L.; Keller, W.; Broer, I. Pea-Derived Vaccines Demonstrate High Immunogenicity and Protection in Rabbits against Rabbit Haemorrhagic Disease Virus. Plant Biotechnol. J. 2009, 7, 537–549. [Google Scholar] [CrossRef]
- Grgacic, E.V.L.; Anderson, D.A. Virus-like particles: Passport to immune recognition. Methods 2006, 40, 60–65. [Google Scholar] [CrossRef]
- Chen, Q.; Lai, H. Plant-Derived Virus-like Particles as Vaccines. Hum. Vaccin. Immunother. 2013, 9, 26–49. [Google Scholar] [CrossRef] [PubMed]
- Metz, S.W.; Martina, B.E.; van den Doel, P.; Geertsema, C.; Osterhaus, A.D.; Vlak, J.M.; Pijlman, G.P. Chikungunya Virus-like Particles Are More Immunogenic in a Lethal AG129 Mouse Model Compared to Glycoprotein E1 or E2 Subunits. Vaccine 2013, 31, 6092–6096. [Google Scholar] [CrossRef]
- Kushnir, N.; Streatfield, S.J.; Yusibov, V. Virus-like Particles as a Highly Efficient Vaccine Platform: Diversity of Targets and Production Systems and Advances in Clinical Development. Vaccine 2012, 31, 58–83. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Wang, S.; Zhang, W.; Liu, X.; Yi, Y.; Yang, S.; Xia, X.; Li, Y.; Zhang, Z. Development of a VLP-Based Vaccine in Silkworm Pupae against Rabbit Hemorrhagic Disease Virus. Int. Immunopharmacol. 2016, 40, 164–169. [Google Scholar] [CrossRef] [PubMed]
- Raeven, R.H.M.; van Riet, E.; Meiring, H.D.; Metz, B.; Kersten, G.F.A. Systems Vaccinology and Big Data in the Vaccine Development Chain. Immunology 2019, 156, 33–46. [Google Scholar] [CrossRef]
- Rappuoli, R. Reverse Vaccinology. Curr. Opin. Microbiol. 2000, 3, 445–450. [Google Scholar] [CrossRef]
- Fiuza, T.S.; Lima, J.P.M.S.; de Souza, G.A. EpitoCore: Mining Conserved Epitope Vaccine Candidates in the Core Proteome of Multiple Bacteria Strains. Front. Immunol. 2020, 11, 816. [Google Scholar] [CrossRef] [PubMed]
- Jain, S.; Baranwal, M. Conserved Peptide Vaccine Candidates Containing Multiple Ebola Nucleoprotein Epitopes Display Interactions with Diverse HLA Molecules. Med. Microbiol. Immunol. 2019, 208, 227–238. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).