
Optimization of Flow-Cytometry Based Assay for Measuring Neutralizing Antibody Responses against Each of the Four Dengue Virus Serotypes

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Cell Lines
2.2. Viral Growth and Titration
2.3. Monoclonal Antibodies
2.4. Plasma Samples
2.5. Focus Reduction Neutralization Test
2.6. Flow Cytometry Based Neutralization Assay
2.7. Statistical Analyses
2.8. Ethics Statement
3. Results
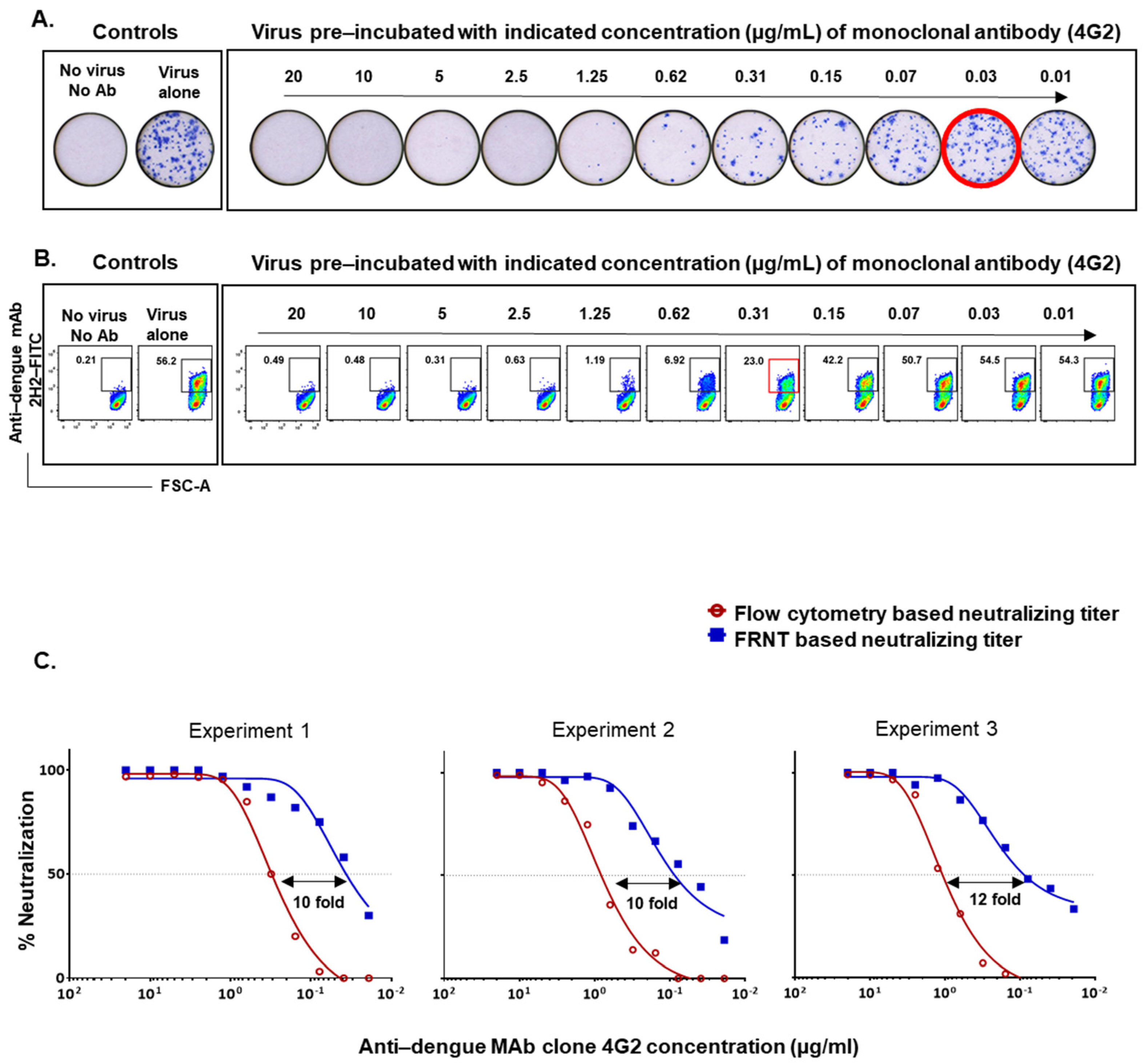
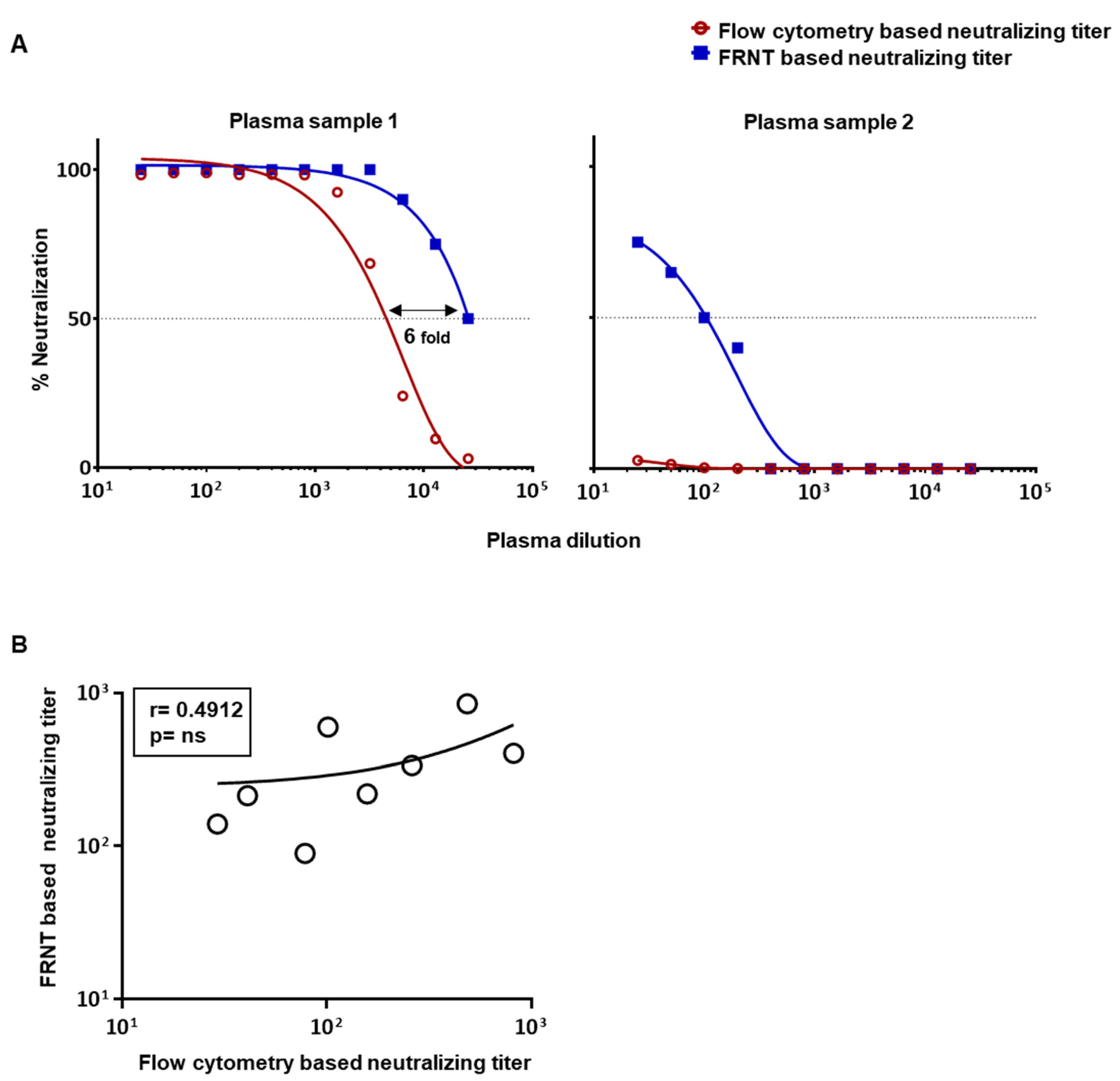
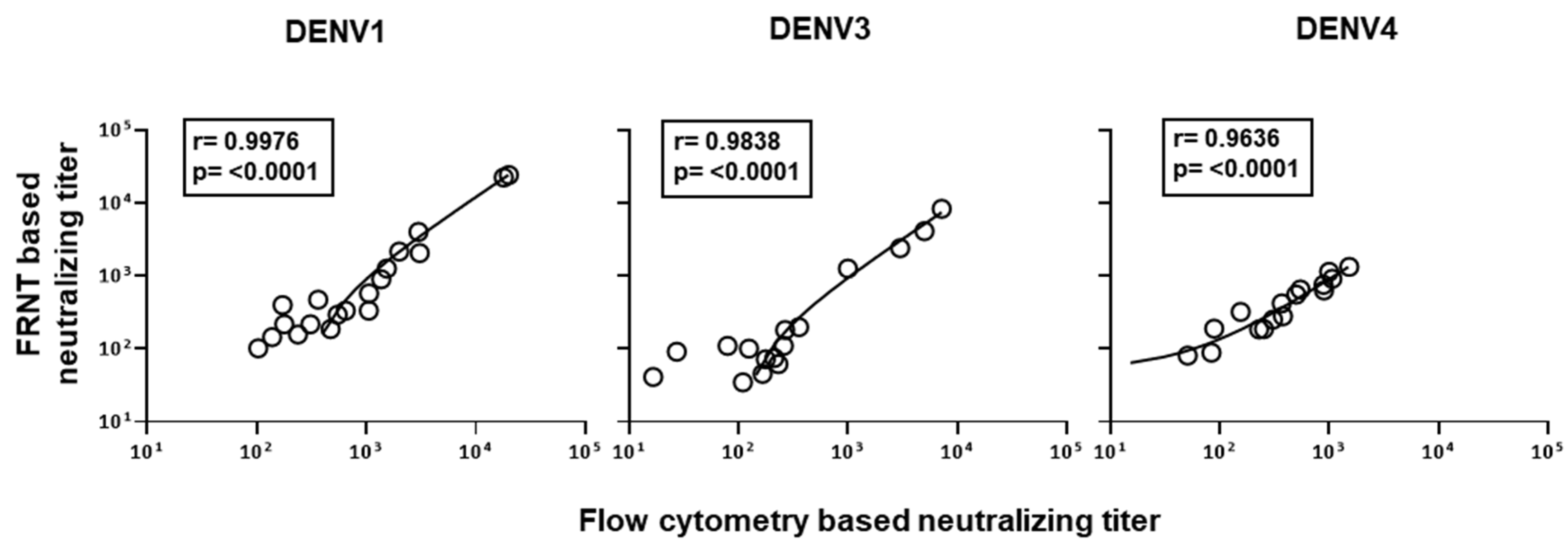
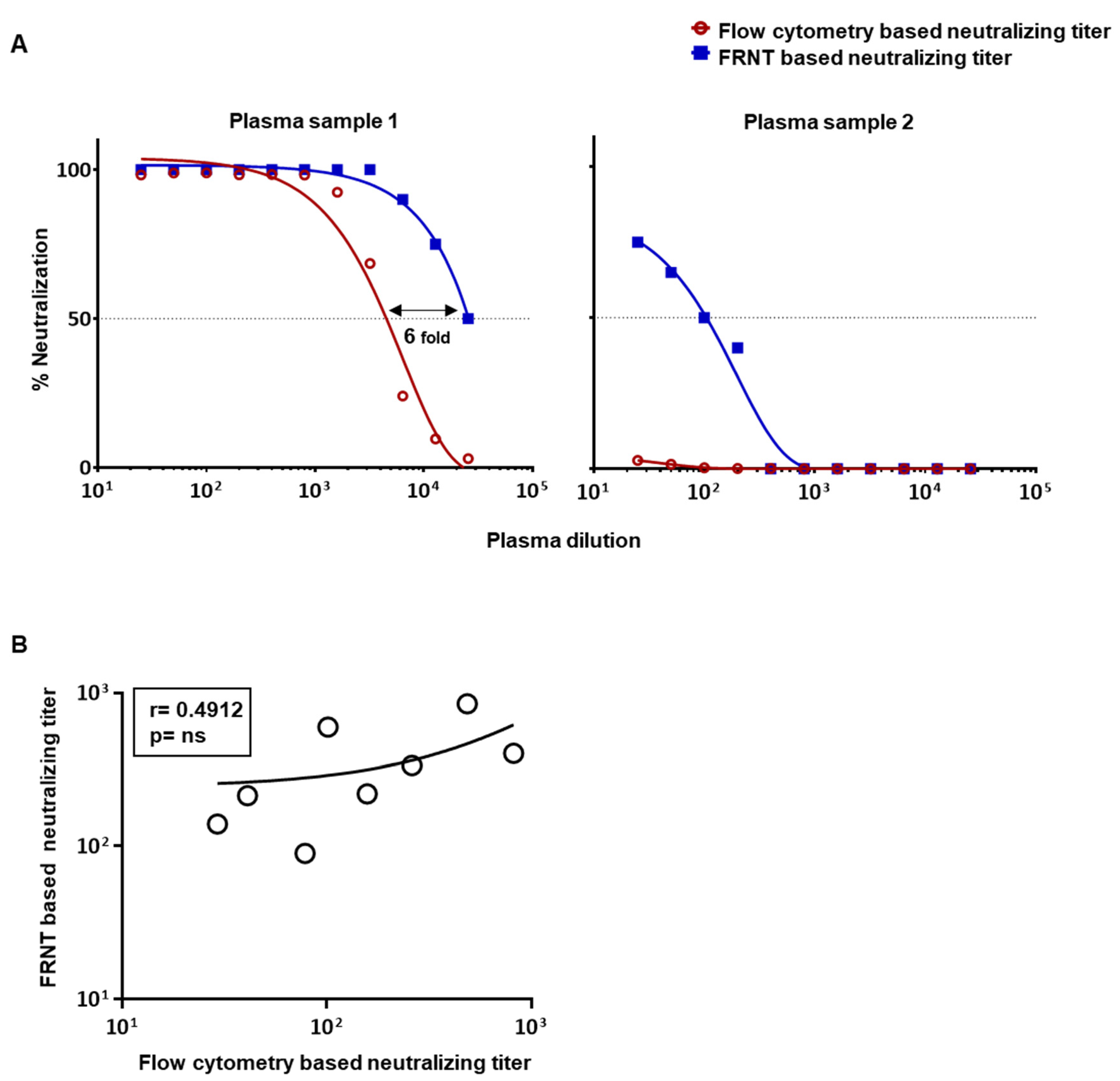
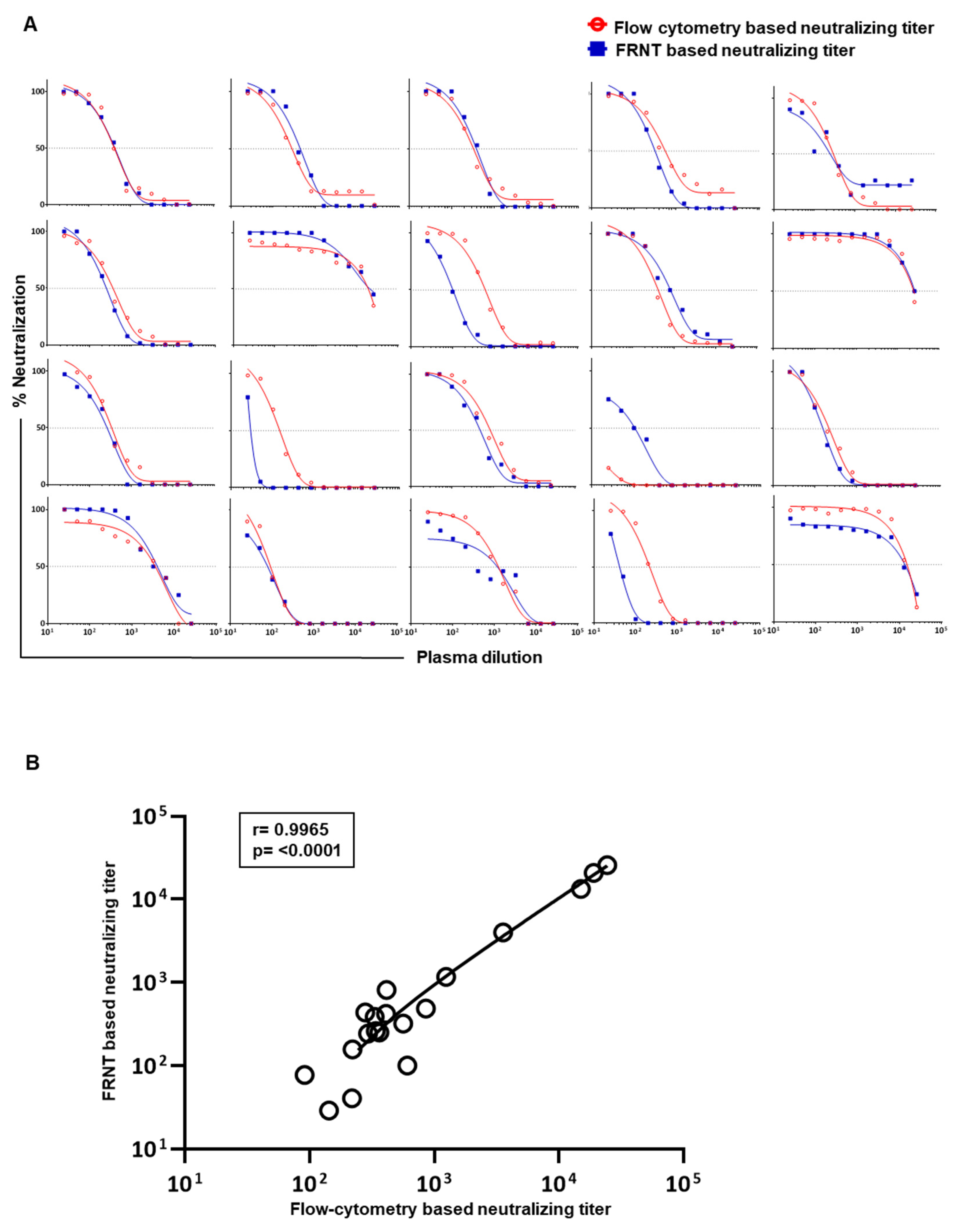
3.1. Comparison of Dengue Virus Specific Neutralization Titers Using the Standard Focus-Based and Flow Cytometry Based Neutralization Tests
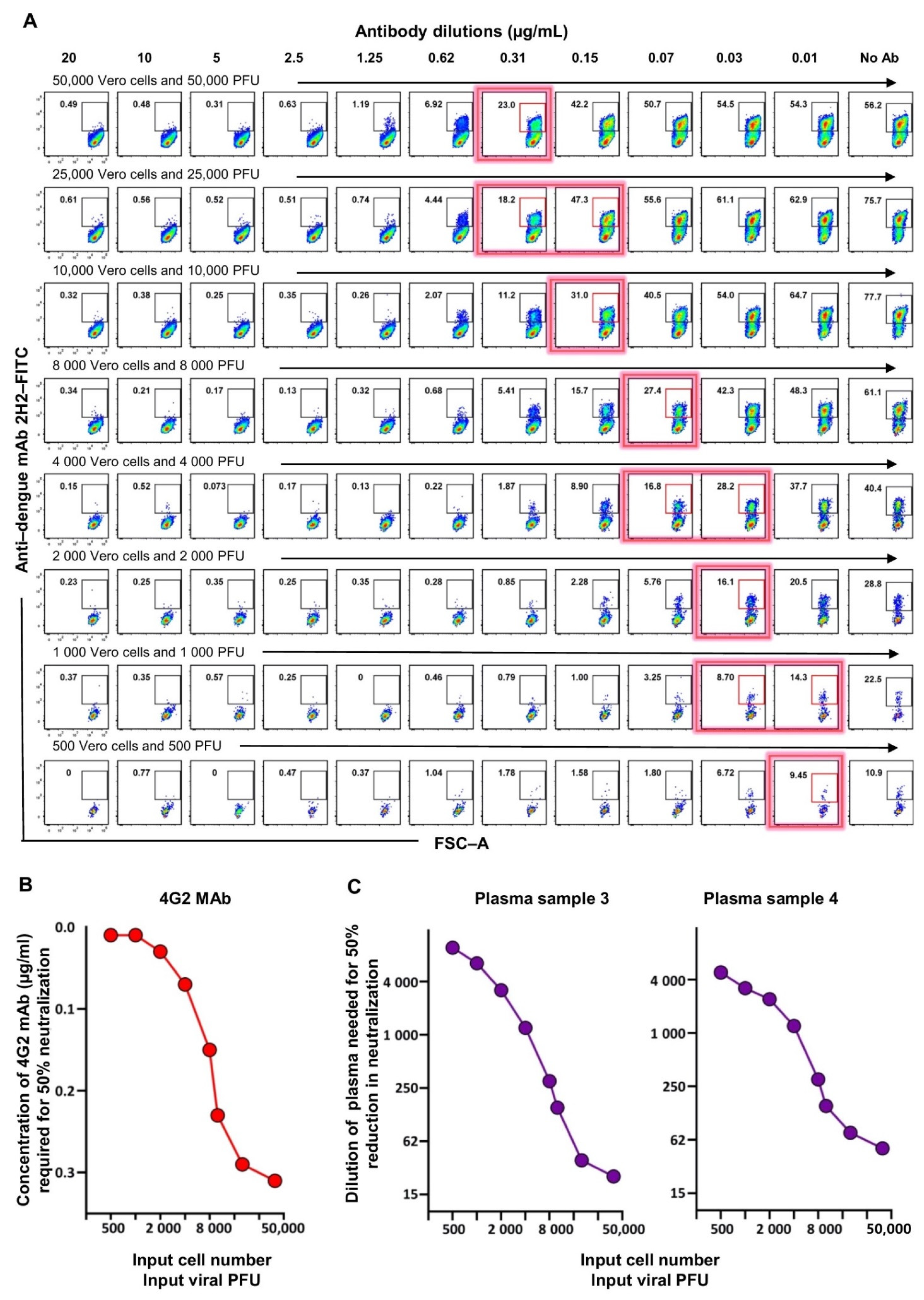
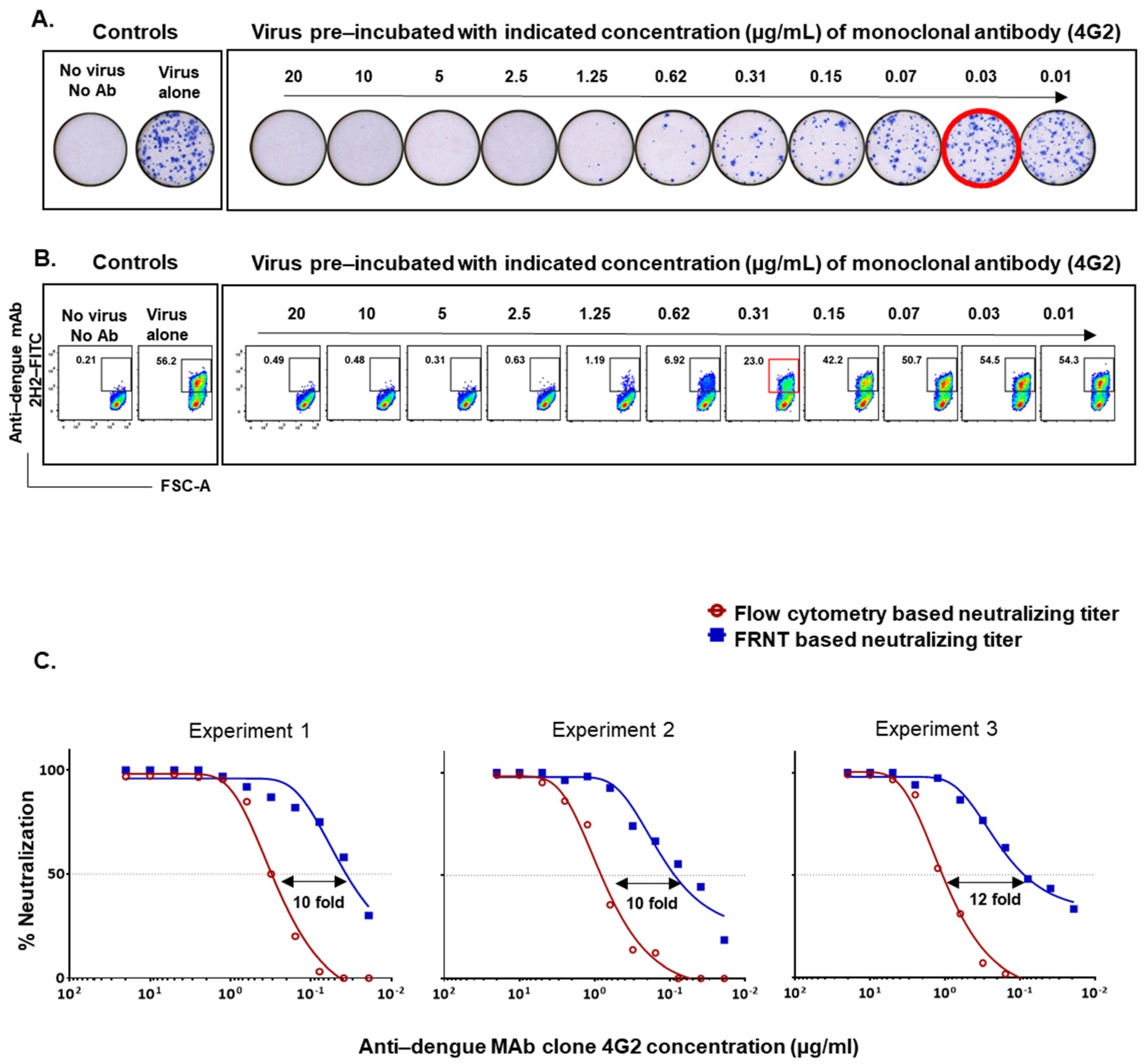
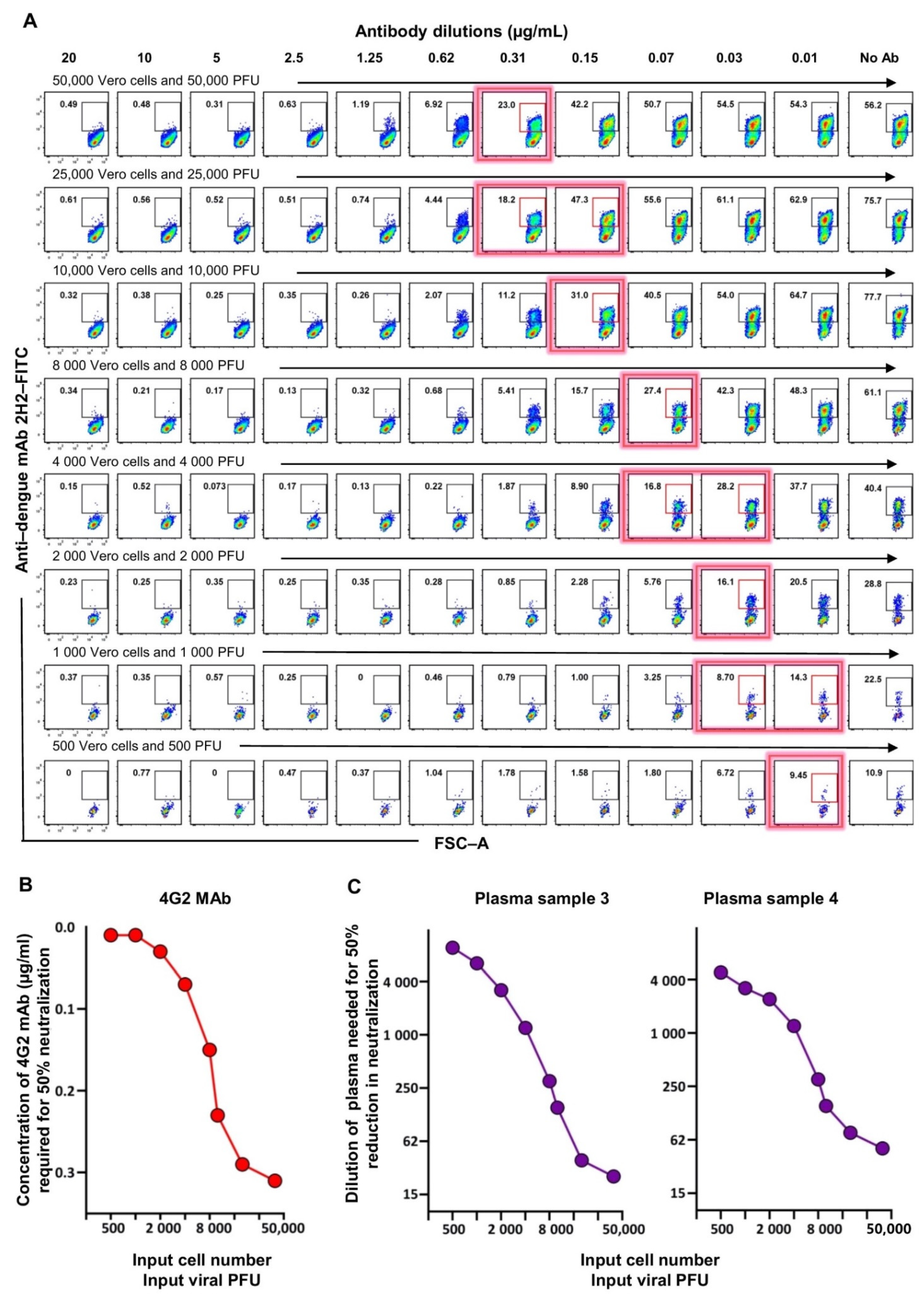
3.2. Optimization of the Flow-Based Assay Such That the Titers Obtained Are in Concordance with the Traditional Foci-Based Neutralizing Assay Titers
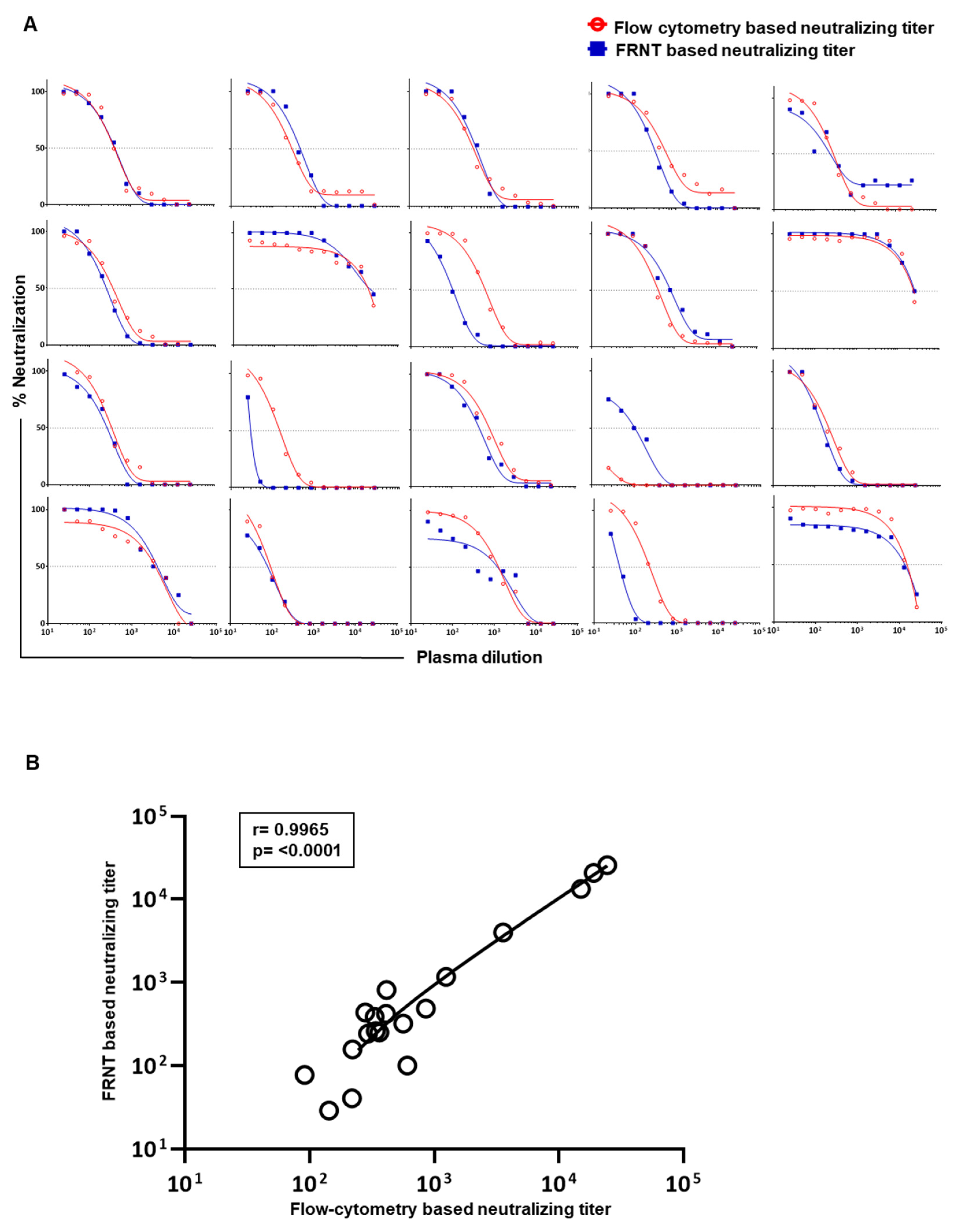
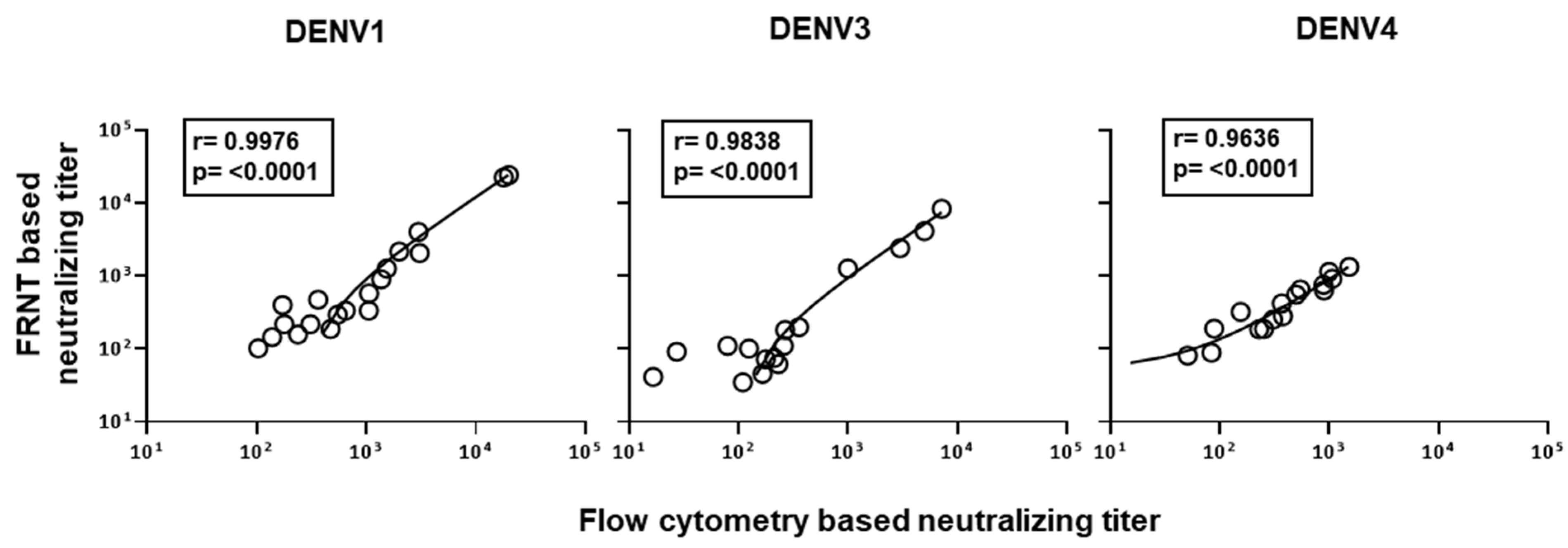
3.3. Evaluation of Correlation between the Neutralizing Titers by the Optimized Flow-Based Neutralizing Assay versus the Traditional Foci Reduction Neutralizing Assay
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bhatt, S.; Gething, P.; Brady, O.; Messina, J.P.; Farlow, A.W.; Moyes, C.; Drake, J.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef]
- Halstead, S.B. Dengue. Lancet 2007, 370, 1644–1652. [Google Scholar] [CrossRef]
- WHO. Dengue Guidelines for Diagnosis, Treatment, Prevention and Control; WHO: Geneva, Switzerland, 2009. [Google Scholar]
- Mutheneni, S.R.; Morse, A.P.; Caminade, C.; Upadhyayula, S.M. Dengue burden in India: Recent trends and importance of climatic parameters. Emerg. Microbes Infect. 2017, 6, e70. [Google Scholar] [CrossRef] [Green Version]
- Hadinegoro, S.R.; Arredondo-Garcia, J.L.; Capeding, M.R.; Deseda, C.; Chotpitayasunondh, T.; Dietze, R.; Ismail, H.I.M.; Reynales, H.; Limkittikul, K.; Rivera-Medina, D.M.; et al. Efficacy and Long-Term Safety of a Dengue Vaccine in Regions of Endemic Disease. N. Engl. J. Med. 2015, 373, 1195–1206. [Google Scholar] [CrossRef] [Green Version]
- Whitehead, S.S. Development of TV003/TV005, a single dose, highly immunogenic live attenuated dengue vaccine; what makes this vaccine different from the Sanofi-Pasteur CYD vaccine? Expert. Rev. Vaccines 2016, 15, 509–517. [Google Scholar] [CrossRef]
- Kraus, A.A.; Messer, W.; Haymore, L.B.; de Silva, A.M. Comparison of Plaque- and Flow Cytometry-Based Methods for Measuring Dengue Virus Neutralization. J. Clin. Microbiol. 2007, 45, 3777–3780. [Google Scholar] [CrossRef] [Green Version]
- Lambeth, C.R.; White, L.J.; Johnston, R.E.; de Silva, A.M. Flow Cytometry-Based Assay for Titrating Dengue Virus. J. Clin. Microbiol. 2005, 43, 3267–3272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timiryasova, T.M.; Bonaparte, M.I.; Luo, P.; Zedar, R.; Hu, B.T.; Hildreth, S.W. Optimization and Validation of a Plaque Reduction Neutralization Test for the Detection of Neutralizing Antibodies to Four Serotypes of Dengue Virus Used in Support of Dengue Vaccine Development. Am. J. Trop. Med. Hyg. 2013, 88, 962–970. [Google Scholar] [CrossRef]
- Russell, P.K.; Nisalak, A.; Sukhavachana, P.; Vivona, S. A plaque reduction test for dengue virus neutralizing antibodies. J. Immunol. 1967, 99, 285–290. [Google Scholar] [PubMed]
- Roehrig, J.; Hombach, J.; Barrett, A.D. Guidelines for Plaque-Reduction Neutralization Testing of Human Antibodies to Dengue Viruses. Viral Immunol. 2008, 21, 123–132. [Google Scholar] [CrossRef]
- Priyamvada, L.; Cho, A.; Onlamoon, N.; Zheng, N.-Y.; Huang, M.; Kovalenkov, Y.; Chokephaibulkit, K.; Angkasekwinai, N.; Pattanapanyasat, K.; Ahmed, R.; et al. B Cell Responses during Secondary Dengue Virus Infection Are Dominated by Highly Cross-Reactive, Memory-Derived Plasmablasts. J. Virol. 2016, 90, 5574–5585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gunisetty, S.; Nayak, K.; Rai, R.C.; Chawla, Y.; Reddy, E.S.; Aggarwal, C.; Maheshwari, D.; Panda, H.; Ansari, N.A.; Singh, P.; et al. Analysis of dengue specific memory B cells, neutralizing antibodies and binding antibodies in healthy adults from India. Int. J. Infect. Dis. 2019, 84, S57–S63. [Google Scholar] [CrossRef]
- Martin, N.C.; Pardo, J.; Simmons, M.; Tjaden, J.A.; Widjaja, S.; Marovich, M.A.; Sun, W.; Porter, K.R.; Burgess, T.H. An immunocytometric assay based on dengue infection via DC-SIGN permits rapid measurement of anti-dengue neutralizing antibodies. J. Virol. Methods 2006, 134, 74–85. [Google Scholar] [CrossRef]
- Matsuda, M.; Yamanaka, A.; Yato, K.; Yoshii, K.; Watashi, K.; Aizaki, H.; Konishi, E.; Takasaki, T.; Kato, T.; Muramatsu, M.; et al. High-throughput neutralization assay for multiple flaviviruses based on single-round infectious particles using dengue virus type 1 reporter replicon. Sci. Rep. 2018, 8, 16624. [Google Scholar] [CrossRef]
- Beltramello, M.; Williams, K.L.; Simmons, C.P.; Macagno, A.; Simonelli, L.; Quyen, N.T.H.; Sukupolvi-Petty, S.; Navarro-Sanchez, E.; Young, P.; de Silva, A.M.; et al. The Human Immune Response to Dengue Virus Is Dominated by Highly Cross-Reactive Antibodies Endowed with Neutralizing and Enhancing Activity. Cell Host Microbe 2010, 8, 271–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magnani, D.M.; Silveira, C.G.T.; Ricciardi, M.J.; Gonzalez-Nieto, L.; Pedreño-Lopez, N.; Bailey, V.K.; Gutman, M.J.; Maxwell, H.S.; Domingues, A.; Costa, P.R.; et al. Potent Plasmablast-Derived Antibodies Elicited by the National Institutes of Health Dengue Vaccine. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [Green Version]
- de Alwis, R.; de Silva, A.M. Measuring antibody neutralization of dengue virus (DENV) using a flow cytometry-based technique. Methods Mol. Biol. 2014, 1138, 27–39. [Google Scholar] [PubMed]
- Summers, P.L.; Cohen, W.H.; Ruiz, M.M.; Hase, T.; Eckels, K.H. Flaviviruses can mediate fusion from without in Aedes albopictus mosquito cell cultures. Virus Res. 1989, 12, 383–392. [Google Scholar] [CrossRef]
- Upasani, V.; Vo, H.T.M.; Auerswald, H.; Laurent, D.; Heng, S.; Duong, V.; Rodenhuis-Zybert, I.A.; Dussart, P.; Cantaert, T. Direct Infection of B Cells by Dengue Virus Modulates B Cell Responses in a Cambodian Pediatric Cohort. Front. Immunol. 2021, 11, 594813. [Google Scholar] [CrossRef]
- Chan, A.H.Y.; Tan, H.C.; Chow, A.Y.; Lim, A.P.C.; Lok, S.-M.; Moreland, N.; Vasudevan, S.; MacAry, P.; Ooi, E.E.; Hanson, B.J. A Human PrM Antibody That Recognizes a Novel Cryptic Epitope on Dengue E Glycoprotein. PLoS ONE 2012, 7, e33451. [Google Scholar] [CrossRef]
- de Alwis, R.; Williams, K.L.; Schmid, M.A.; Lai, C.Y.; Patel, B.; Smith, S.A.; Crowe, J.E.; Wang, W.K.; Harris, E.; de Silva, A.M. Faculty Opinions recommendation of Dengue viruses are enhanced by distinct populations of serotype cross-reactive antibodies in human immune sera. PLoS Pathog. 2016, 10, e1004386. [Google Scholar] [CrossRef]
- Ranganathan, P.; Aggarwal, R. Common pitfalls in statistical analysis: The use of correlation techniques. Perspect. Clin. Res. 2016, 7, 187–190. [Google Scholar] [CrossRef] [PubMed]
- Deng, S.-Q.; Yang, X.; Wei, Y.; Chen, J.-T.; Wang, X.-J.; Peng, H.-J. A Review on Dengue Vaccine Development. Vaccines 2020, 8, 63. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, L.M.; Halloran, M.E.; Durbin, A.P.; Longini, I.M. The dengue vaccine pipeline: Implications for the future of dengue control. Vaccine 2015, 33, 3293–3298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajamanonmani, R.; Nkenfou, C.; Clancy, P.; Yau, Y.H.; Shochat, S.G.; Sukupolvi-Petty, S.; Schul, W.; Diamond, M.S.; Vasudevan, S.G.; Lescar, J. On a mouse monoclonal antibody that neutralizes all four dengue virus serotypes. J. Gen. Virol. 2009, 90, 799–809. [Google Scholar] [CrossRef]
- Lim, X.N.; Shan, C.; Marzinek, J.K.; Dong, H.; Ng, T.S.; Ooi, J.S.G.; Fibriansah, G.; Wang, J.; Verma, C.S.; Bond, P.J.; et al. Molecular basis of dengue virus serotype 2 morphological switch from 29 °C to 37 °C. PLoS Pathog. 2019, 15, e1007996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waickman, A.T.; Gromowski, G.D.; Rutvisuttinunt, W.; Li, T.; Siegfried, H.; Victor, K.; Kuklis, C.; Gomootsukavadee, M.; McCracken, M.K.; Gabriel, B.; et al. Transcriptional and clonal characterization of B cell plasmablast diversity following primary and secondary natural DENV infection. EBioMedicine 2020, 54, 102733. [Google Scholar] [CrossRef]
- Henchal, E.A.; McCown, J.M.; Gentry, M.K.; Brandt, W.E. Dengue Virus-Specific and Flavivirus Group Determinants Identified with Monoclonal Antibodies by Indirect Immunofluorescence. Am. J. Trop. Med. Hyg. 1982, 31, 830–836. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharma, P.; Nayak, K.; Reddy, E.S.; Farooqi, H.; Murali-Krishna, K.; Chandele, A. Optimization of Flow-Cytometry Based Assay for Measuring Neutralizing Antibody Responses against Each of the Four Dengue Virus Serotypes. Vaccines 2021, 9, 1339. https://doi.org/10.3390/vaccines9111339
Sharma P, Nayak K, Reddy ES, Farooqi H, Murali-Krishna K, Chandele A. Optimization of Flow-Cytometry Based Assay for Measuring Neutralizing Antibody Responses against Each of the Four Dengue Virus Serotypes. Vaccines. 2021; 9(11):1339. https://doi.org/10.3390/vaccines9111339
Chicago/Turabian StyleSharma, Pragati, Kaustuv Nayak, Elluri Seetharami Reddy, Humaira Farooqi, Kaja Murali-Krishna, and Anmol Chandele. 2021. "Optimization of Flow-Cytometry Based Assay for Measuring Neutralizing Antibody Responses against Each of the Four Dengue Virus Serotypes" Vaccines 9, no. 11: 1339. https://doi.org/10.3390/vaccines9111339
APA StyleSharma, P., Nayak, K., Reddy, E. S., Farooqi, H., Murali-Krishna, K., & Chandele, A. (2021). Optimization of Flow-Cytometry Based Assay for Measuring Neutralizing Antibody Responses against Each of the Four Dengue Virus Serotypes. Vaccines, 9(11), 1339. https://doi.org/10.3390/vaccines9111339